一种非同源末端连接检测系统及其应用
本发明属于基因工程领域,具体的涉及基于基因工程方法对肿瘤相关药物的筛选和开发领域。
背景技术:
恶性肿瘤是目前危害大众健康的主要疾病,放射治疗、化学药物治疗是目前术后治疗的主要手段。放射治疗与dna损伤类化疗药物(铂类、环磷酰胺类等)主要通过诱导基因组dna双链断裂(dnadouble-strandbreaks,dsbs)所产生的细胞毒性发挥治疗效用。基因组dna双链断裂是细胞面对的最危险的损伤之一,有报道称细胞内单个未修复dsb就足以启动细胞程序性死亡(programmedcelldeath)。为应对dsb以保持基因组的稳定性,细胞演化出一套有效的dsb位点识别、信号传导、细胞周期调控、损伤修复系统。真核细胞有两种dsb损伤修复途径:同源重组(homologousrecombination,hr)和非同源末端连接(non-homologousendjoining,nhej)。同源重组修复正确率高(error-free)需要完好的姊妹染色体作为模板,通常发生在细胞周期的s、g2期。非同源末端连接在细胞周期的各个阶段都可进行,不需要修复模板,通常会引入损伤位点附近的碱基插入或缺失(error-prone)。非同源末端连接虽易引入突变,但却修复了约85%的dsbs。肿瘤细胞对dsb修复的效率是影响放射治疗及dna损伤化疗药物效果的主要因素。因此,抑制dsb修复通路是提高放化疗效果的关键。目前,通过筛选靶向dsb修复通路中关键蛋白的小分子化合物发现了大量dsb修复的抑制因子。例如,nu7441、ku0060648可抑制dna-pkcs,ku55933抑制atm。
近年来,基于不同的原理和检测方法,研究者已经开发出多种nhej报告系统。这些测定法采用伽马射线,i-scei核酸酶,或crispr-cas9核酸酶产生dsb,使用中性单细胞电泳检测,荧光蛋白测定,克隆形成实验,免疫荧光染色,或高分辨率熔解曲线(hrm)分析来监测nhej修复活性。这些方法需要辐射,流式细胞仪,免疫荧光成像,或基因组dna提取和实时定量pcr。这些繁琐的步骤和对昂贵仪器的依赖限制了这些系统应用,尤其是从大型化合物库中高通量筛选nhej抑制剂时的实用性。荧光素酶生物发光在高通量筛选中一直保持着突出的地位。基于荧光素酶的报告基因检测提供了灵敏的方法来量化基因表达的细微变化。
以crispr为代表的基因编辑技术彻底改变了生命科学的研究手段,也为dsb损伤修复研究带来了新的研究工具。因此,目前急需开发一种基于crispr系统和荧光素酶生物发光,操作方便,灵敏,可在大规模小分子化合物库中筛选dsb损伤修复抑制剂的方法。
技术实现要素:
本发明提供了一种非同源末端连接(non-homologousendjoining,nhej)检测系统,包括基于crispr系统的报告基因体系和报告基因检测体系,所述基于crispr系统的报告基因体系包括荧光素酶报告基因突变序列,所述荧光素酶报告基因突变序列使荧光素酶报告基因处于关闭状态;利用crispr系统在所述荧光素酶报告基因突变序列引入dna双链断裂,经非同源末端连接使所述荧光素酶报告基因突变序列修复,荧光素酶报告基因读码框恢复正常,使荧光素酶报告基因打开。本发明所述荧光素酶报告基因关闭是指荧光素酶基因不表达,不产生生物发光。本发明所述荧光素酶报告基因打开是指荧光素酶基因表达,产生生物发光,并且只有在cripsr系统切割产生dna双链断裂后,经过nhej后,才有荧光素酶信号。本发明优选的cripsr系统为cripsr-cas9系统。
优选的是,所述荧光素酶报告基因突变序列是在荧光素酶报告基因开放阅读框引入移码突变导致荧光素酶报告基因表达失败,进而不产生生物发光(bioluminescence),即荧光素酶报告基因处于关闭状态;crispr系统通过引入dna双链断裂(dsb)触发非同源末端链接修复所述移码突变,使荧光素酶报告基因开放阅读恢复表达。
上述任一项中优选的是,所述荧光素酶报告基因突变序列通过向所述荧光素酶报告基因序列开放阅读框中引入碱基得到。
