一种生产左旋龙脑的重组酵母工程菌的制作方法
1.本发明涉及基因工程及发酵工程领域,具体涉及一种生产天然冰片成冰片的基因工程菌及其构建方法与应用。
背景技术:
2.冰片,又称龙脑,双环单萜类化合物,根据旋光性分为右旋龙脑和左旋龙脑,右旋龙脑又称天然冰片,植物来源主要是樟科植物龙脑樟(cinnamomum camphora(l.)presl)和梅片树(cinnamomum burmanni(nees et t.nees)blume);左旋龙脑又称艾片,主要植物来源是菊科植物艾纳香(blumea blasamifera(l.)dc.)。在薰衣草、乳香、松树、冷杉和香茅油、乳香中也含有龙脑。
3.龙脑是珍贵的传统中药材和高级香料,广泛用于医药、食品、日化、农药等领域,目前制药企业和药厂的大多数成药原料和配方倾向于使用价廉易得的合成冰片,但是合成冰片除龙脑以外还含有有毒的异龙脑,在冰片植物来源上,研究者采用植物提取法,但是植物中天然化合物含量少,且提取率,植物生长周期长和种植面积受限等限制了天然冰片的获取。利用合成生物学策略生产龙脑和其它单萜类化合物及其衍生物是可行的思路。研究发现,单萜类化合物的生物合成均来源于共同的前体香叶基焦磷酸(geranyl diphosphate,gpp),在植物体内,乙酰辅酶a经甲羟戊酸(mevalonate,mva)途径和4-磷酸-2-甲基赤藓糖(2-c-methyl-d-erythritol-4-phosphate,mep)途径生成异戊烯基焦磷酸(isopentenyl diphosphate,ipp)及其同分异构体二甲基烯丙基焦磷酸(dimethylallyldiphosphate,dmapp),在牻牛儿基焦磷酸单萜合酶的催化下,ipp和dmapp缩合生成gpp,最终在单萜合酶的作用下形成种类繁多的单萜类化合物(o'brien t e,bertolani s j,zhang y,et al.predicting productive binding modes for substrates and carbocation intermediates in terpene synthases-bornyl diphosphate synthase as a representative case[j].acs catal,2018,8:3322.)。张超等(cn110669713a)公开了一种高产单萜类成分的酵母工程菌,并在酿酒酵母中重构柠檬烯合酶的生物合成途径,能够异源合成d-柠檬烯,d-柠檬烯的产量27.3mg/l;周萍萍等(cn111411101a)利用顺式芳樟醇合成酶t67omclis,构建芳樟醇生物合成途径,通过过度表达整个mva通路和引入一个具有降低fpp形成能力的erg20
f96w/n127w
变体,使得前体gpp的供应得到加强,最终菌株在摇瓶中产生53.14mg/l芳樟醇培养物。可见,单萜工程菌的构建使单萜产量得到提高,但是冰片相关的工程菌还没有报道。
技术实现要素:
[0004]
本发明的一方面在于提供一种生产左旋龙脑的酵母工程菌(重组菌),所述重组菌为体内含有或表达艾纳香单萜合酶bbtps3的酵母菌。本发明所述的艾纳香单萜合酶bbtps3在申请人的在先专利(cn201910775971.6)中已有公开,其为来源于艾纳香(blumea blasamifera(l.)dc.),可用于合成或制备左旋龙脑的单萜合酶,该专利的全部内容被引入
本技术并作为本技术的一部分。
[0005]
另一方面,所述酵母菌含有经过密码子优化的艾纳香单萜合酶bbtps3基因。本发明根据酿酒酵母密码子偏好性进行改进,改进的方法是本领域的常规技术手段,例如,根据本领域已知的相关基因的序列并参照酿酒酵母密码子偏好性进行人工合成(酿酒酵母密码子偏好性设计可参考http://www.kazusa.or.jp/codon/cgi-bin/showcodon.cgi?species=493所公开的信息进行),任何依据酿酒酵母密码子偏好而设计得到的上述基因,均可以用于本发明。
[0006]
优选的,本发明经过密码子优化的bbtps3编码核酸如seq id no:1所示。
[0007]
