一种甘蔗梢腐病效应因子Fs_00548基因及其应用
一种甘蔗梢腐病效应因子fs_00548基因及其应用
技术领域
1.本发明涉及基因工程技术领域,尤其涉及一种甘蔗梢腐病效应因子fs_00548基因及其应用。
背景技术:
2.甘蔗是我国重要的糖料作物,甘蔗产糖量是全国的产糖总量的重要组成部分
1.,并且甘蔗是重要的能源作物,可以作为乙醇生产原料
2.。甘蔗梢腐病(sugarcane pokkah boeng disease)是一种全球性的真菌性病害,早在1896年就在爪哇被报道。该病主要发生在梢头的嫩叶部位,感病梢头部位叶片扭缠在一起,严重时会出现梢头生长点腐烂,幼嫩心叶坏死,整株甘蔗枯死的情况。近年来,蔗区梢腐病发生日益严重,有些感病品种的发病率高达70%~80%,导致甘蔗生产中造成了严重的产量损失(约10~40%)。因此,甘蔗梢腐病已逐渐成为我国甘蔗的主要病害之一。
3.1995年bailey等从fusarium oxysporum的培养物中纯化出能诱导植物坏死和产生乙烯的24kda蛋白(necrosis
‑
and ethylene
‑
inducing peptide 1
‑
like proteins,nep1
‑
like proteins)。随后发现在植物病原菌中这类蛋白都含有一个保守结构域npp1(necrosis
‑
inducing phytophthora protein)。staats等通过实验验证这类蛋白只可以在双子叶植物上诱导坏死,不能在单子叶植物叶片上诱导坏死。随后,odalys等克隆了病原菌moniliophthora perniciosa编码的2种nep(necrosis and ethylene
‑
inducing proteins)基因,均可以在烟草细胞上诱导细胞死亡。zafer等
8.从核盘菌sclerotinia sclerotiorum中克隆到两个基因ssnep1和ssnep2,并证明它们都可以诱导烟草细胞坏死。parthasarathy等克隆了verticillium dahliae中的7个nlp基因,发现只有2个可以诱导本氏烟草叶片死亡。stan等[发现了葡萄霜霉病hyaloperonospora arabidopsidis中的hanlp3是一种新型的mamp,可以诱导拟南芥的免疫反应。chen等克隆了辣椒疫霉病原菌phytophthora capsic的10个nlps基因,但是只有三个基因可以诱导细胞死亡。rebeca等克隆了葡萄灰霉病原菌diplodia seriata中的4个nlp基因,并成功表达了蛋白,发现只有dsernep1和dsernep2可以诱导细胞坏死。
[0004]
npp1类基因根据结构域中半胱氨酸含量分为2类,含有2个半胱氨酸为ⅰ型,含有4个半胱氨酸为ⅱ型,ⅰ型npp1分布于真菌、卵菌和细菌中,ⅱ型npp1仅分布于真菌和细菌中。所以真菌中既有ⅰ型npp1,也有ⅱ型npp1,如稻瘟菌magnaporthe grisea基因组中编码ⅰ型和ⅱ型npp1的基因各有两个
[13]
。npp1有一个7肽的保守结构域“ghrhdwen”。feng等通过点突变的方法鉴定了其中的活性位点d
112
,h
120
,d
123
,和e
125
。zafer等发现核盘菌sclerotinia sclerotiorum中的两个nep基因诱导细胞坏死依赖于ca
2+
信号通路和环腺苷酸通路。teh等发明发现茉莉酸和水杨酸可以增加病原菌ganoderma boninense中gbnep基因的转录丰度,并且也发现gbnep诱导细胞坏死可能依赖于ca
2+
信号通路。
[0005]
甘蔗镰孢菌fusarium sacchari(f.sacchari)引起的甘蔗梢腐病严重影响了甘蔗产量和质量。解析病原菌的致病机理,对指导甘蔗抗病育种和绿色防控具有重要意义。发明
发现nlps(nep1
‑
like proteins)类基因在真菌侵染及定殖过程中发挥重要作用。
[0006]
目前,缺乏一种可以诱发植物自身免疫的真菌肽的甘蔗梢腐病效应因子fs_00548基因及其应用。
技术实现要素:
[0007]
针对现有技术的不足,本发明提供了一种可以诱发植物自身免疫的真菌肽的甘蔗梢腐病效应因子fs_00548基因及其应用。
[0008]
为了解决现有技术的问题,本发明提供了如下技术方案:本发明的一种甘蔗梢腐病效应因子fs_00548基因,来自普通甘蔗梢腐病病原菌,所述的甘蔗梢腐病效应因子fs_00548具有如seq id no.1所示的核苷酸序列。
[0009]