本发明优选的方案是,在荧光素酶报告基因的开放阅读框中引入碱基得到荧光素酶报告基因突变序列,使荧光素酶基因发生移码突变,进而使荧光素酶报告基因关闭。然后通过crispr系统在所述荧光素酶报告基因突变序列的基因突变位点引入dsb,在接下来触发的nhej中,断裂的dna连接的同时引入的碱基被切除,从而使荧光素酶报告基因的阅读框恢复,荧光素酶报告基因打开,荧光素酶表达,产生生物发光。本发明的特点在于,1)荧光素酶报告基因的突变位于荧光素酶报告基因开放阅读框内,因而在荧光素酶报告基因处于关闭状态时,由于其本身的阅读框破坏,不会产生背景荧光。进一步优选的是,在靠近荧光素酶的n端(编码区的5’端)引入碱基,尽量靠近n端可保证荧光素酶基因的大部分不被表达,不产生荧光信号,如果靠近c端,则大部分荧光素酶片段表达,有可能具有酶活性,产生荧光信号;2)引入突变的位点与crispr系统引入dsb的位点重叠,即crispr系统的grna识别位点为引入碱基的突变位点。本发明应用于各种荧光素酶基因,包括fireflyluciferase,renillaluciferase,gaussialuciferase,vargulaluciferases等,以作为nhejreporter。
上述任一项中优选的是,所述荧光素酶报告基因突变序列为seqidno:1所示的核苷酸序列,荧光素酶优选为fireflyluciferase。
上述任一项中优选的是,所述检测系统,包括以下步骤:
步骤1:在荧光素酶报告基因的205位点插入两个碱基ctoatc,得到所述荧光素酶报告基因突变序列;
步骤2:将所述荧光素酶报告基因突变序列克隆于质粒载体后,包装病毒,感染细胞,产生基因组稳定整合的动物细胞;
步骤3:利用crispr系统打开荧光素酶报告基因的表达,crispr系统中包含的grna的核苷酸序列如seqidno:2所示;
步骤4:通过检测荧光素酶报告系统的荧光信号对非同源末端连接进行检测。
dsb经nhej修复后,会产生不同长度的碱基插入、删除。主要是删除。一部分碱基删除会恢复荧光素酶野生型长度(1653bp),即删去2个碱基。(荧光素酶报告基因突变序列为1655bp)
本发明还提供了一种非同源末端连接损伤修复抑制剂的筛选方法,将目的试剂作用于上述任一项所述的非同源末端连接检测系统,如果荧光素酶报告基因的荧光信号下降,则目的试剂为非同源末端连接损伤修复的抑制剂。
本发明还提供了一种化疗药物的筛选方法,利用权利要求1至5任一项所述的非同源末端连接检测系统,筛选针对dna双链断裂损伤修复通路的抑制剂。
优选的,将目的试剂作用于所述的非同源末端连接检测系统,如果荧光素酶报告基因的荧光信号下降,则目的试剂为非同源末端连接损伤修复的抑制剂。
crispr系统广泛存在于细菌之中,是细菌为抵御病毒入侵或质粒转移所进化出的一种保护机制。crispr系统作用于入侵病毒的靶dna链而使其产生dsb。非同源末端连接(nhej)是真核细胞dsb损伤修复细胞的主要途径之一。非同源末端连接(nhej)可以在细胞周期的各个阶段进行,且不需要修复模板,通常会引入损伤位点附近的碱基插入或缺失。虽然非同源末端连接(nhej)常引入突变,但却修复了85%的dsb。本发明所提供的一种基于crispr系统的荧光素酶报告基因打开(turnon)nhej检测系统,并且操作方便,可大规模的在小分子化合物库中筛选dsb损伤修复抑制剂。
附图说明
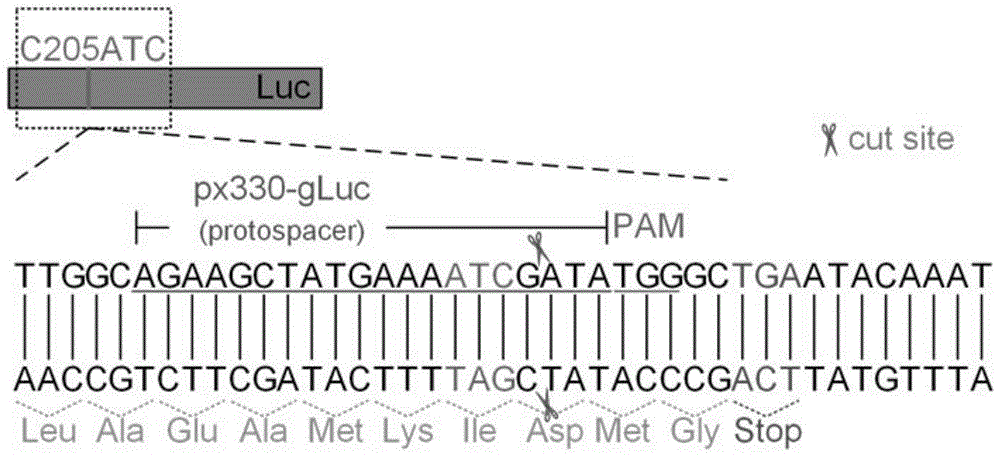
图1本发明优选实施例1荧光素酶报告基因205位点插入两个碱基后序列图。
图2本发明优选实施例1基于crispr系统的荧光素酶报告基因打开dsb检测系统模式图。
图3本发明优选实施例1荧光素酶载体亚克隆到pcdna3.0载体,荧光素酶信号继续沉默结果图。
图4本发明优选实施例1px330空载体,px330-gluc的荧光素酶信号图。
图5本发明优选实施例2nu7441、ku0060648、ku55933作用于非同源末端连接检测系统结果图。
具体实施方式