另一方面,本发明还包括对关键酶bbtps3进行蛋白修饰。将bbtps3的n端氨基酸截短,形成3个截短的蛋白(下文称t14-bbtps3,t18-bbtps3,t38-bbtps3),发现t14-bbtps3和t18-bbtps3截短体蛋白的左旋龙脑产量得到了明显的提高。本发明所述t14-bbtps3、t18-bbtps3、t38-bbtps3的编码核酸如seq id no:2、seq id no:3和seq id no:4所示。
[0008]
另一方面,在截短体蛋白的起始密码子atg前面增加通用kozak序列(seq id no:5),得到了3个新的截短体(下文称t14-bbtps3k,t18-bbtps3k,t38-bbtps3k),发现没有提升左旋龙脑产量,随后尝试酵母特异性kozak序列(seq id no:6),得到1个新的截短体(下文称t14-bbtps3k2),进一步提升右旋龙脑产量。
[0009]
另一方面,本发明在以上实验的基础上,考察柔性连接肽对产量的影响,选取5个柔性连接肽进行考察,分别是“gggs”,“gsg”,“yrsqi”,“vipfis”,“wrfspklq”,构建erg20
f96w-n127w
(又称为erg20ww,序列为seq id no:7)在前,bbtps3-14在后的融合蛋白(下文称erg20ww-gggs-t14-bbtps3k2(seq id no:8),erg20ww-gsg-t14-bbtps3k2(seq id no:9),erg20ww-yrsqi-t14-bbtps3k2(seq id no:10),erg20ww-vipfis-t14-bbtps3k2(seq id no:11),erg20ww-wrfspklq-t14-bbtps3k2(seq id no:12)、erg20
f96w
,
n127w-gggs-t14-bbtps3k2-2(seq id no:13);或者构建erg20ww在后,bbtps3-14在前的融合蛋白(下文称t14-bbtps3k2-gggs-erg20
f96w,n127w-2(seq id no:14)的融合蛋白进一步提升左旋龙脑产量。
[0010]
在某些实施例中,本发明所述体内含有或表达艾纳香单萜合酶或艾纳香单萜合酶bbtps3融合蛋白为将所述艾纳香单萜合酶的编码核酸或所述艾纳香单萜合酶bbtps3融合蛋白的编码核酸导入所述酵母菌;和/或,所述将艾纳香单萜合酶的编码核酸导入所述酵母菌为将含有所述艾纳香单萜合酶的编码核酸表达盒导入所述酵母菌;所述将艾纳香单萜合酶融合蛋白的编码核酸导入所述酵母菌为将含有所述艾纳香单萜合酶融合蛋白的编码核酸表达盒导入所述酵母菌;和/或,所述含有艾纳香单萜合酶的编码核酸表达盒通过表达所述艾纳香单萜合酶的编码核酸表达盒的载体导入所述酵母菌;优选通过质粒形式导入所述酵母菌。
[0011]
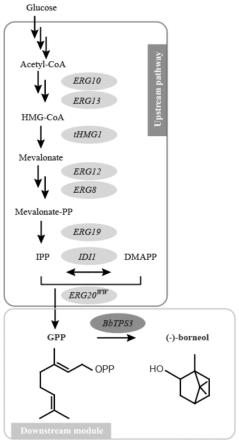
在某些实施例中,本发明通过底盘菌的优化,以提高前体物质gpp的供给(以下简称m/d)。例如,本发明所述的酵母菌包含来自于酵母甲羟戊酸途径中相关的内源性基因,如乙酰乙酰辅酶a转移酶基因erg10、hmg-coa合酶基因erg13、hmg辅酶a还原酶基因thmg1、磷酸甲羟戊酸激酶基因erg8、甲羟戊酸焦磷酸脱羧酶基因erg19、法尼基焦磷酸合酶基因erg20、甲羟戊酸激酶基因erg12、异戊烯焦磷酸异构酶基因idi1。在某些优选的实施例中,本发明使用的erg20为突变体erg20
f96w/n127w
。
[0012]
在某些实施例中,本发明所述酵母为酿酒酵母(saccharomyces cerevisiae);和/或所述酿酒酵母为cen.pk2-1d,基因型为matα,ura3-52,trp1-289,leu2-3112,his3δ1,mal2-8c,suc2。
[0013]
本发明的第五方面,提供了一种生产左旋龙脑的方法,所述方法包括:
[0014]
将生产左旋龙脑的重组酵母菌接种到培养基中,进行发酵,提取分离发酵后的菌液,得到目标产物左旋龙脑,其中,所述重组酵母菌能发酵生产gpp。
附图说明
[0015]
图1为酿酒酵母中gpp和左旋龙脑的生物合成途径示意图;
[0016]