本发明所述的甘蔗梢腐病效应因子fs_00548基因编码的蛋白质,所述的蛋白质具有如seq id no.2所示的氨基酸序列。
[0010]
本发明所述的甘蔗梢腐病效应因子fs_00548基因编码的蛋白质的信号肽具有分泌活性。
[0011]
本发明用于克隆所述甘蔗梢腐病效应因子fs_00548基因序列的引物对,其特征在于:所述的引物对为pvx
‑
gfp引物对、pvx
‑
bax引物对、pvx
‑
fs_00548引物对、pvx
‑
fs_03159引物对、pvx
‑
fs_06646引物对、pvx
‑
fs_06646引物对、pvx
‑
fs_11062引物对、pvx
‑
fs_00548
△
sp引物对、psuc2
‑
fs_00548sp引物对、psuc2
‑
fs_03159sp引物对、psuc2
‑
fs_11062sp引物对、nb_ef
‑
1引物对、fv_actin引物对和fs_00548
‑
rt引物对;
[0012]
所述的pvx
‑
gfp的正向引物具有如seq id no.3所示的核苷酸序列;
[0013]
所述的pvx
‑
gfp的反向引物具有如seq id no.4所示的核苷酸序列。
[0014]
所述的pvx
‑
bax的正向引物具有如seq id no.5所示的核苷酸序列;
[0015]
所述的pvx
‑
bax的反向引物具有如seq id no.6所示的核苷酸序列;
[0016]
所述的pvx
‑
fs_00548的正向引物具有如seq id no.7所示的核苷酸序列;
[0017]
所述的pvx
‑
fs_00548的反向引物具有如seq id no.8所示的核苷酸序列;
[0018]
所述的pvx
‑
fs_03159的正向引物具有如seq id no.9所示的核苷酸序列;
[0019]
所述的pvx
‑
fs_03159的反向引物具有如seq id no.10所示的核苷酸序列;
[0020]
所述的pvx
‑
fs_06646的正向引物具有如seq id no.11所示的核苷酸序列;
[0021]
所述的pvx
‑
fs_06646的反向引物具有如seq id no.12所示的核苷酸序列;
[0022]
所述的pvx
‑
fs_11062的正向引物具有如seq id no.13所示的核苷酸序列;
[0023]
所述的pvx
‑
fs_11062的反向引物具有如seq id no.14所示的核苷酸序列;
[0024]
所述的pvx
‑
fs_00548
△
sp的正向引物具有如seq id no.15所示的核苷酸序列;
[0025]
所述的pvx
‑
fs_00548
△
sp的反向引物具有如seq id no.16所示的核苷酸序列;
[0026]
所述的psuc2
‑
fs_00548sp的正向引物具有如seq id no.17所示的核苷酸序列;
[0027]
所述的psuc2
‑
fs_00548sp的反向引物具有如seq id no.18所示的核苷酸序列;
[0028]
所述的psuc2
‑
fs_03159sp的正向引物具有如seq id no.19所示的核苷酸序列;
[0029]
所述的psuc2
‑
fs_03159sp的反向引物具有如seq id no.20所示的核苷酸序列;
[0030]
所述的psuc2
‑
fs_11062sp的正向引物具有如seq id no.21所示的核苷酸序列;
[0031]
所述的psuc2
‑
fs_11062sp的反向引物具有如seq id no.22所示的核苷酸序列;
[0032]
所述的nb_ef
‑
1的正向引物具有如seq id no.23所示的核苷酸序列;
[0033]
所述的nb_ef
‑
1的反向引物具有如seq id no.24所示的核苷酸序列;
[0034]
所述的fv_actin的正向引物具有如seq id no.25所示的核苷酸序列;
[0035]
所述的fv_actin的反向引物具有如seq id no.26所示的核苷酸序列;
[0036]
所述的fs_00548
‑
rt的正向引物具有如seq id no.27所示的核苷酸序列;
[0037]
所述的fs_00548
‑
rt的反向引物具有如seq id no.28所示的核苷酸序列;
[0038]
本发明所述的fs_00548基因在制备抗甘蔗梢腐病真菌肽药物中的应用。
[0039]
进一步地,所述的fs00548基因在本氏烟草叶片上诱导细胞死亡,qrt
‑
pcr结果显示该基因在整个侵染过程中都转录表达。
[0040]