本发明通过以下实施例进行更加清晰、完整的描述,但所描述的实例仅是本发明一部分实施例,并非全部。所述实施例为帮助理解本发明,不应依此来局限本发明的保护范围。本发明所使用试剂均为市售商业化试剂,相关实验步骤如无特殊说明均为现有技术中的常规操作,记载于相关文献及工具书中。
实施例1
实施例1提供了一种基于crispr系统的荧光素酶报告基因打开(turnon)dsb检测系统,其突变序列比对及原理如图1、2所示。
在荧光素酶报告基因的205位点插入两个碱基得到seqidno:1所示的荧光素酶报告基因突变序列,如图1所示,荧光素酶报告基因读码框破坏而使荧光素酶失活,荧光素酶报告基因关闭。通过crispr系统(实施例1优选为cripsr-cas9系统),在205位点附近引入dsb,断裂的双链dna经非同源末端连接(nhej)修复产生插入、缺失突变(indel),使荧光素酶报告基因读码框恢复正常进而产生荧光素酶活性,如图2所示,荧光素酶报告基因打开。使用此系统可以快速筛选dsb损伤修复的抑制剂。
实施例1中,所述检测系统在荧光素酶报告基因205位点插入两个碱基(ctoatc),得到荧光素酶报告基因突变序列,使荧光素酶失活,只产生少量的荧光背景信号,如图3所示。本实施例seqidno:1所示的核苷酸序列通过生物公司合成。
实施例1中,将荧光素酶报告基因突变序列(荧光素酶报告基因ctoatc)克隆到pcdna3.0载体中的hindⅲ和bamhⅰ限制酶酶切位点之间得到pcdna3.0-luc(c205atc)。pcdna3.0-luc(c205atc)质粒构建过程为本领域的常规操作。突变序列相对于野生型序列在205位点引入2bp碱基,在产生移码突变的同时引入提前终止密码子。
将荧光素酶突变序列luc(c205atc)包装慢病毒,感染hek293t细胞,获得稳定整合的hek293t细胞系,即筛选获得稳定表达含205ctoatc突变荧光素酶的hek293t细胞系。本发明采用的慢病毒系统包括三个包装载体(pcmv-vsv-g,pmdlg/prre和prsv-rev)。突变luc(c205atc)被亚克隆到plv-ef1a-mcs-ires-bsd载体中。将上述四个质粒共转染到hek293t细胞中以包装慢病毒。通过慢病毒感染和随后的bsd选择产生了稳定表达luc(c205atc)的hek293t细胞,此细胞系用来进行抑制剂筛选。具体步骤及慢病毒系统中各质粒的来源、作用参见:engulfedcadherinfingersarepolarizedjunctionalstructuresbetweencollectivelymigratingendothelialcells.doi:10.1038/ncb3438.本发明中,pcdna3.0-luc(c205atc)将携带的luc(c205atc)通过酶切、连接等分子克隆常规手段将luc(c205atc)序列被克隆到plv-ef1a-mcs-ires-bsd载体中。本发明中,pcdna3.0-luc(c205atc)另一作用为测定荧光素酶是否沉默,如图3所示。
将grna构建于px330质粒。px330质粒购自addgene(plasmid#42230),经bbsi酶切,编码grna的引物经退火后形成双链,通过酶切位点购入载体。如seqidno:2所示的grna构建于px330质粒,得到px330-gluc质粒;如seqidno:3所示的随机grna序列构建于px330质粒,得到px330-scrambled质粒。
用px330空载体、px330-gluc分别转染所述稳定表达含205ctoatc突变荧光素酶的hek293t细胞系。转染前一天将所述稳定表达含205ctoatc突变荧光素酶的hek293t细胞系接种到24孔板中。使用lipofectamine2000试剂将500ngpx330、500ngpx330-gluc分别和10ngprl-cmv载体(prl-cmv购自promega公司,编码renilla荧光素酶基因,在双荧光素酶报告实验中起内参的作用)共转染到所述稳定表达含205ctoatc突变荧光素酶的hek293t细胞系中。转染48小时后,使用双荧光素酶报告基因检测系统(promega)测量荧光素酶活性。结果显示px330-gluc可以有效的打开(turnon)荧光素酶基因的表达,如图4所示,基于荧光素酶测量,本申请提供的检测系统较背景荧光信号升高了3000多倍。(本发明采用的crispr系统中,px330载体同时表达guildrna和cas9蛋白,具体操作方法参见:science.2013;339(6121):819-23.doi:10.1126/science.1231143.multiplexgenomeengineeringusingcrispr/cassystems)。