图2为改造的底盘菌菌株中香叶醇产量图,其中a为菌株m/d的生长曲线;b为菌株m/d香叶醇产量;
[0017]
图3为左旋龙脑的gc-ms定性检测图,其中a为左旋龙脑的气象色谱检测结果;b为左旋龙脑的质谱鉴定结果;
[0018]
图4a为艾纳香单萜合酶bbtps3不同n端截短方式对左旋龙脑产量的影响(md-b3、md-b5和md-b7),以及不同kozak序列对左旋龙脑产量的影响(md-b4、md-b6、md-b8和md-b9);
[0019]
图4b为不同连接肽的融合蛋白对左旋龙脑产量的影响。
具体实施方式
[0020]
实验材料
[0021]
表1载体和菌株信息
[0022][0023]
表2引物信息
[0024]
[0025]
[0026][0027]
真核质粒构建(pesc-leu)
[0028]
bbtps3-f’aggagaaaaaaccccgatggttggtttccagaagcactc
[0029]
bbtps3-r’gtgagtcgtattacggggtctttggcttcaacaacaaggag
[0030]
表3蛋白改造引物信息
[0031][0032]
主要试剂:
[0033]
gene jet gel extraction kit(美国thermo scientific公司);e.z.n.atm plasmid mini kit i(美国omega bio-tek公司);peasy-uni seamless cloning and assembly kit(北京全式金生物技术有限公司);2
×
easytaq pcr supermix(+dye)(北京全式金生物技术有限公司);easypure genomic dna kit(北京全式金生物技术有限公司);
phusion high-fidelity master mix(美国neb公司);bsai(美国neb公司);bamhi-hf(美国neb公司);t4 dna ligase美国neb公司;sd-ura酵母培养基(北京泛基诺科技有限公司);sd-leu-ura酵母培养基(北京泛基诺科技有限公司);frozen-ez yeast transformation ii kit
tm
(zymo research生物科技公司;
[0034]
ypd固体平板:1%酵母膏+2%蛋白胨+2%葡萄糖+1.5%琼脂;不加琼脂则成为相应液体培养基(ypd液体培养基);
[0035]
ypl诱导培养基:1%酵母膏+2%蛋白胨+2%半乳糖;
[0036]
sd-ura固体平板:sd-ura+2%葡萄糖+2%琼脂;不加琼脂则成为相应液体培养基(sd-ura液体培养基);
[0037]
sd-ura-leu固体平板:sd-ura-leu+2%葡萄糖+2%琼脂;不加琼脂则成为相应液体培养基(sd-ura-leu液体培养基)。
[0038]
实施例1重组酵母底盘菌的构建
[0039]
以增加左旋龙脑的前体物质gpp为指标,对酿酒酵母菌cen.pk2-1d进行改造,改造方法参考文献(jianga g z,yaoa m d,wanga y,et al.manipulation of ges and erg20 for geraniol overproduction in saccharomyces cerevisiae[j].metabolic engineering,2017,41:57-66.)(xiao-jing g,wen-hai x,ying w,et al.metabolic engineering of saccharomyces cerevisiae for 7-dehydrocholesterol overproduction[j].biotechnology for biofuels,2018,11(1):192.)。(本发明重组酵母底盘菌信息见表1)。
[0040]
1、酵母菌株的构建
[0041]
本研究中使用的初始菌株为cen.pk2-1d来自酿酒酵母(表1)。所有mva通路的内源性基因erg10、erg13、thmg1、erg12、erg8、erg19、idi1、erg20均来自cen.pk2-1d基因组dna。本研究中使用的erg20突变体erg20
f96w/n127w