有益效果:本发明可以诱发植物自身免疫的真菌肽,在致病甘蔗镰孢菌全基因组测序(尚未公布)基础上,分析其中的分泌蛋白,通过blastp找到4个甘蔗梢腐病效应因子fs_00548,fs_03159,fs_06646,fs_11062。通过烟草叶片的瞬时表达,发现只有fs_00548基因可以在本氏烟草叶片上诱导细胞死亡,并通过rt
‑
pcr和western bot对这些基因在侵染烟草后的转录的翻译情况进行了验证。qrt
‑
pcr结果显示该基因在整个侵染过程中都转录表达。根据结果初步证明,fs_00548可能是一个nep1
‑
like基因,n端的信号肽具有分泌活性,可以在烟草细胞上诱导细胞坏死。
[0041]
与现有技术相比,本发明具有如下优点:
[0042]
(1)本发明从病原菌基因组入手,寻找含有保守npp1结构域的基因,并初步验证初该基因可以诱发烟草叶片的免疫反应,说明该蛋白可能是一种可以诱导植物免疫的激发子,为甘蔗病害的绿色防控提供参考。
[0043]
(2)本发明发现了一种在侵染过程中高表达的npp1基因,初步分析其可能在真菌与植物互作中发挥重要功能,为进一步探索病原菌与植物互作提供依据。
附图说明
[0044]
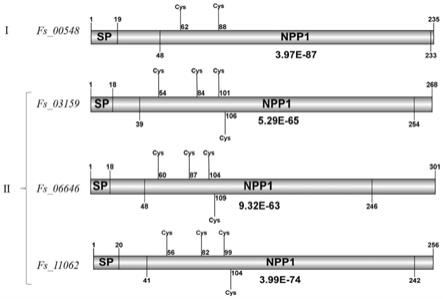
图1为本发明的4个含有npp1结构域的基因结构图;
[0045]
通过blastp在甘蔗梢腐病病原菌fusarium sacchari中找到了4个含有npp1结构域的基因,运用ibs 1.0软件进行了基因结构绘图展示,并标注出基因的信号肽区域,结构域具体位置以及其中的半胱氨酸含量及位置信息。
[0046]
图2为本发明的蛋白质序列比对分析图;
[0047]
运用dnaman软件,进行蛋白质序列比对分析,发现在其结构域中含有保守的肽段“ghrhdwen”,并展示了保守区域的起止位置。fs 03159的肽段与其他不同。
[0048]
图3为本发明的重组农杆菌在烟草叶片进行渗透注射的瞬时表达图;
[0049]
将重组农杆菌在烟草叶片背面进行渗透注射,每个位点做好标记。每天观察叶片状态,7d后拍照记录,然后将叶片放入无水乙醇中脱色,拍照记录。侵染2d后每个位点采样,提取rna,反转录,进行rt
‑
pcr验证基因是否发生转录;提取总蛋白,进行western blot实验,验证蛋白翻译情况。
[0050]
图4为本发明的运用酵母信号肽分泌功能验证方法图;
[0051]
运用酵母信号肽分泌功能验证方法,对候选基因的信号肽分泌功能进行验证,以大豆疫霉中的avr1b为阳性对照;稻瘟菌中的非分泌蛋白mg87作为阴性对照,结果显示fs_
00548,fs_03159的信号肽具有分泌活性。
[0052]
图5为本发明的基因在真菌侵染过程中的表达情况图;
[0053]
通过qrt
‑
pcr方法,验证该基因在真菌侵染过程中的表达情况。以菌丝中该基因的表达量作为参照,fs_00548在甘蔗镰孢菌侵染甘蔗的12h出现显著的诱导表达,直到72h达到表达高峰(高达48倍)。随后表达量逐步下降。
具体实施方式
[0054]
为了使本发明实现的技术手段、创作特征、达成目的与功效易于明白了解,下面结合具体实施方式,进一步阐述本发明。
[0055]
实施例1
[0056]
试验例1
[0057]
1材料与方法
[0058]
1.1实验材料
[0059]
供试植物:甘蔗品种中蔗1号,本实验室提供,温室大棚中桶栽培养,五叶展齐备用,该品种对f.sacchari表现为感病。本氏烟草,本实验室提供,光照培养箱中培养(28℃光照16h,25℃夜晚8h)。供试菌株:大肠杆菌top10,从上海唯地公司购买,37℃恒温培养;农杆菌gv3101(pjic sa_rep),从上海唯地公司购买,28℃恒温培养;酵母菌株ytk12,由中国农业科学院刘文德老师馈赠,30℃恒温培养。供试载体:pvx病毒载体由西北农林科技大学康振生院士馈赠;载体psuc2,psuc2
‑
mg87,psuc2
‑
avr1b由中国农业科学院刘文德老师馈赠。
[0060]
1.2实验方法
[0061]
1.2.1目的基因序列分析预测基因的保守结构域在conserved domains(https://www.ncbi.nlm.nih.gov/structure/cdd/wrpsb.cgi)和
[0062]
pfam(http://pfam.xfam.org/search)上进行。预测基因编码蛋白的信号肽区域在signalp
[0063]
4.1(www.cbs.dtu.dk/services/signalp/)和
[0064]
smart(http://smart.embl