实施例2
实施例2利用实施例1所提供的dsb检测系统,对其作为nhej损伤修复的抑制剂筛选方法进行验证。nu7441、ku0060648、ku55933为已知nhej的抑制剂。转染前一天将所述稳定表达含205ctoatc突变荧光素酶的hek293t细胞系接种到24孔板中。使用lipofectamine2000试剂将500ngpx330、500ngpx330-gluc、500ngpx330-scrambled分别和10ngprl-cmv载体共转染到所述稳定表达含205ctoatc突变荧光素酶的hek293t细胞系中。转染24小时后,将不同浓度的nhej抑制剂nu7441(0.01μm、0.1μm、0.5μm、1μm、5μm)、ku0060648(0.01μm、0.1μm、0.5μm、1μm)、ku55933(0.01μm、0.1μm、1μm、5μm、10μm、20μm)分别添加到细胞培养基,其中,每种nhej抑制剂的不同浓度均为4重复,并孵育48小时。48小时后,使用双荧光素酶报告基因检测系统(promega)测量荧光素酶活性。结果显示,将此三种抑制剂作用于此系统,可以发现荧光素酶信号呈浓度梯度依赖性下降,如图5所示,nu7441、ku0060648、ku55933呈浓度梯度依赖性的抑制效果。(nu7441、ku0060648、ku55933购自selleckchemicals(美国德克萨斯州休斯顿)(px330-scrambled为含有编码随机grna序列的载体,和px330一样起到对照质粒的作用。本发明优选的grna随机序列由randomdnasequencegenerator随机序列生成器产生(http://faculty.ucr.edu/~mmaduro/random.htm))。
利用深度测序方法对上述结果进行检测验证发现,px330-gluc转染后会导致3246±262rpm(三次实验平均±标准差)的野生型长度的荧光素酶读数。rpm是readpermillion的缩写,意思是每百万个读数(read)中野生型长度出现的次数。深度测序结果只是辅助说明系统的特征。图5通过对已知nhejinhibitor的反应证明了本申请的有效性。本申请提供的方法具有灵敏度高的特点,5μmnu7441处理导致luc活性下降约75%(p<0.01),1μmnu7441处理luc活性下降约35%(p<0.01)。
本实施例通过对已知nhej抑制剂对本发明所提供的检测系统进行验证,表现良好,表明此系统可以有效筛选针对dna双链断裂损伤修复通路的抑制剂,为发现新的放化疗增敏剂以及新的dna损伤化疗药物打下基础。
序列表
<110>天津市肿瘤医院(天津医科大学肿瘤医院)
<120>一种非同源末端连接检测系统及其应用
<160>3
<170>siposequencelisting1.0
<210>1
<211>1655
<212>dna
<213>人工序列(artificialsequence)
<400>1
atggaagacgccaaaaacataaagaaaggcccggcgccattctatccgctggaagatgga60
accgctggagagcaactgcataaggctatgaagagatacgccctggttcctggaacaatt120
gcttttacagatgcacatatcgaggtggacatcacttacgctgagtacttcgaaatgtcc180
gttcggttggcagaagctatgaaaatcgatatgggctgaatacaaatcacagaatcgtcg240