被报道具有更高的单萜生产效率。采用m2s整合方法将基因表达盒整合到酵母染色体中(li s,ding w,zhang x,et al.development of a modularized two-step(m2s)chromosome integration technique for integration of multiple transcription units in saccharomyces cerevisiae[j].biotechnology for biofuels,2016,30(1):232-243.)。简单描述为通过添加bsai酶切位点扩增erg10和erg13,并将头对头启动子(pgal1-pgal10)连接到终止载体t1-(tpi1-pgi1)中,生成质粒t1-(erg10-erg13)。将两个终止子插入质粒中,分别在终止子设计同源臂l1和l2。同样,t2-(thmg1-thmg1)、t3-(thmg1-erg12)、t4-(erg8-erg19)和t5-(idi1-erg20
f96w-n127w
)分别具有特异的同源臂l2和l3、l3和l4、l4和l5、l5和l6。
[0042]
分别扩增每个带有同源臂的表达模块。本次将外源基因构建在酿酒酵母基因组15位点(yprcδ15),使用ura筛选标记。从cen.pk2-1d基因组dna中扩增得到上游同源臂yprcδ15-up;从pesc-ura载体中扩增包含启动子的ura3模块;从t1载体中扩增得到l1。将3个模块进行overlap pcr(重叠pcr)组装形成选择标记模块yprcδ15up-ura3-l1。从cen.pk2-1d基因组dna中扩增得到下游同源臂yprcδ15down,从t5载体中扩增得到l6,然后组合生成下游同源臂模块l6-yprcδ15。
[0043]
将所有模块均电转入cen.pk2-1d用于组装和整合。转化产物滴在缺陷型sd-ura固体平板中央,用涂布器涂布均匀,直至涂布的全部菌液吸收完全,置于30℃恒温箱中倒立培
养2-3d。挑取单菌落测序验证,得到阳性菌株md(附图1)。
[0044]
2、发酵
[0045]
为了测定md菌株产gpp的能力,对菌株进行发酵检测。详细过程为:
[0046]
(1)挑取sd-ura固体平板上长出来的md的单菌落,置于10ml sd-ura液体培养基中,30℃200rpm 48h;
[0047]
(2)室温5000g 5min离心收集菌体,转接入10ml ypl诱导培养基,30℃200rpm诱导培养48h,得到发酵产物。
[0048]
3、发酵产物萃取
[0049]
目标成分为萜类化合物,脂溶性,易溶于乙酸乙酯,因此选取乙酸乙酯为溶剂萃取发酵产物,得到目标化合物。其中,所述萃取步骤如下:
[0050]
(1)发酵结束后,吸取5ml发酵液,加入等体积乙酸乙酯超声萃取1h,期间每个10分钟上下颠倒混匀,并加入冰使超声温度维持在10℃,以免产物挥发;
[0051]
(2)13,000g离心10分钟,吸取上层有机相,加入适量的无水硫酸钠(120℃烘干30min),边加边摇,除去萃取液的水分;
[0052]
(3)转移至液相小瓶中,密封,用于gc-ms检测。
[0053]
4、发酵产物gc-ms检测
[0054]
gc-ms分析条件为:色谱柱为tr-5ms(30m
×
0.25mm),取1μl样品进样,无分流模式下,50℃保持2min,5℃
·
min-1
升至230℃,保持5min,10℃
·
min-1
升至300℃,保持2min;进样口温度280℃,传输线温度280℃,电子能量70ev,对样品进行50-300m/z范围扫描。以geraniol标准品作为对照,制作标准曲线,并对样品中geraniol含量进行测定。
[0055]
在md菌株中,检测到其geraniol的产量为12.52mg
·
l-1
。(附图2)。
[0056]
实施例2 bbtps3真核表达载体的构建
[0057]
将从艾纳香中克隆得到的单萜合酶bbtps3(参见在先中国专利cn201910775971.6,其所有内容被引入本专利中)根据酿酒酵母密码子偏好性进行改进,改进的方法是本领域的常规技术手段。示例性的单萜合酶基因改进后的密码子序列如seq id no:1所示。将获得上述改进的单萜合酶基因,设计同源臂引物,通过同源重组的方式将其构建至真核表达载体pesc-leu的bamhi位点。具体操作如下:
[0058]
(1)设计带有bamhi位点同源臂的引物bbtps3-f’和bbtps3-r’将单萜合酶bbtps3从pet32a::bbtps3质粒(中国专利cn201910775971.6)中进行pcr扩增,回收并进行纯化获得纯化pcr产物。其中,所述引物序列见表2(下划线所示序列为载体同源区):
[0059]
(2)取pesc-leu载体(agilent technologies公司),用限制性内切酶bamhi进行酶切,回收线性化的载体骨架。
[0060]
(3)取步骤1)得到的纯化pcr产物,按北京全式金生物技术有限公司peasy-uni seamless cloning and assembly kit说明书操作,将其克隆至步骤2)的线性化的载体骨架,得到重组质粒pesc-leu::bbtps3。
[0061]
将重组质粒pesc-leu::bbtps3转化至大肠杆菌trans5α感受态细胞,涂布lb平板,并阳性克隆pcr鉴定(购自北京全式金生物技术有限公司),得到pesc-leu::bbtps3重组菌,用e.z.n.atm plasmid mini kit i(omega bio-tek公司)试剂盒提取pesc-leu::bbtps3重组质粒。
[0062]
实施例3左旋龙脑的发酵
[0063]
将构建好的pesc-leu::bbtps3重组质粒转入md菌株,进行发酵检测左旋龙脑的产量,具体操作如下:
[0064]
1.md酵母感受态制备
[0065]
采用zymo research frozen-ez yeast transformation ii试剂盒做酵母感受态细胞:
[0066]
(1)从sd-ura固体平板上挑取新活化的md酵母菌单菌落,接种于10ml sd-ura液体培养基中,30℃下振荡培养至od600=0.8-1.0左右;
[0067]
(2)室温,500g离心4min,去上清;
[0068]
(3)加入10ml frozen-ez solution 1悬浮菌体,室温,500g离心4min,去上清;
[0069]
(4)加入1ml frozen-ez solution 2悬浮菌体,得到by-mono酵母感受态细胞,分装至灭菌的1.5ml ep管中,每管50μl;
[0070]
(5)缓慢降温至-70℃(4℃,1h;-20℃,1h;-40℃,1h;-70℃保存),禁止用液氮速冻感受态细胞。
[0071]
2.重组质粒pesc-leu::bbtps3转化至md酵母感受态细胞
[0072]
(1)取0.2-1μg重组质粒pesc-leu::bbtps3(少于5μl)与50μl by-mono酵母感受态细胞混合;
[0073]
(2)加入500μl frozen-ez solution 3,剧烈混匀;
[0074]
(3)30℃孵育1-2h,期间在此混匀2-3次;
[0075]
(4)取50-150μl孵育的菌液,涂布sd-ura-leu固体平板上,晾干后,置于30℃倒置培养48h,得到转入重组质粒pesc-leu::bbtps3的重组酵母,将其命名为md-b1。进行发酵,左旋龙脑产物检测见实施例1。发酵而得的初始菌株中含有1.24mg
·
l-1
的左旋龙脑(附图3)。
[0076]
实施例4左旋龙脑高产模块的蛋白结构优化
[0077]
本发明中将获得的酵母密码子优化的bbtps3继续对其进行了进一步修饰。我们根据chlorop1.1(chlorop(http://www.cbs.dtu.dk/services/chlorop/))预测的结果显示bbtps3没有转运肽,但是仍有转运肽长度的预测,以此为参考进行截短试验,我们设计了三个截短位点e14,r18和n38进行截短,分别命名为t14-bbtps3,t18-bbtps3和t38-bbtps3。
[0078]