tatgcagtgaaaactctcttcaattctttatgccggtgttgggcgcgttatttatcggag300
ttgcagttgcgcccgcgaacgacatttataatgaacgtgaattgctcaacagtatgggca360
tttcgcagcctaccgtggtgttcgtttccaaaaaggggttgcaaaaaattttgaacgtgc420
aaaaaaagctcccaatcatccaaaaaattattatcatggattctaaaacggattaccagg480
gatttcagtcgatgtacacgttcgtcacatctcatctacctcccggttttaatgaatacg540
attttgtgccagagtccttcgatagggacaagacaattgcactgatcatgaactcctctg600
gatctactggtctgcctaaaggtgtcgctctgcctcatagaactgcctgcgtgagattct660
cgcatgccagagatcctatttttggcaatcaaatcattccggatactgcgattttaagtg720
ttgttccattccatcacggttttggaatgtttactacactcggatatttgatatgtggat780
ttcgagtcgtcttaatgtatagatttgaagaagagctgtttctgaggagccttcaggatt840
acaagattcaaagtgcgctgctggtgccaaccctattctccttcttcgccaaaagcactc900
tgattgacaaatacgatttatctaatttacacgaaattgcttctggtggcgctcccctct960
ctaaggaagtcggggaagcggttgccaagaggttccatctgccaggtatcaggcaaggat1020
atgggctcactgagactacatcagctattctgattacacccgagggggatgataaaccgg1080
gcgcggtcggtaaagttgttccattttttgaagcgaaggttgtggatctggataccggga1140
aaacgctgggcgttaatcaaagaggcgaactgtgtgtgagaggtcctatgattatgtccg1200
gttatgtaaacaatccggaagcgaccaacgccttgattgacaaggatggatggctacatt1260
ctggagacatagcttactgggacgaagacgaacacttcttcatcgttgaccgcctgaagt1320
ctctgattaagtacaaaggctatcaggtggctcccgctgaattggaatccatcttgctcc1380
aacaccccaacatcttcgacgcaggtgtcgcaggtcttcccgacgatgacgccggtgaac1440
ttcccgccgccgttgttgttttggagcacggaaagacgatgacggaaaaagagatcgtgg1500
attacgtcgccagtcaagtaacaaccgcgaaaaagttgcgcggaggagttgtgtttgtgg1560
acgaagtaccgaaaggtcttaccggaaaactcgacgcaagaaaaatcagagagatcctca1620
taaaggccaagaagggcggaaagatcgccgtgtaa1655
<210>2
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>2
agaagctatgaaaatcgata20
<210>3
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>3
atcatagagtataaagagaa20
- 还没有人留言评论。精彩留言会获得点赞!