将蛋白截短质粒pesc-leu::t14-bbtps3,pesc-leu::t18-bbtps3和pesc-leu::t38-bbtps3转化酵母感受态细胞,获得菌株md-b3,md-b5,md-b7,进行发酵测定冰片的产量(图3)。另一方面,在截短蛋白基础上,bbtps3选取常用kozak序列“gccacc”,在不同截短的质粒版本中构建,设计特异性引物对bbtps3进行pcr扩增(表3),构建添加kozak序列的质粒pesc-leu::bbtps3k,pesc-leu::t14-bbtps3k,pesc-leu::t18-bbtps3k和pesc-leu::t38-bbtps3k。将构好的截短质粒转化酵母感受态细胞,获得菌株md-b2,md-b4,md-b6,md-b8,进一步测定冰片的产量。
[0079]
通过对比发现,酵母特异性kozak序列“aaaaaa”对冰片产量的提高更多,所以将bbtps3中冰片产量最高的截短质粒pesc-leu::t14-bbtps3构建加有酵母特异性kozak序列“aaaaaa”的版本,得到质粒pesc-leu::t14-bbtps3k2,获得菌株md-b9。
[0080]
具体操作如下:
[0081]
(1)设计截短引物,见表3,将单萜合酶bbtps3从pet32a::bbtps3质粒中进行pcr扩增,回收并进行纯化获得纯化pcr产物。其中,所述引物序列见表3(下划线所示序列为载体同源区):
[0082]
(2)取pesc-leu载体(agilent technologies公司),用限制性内切酶bamhi进行酶切,回收线性化的载体骨架。
[0083]
(3)取步骤1)得到的纯化pcr产物,按北京全式金生物技术有限公司peasy-uni seamless cloning and assembly kit说明书操作,将其克隆至步骤2)的线性化的载体骨架,得到重组质粒pesc-leu::bbtps3。
[0084]
将重组质粒pesc-leu::t14-bbtps3,pesc-leu::t18-bbtps3,pesc-leu::t38-bbtps3,pesc-leu::bbtps3k,pesc-leu::t14-bbtps3k,pesc-leu::t18-bbtps3k和pesc-leu::t38-bbtps3k转化至大肠杆菌trans5α感受态细胞,涂布lb平板,并阳性克隆pcr鉴定(购自北京全式金生物技术有限公司),得到重组菌,用e.z.n.atm plasmid mini kit i(omega bio-tek公司)试剂盒提取重组质粒。
[0085]
按实施例3的方法将重组质粒转入md菌株,得到新的菌株md-b2,md-b3,md-b4,md-b5,md-b6,md-b7,md-b8,md-b9进行发酵萃取,产量如附图4a所示。结果发现截短体md-b3和md-b5融合蛋白的左旋龙脑产量得到了提高,其中,菌株md-b3和md-b5的产量分别为2.13mg
·
l-1
和1.39mg
·
l-1
。此外,通过酵母特有的kozak序列优化的菌株md-b9产量也得到了提升,为4.87mg
·
l-1
。
[0086]
实施例5蛋白融合提升左旋龙脑产量
[0087]
为了进一步提高冰片的产量,我们进一步分别构建了突变体erg20
f96w/n127w
和bbtps3的融合蛋白,并选取了不同的连接肽连接,优化融合蛋白的结构。
[0088]
选取5条柔性连接肽进行实验,分别是gggs、gsg、wrfspklq、vipfis和yrsqi。bbtps3分别构建pesc-leu::erg20
f96w/n127w-gggs-bbtps3,pesc-leu::erg20
f96w/n127w-gsg-bbtps3,pesc-leu::erg20
f96w/n127w-wrfspklq-bbtps3,pesc-leu::erg20
f96w/n127w-vipfis-bbtps3,pesc-leu::erg20
f96w/n127w-yrsqi-bbtps3,共5个融合蛋白质粒。
[0089]
同时选取pesc-leu::erg20
f96w/n127w-gggs-t14-bbtps3k2考察连接肽核酸序列及蛋白前后顺序的影响,分别构建pesc-leu::erg20
f96w/n127w-gggs-t14-bbtps3k2-2,pesc-leu::t14-bbtps3k2-gggs-erg20
f96w/n127w-2两个融合蛋白质粒,构建示意图如图4b。
[0090]
将这7个质粒转化酵母感受态细胞,获得菌株md-b10,md-b11,md-b12,md-b13,md-b14,md-b15,md-b15进行发酵测定冰片的产量(图4b)。
[0091]
以构建pesc-leu::erg20
f96w/n127w-gggs-bbtps3质粒为例,将突变体erg20
f96w/n127w
构建到bamhi位点,bbtps3构建到sali位点,中间用柔性连接肽“gggs”连接,实验操作步骤如下:
[0092]
1)将质粒pesc-leu用限制性内切酶bamhi和限制性内切酶sali进行双酶切,放置37℃恒温金属浴中孵育1h,反应产物跑琼脂糖凝胶得到单一目的条带,进行切胶回收,得到线性化载体;
[0093]
2)使用带有pesc-leu中bamhi和sali位点同源臂序列的特异性引物(表3)pcr扩增erg20
f96w/n127w
序列,反应产物跑琼脂糖凝胶得到单一目的条带,进行切胶回收,得到erg20
f96w/n127w
dna纯化片段;
[0094]
3)将线性化载体与erg20
f96w/n127w
dna纯化片段利用peasy-uni seamless cloning and assembly kit同源重组试剂盒进行无缝拼接,反应产物转化克隆感受态细胞trans1-t1,进行阳性菌pcr鉴定,送测序,得到质粒pesc-leu::erg20
f96w/n127w
;
[0095]
4)以3)中获得的阳性质粒pesc-leu::erg20
f96w/n127w
为模板,用限制性内切酶sali进行单酶切,放置37℃恒温金属浴中孵育1h,反应产物跑琼脂糖凝胶得到单一目的条带,进行切胶回收,得到线性化载体;
[0096]
5)使用带有pesc-leu中sali位点同源臂序列及带有柔性连接肽“gggs”序列的特异性引物(表3)pcr扩增bbtps3序列,反应产物跑琼脂糖凝胶得到单一目的条带,进行切胶回收,得到bbtps3的dna纯化片段;
[0097]
6)将线性化载体与带有柔性连接肽“gggs”的bbtps3的dna纯化片段利用peasy-uni seamless cloning and assembly kit同源重组试剂盒进行无缝拼接,反应产物转化克隆感受态细胞trans1-t1,进行阳性菌pcr鉴定,送测序,得到质粒pesc-leu::erg20
f96w/n127w-gggs-bbtps3。
[0098]
bbtps3按照此方法分别构建bbtps3融合蛋白质粒pesc-leu::erg20
f96w/n127w-gggs-bbtps3,pesc-leu::erg20
f96w/n127w-gsg-bbtps3,pesc-leu::erg20
f96w/n127w-wrfspklq-bbtps3,pesc-leu::erg20
f96w/n127w-vipfis-bbtps3,pesc-leu::erg20
f96w/n127w-yrsqi-bbtps3,引物见表3。
[0099]
按实例3项下2描述的方法将以上7个构好的融合蛋白质粒转化酵母感受态细胞,获得产左旋龙脑的酵母菌株md-b10,md-b11,md-b12,md-b13,md-b14,md-b15,md-b16,进一步测定冰片的产量,分别为12.41mg
·
l-1
、10.52mg
·
l-1
、12.68mg
·
l-1
、10.81mg
·
l-1
、9.10mg
·
l-1
、12.19mg
·
l-1
、9.16mg
·
l-1
。
[0100]
以上对本发明进行了详述。对于本领域技术人员来说,在不脱离本发明的宗旨和范围,以及无需进行不必要的实验情况下,可在等同参数、浓度和条件下,在较宽范围内实施本发明。虽然本发明给出了特殊的实施例,应该理解为,可以对本发明作进一步的改进。总之,按本发明的原理,本技术欲包括任何变更、用途或对本发明的改进,包括脱离了本技术中已公开范围,而用本领域已知的常规技术进行的改变。按以下附带的权利要求的范围,可以进行一些基本特征的应用。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1