‑
heidelberg.de/smart/set_mode.cgi?normal=1)上进行。预测基因编码蛋白的跨膜区域在tmhmm(http://www.cbs.dtu.dk/services/tmhmm/)上进行。克隆基因引物设计在primer
‑
blast(https://www.ncbi.nlm.nih.gov/tools/primer
‑
blast/)上进行。qrt
‑
pcr引物设计在idt(https://www.ncbi.nlm.nih.gov/tools/primer
‑
blast/)上进行。primer sequences used in this study如表1所示:正向引物f;反向引物r。
[0065]
表1
[0066][0067][0068]
1.2.2目的基因的克隆与载体构建利用表1中的引物进行pcr扩增,以f.sacchari侵染中蔗1号叶片的rna反转录产物为模板,扩增后构建至t载体,测序验证。pvx载体含有cla
ⅰ‑
sma
ⅰ‑
not
ⅰ‑
salⅰ酶切位点,选取claⅰ和notⅰ酶切位点,保留了ha标签,运用in
‑
fusion的方法进行载体构建。bax基因由上海生工合成。根据软件预测信号肽区段,扩增fs_00548
基因去掉信号肽的区域,构建至pvx载体。根据psuc2t7m13ori(psuc2)载体的ecor
ⅰ‑
xhoⅰ酶切位点,设计引物扩增信号肽区域连接至psuc2载体,psuc2
‑
avr1b作为阳性对照,psuc2
‑
mg87作为阴性对照,转化至酵母菌株ytk12中。
[0069]
1.2.3农杆菌介导的烟草瞬时转化本实验中所有构建的重组pvx载体均通过冻融法转化至农杆菌gv3101中。阳性重组农杆菌用液体lb培养基(50mg
·
ml
‑1kan,20mg
·
ml
‑1rif,50mg
·
ml
‑1gen)摇菌,28℃,220rpm震荡培养24h。4000rpm,5min收集菌体,用10ml 10mm mgcl2洗涤菌液,重复三次,并调节od
600
=0.4,28℃黑暗条件下静置2h。选取一个月大小的烟草叶片,用去掉针头的注射器在叶片背面进行渗透注射,以接种空载体和pvx
‑
gfp的位点作为阴性对照,接种pvx
‑
bax的位点作为阳性对照。每个处理重复3次,接种后的植物置于温室培养,每天观察记录症状变化,7d后拍照记录。
[0070]
1.2.4 western blot由于pvx载体含有一段ha标签蛋白,构建载体时保留了ha标签蛋白序列。将重组载体转化进入农杆菌gv3101中,鉴定出阳性克隆后交给陕西艾优稷公司进行western blot实验。
[0071]
1.2.5信号肽分泌功能验证利用酵母信号肽分泌系统对fs_00548蛋白的信号肽分泌功能进行验证。酵母菌株ytk12,无色氨酸合成基因,无蔗糖转化酶基因;psuc2有色氨酸合成基因,有一个缺失atg及信号肽的蔗糖转化酶基因;cmd
‑
w培养基(cmd
‑
w medium:0.67%yeast n base without amino acids,0.075%tryptophan dropout supplement,2%sucrose,0.1%glucose,and 2%agar),不含色氨酸,以蔗糖和葡萄糖为碳源;ypraa培养基(1%yeast extract,2%peptone,2%raffinose,and 2mg
·
ml
‑
1 antimycin a)不含色氨酸,以棉籽糖为碳源。以未转化的ytk12菌株,空载psuc2和psuc2
‑
mg87为阴性对照,psuc2
‑
avr1b作为阳性对照。如果预测的信号肽具有分泌功能,则可以将蔗糖转化酶分泌至细胞外,使2,3,5
‑
氯化三苯基四氮唑(ttc)还原为红色的1,3,5
‑
三苯甲臜,通过将转化的酵母进行ttc染色可以进一步确定其信号肽的分泌功能。将阳性克隆接种于10ml ypda培养基中,30℃震荡培养2d,12000rpm,1min收集菌体,ddh2o洗涤两次,最终用750μl无菌水重悬菌体。加入250μl 10mmol
·
l
‑1醋酸
‑
醋酸钠缓冲液(ph=4.7),500μl 10%蔗糖溶液,37℃孵育10min;12000rpm离心1min,取100μl上清液加900μl 0.1%ttc,室温静置5min,观察颜色变化,若颜色变为红色说明信号肽具有分泌功能。
[0072]
1.2.6目的基因的表达模式分析甘蔗镰孢菌f.sacchari在pda平板上生长7d,在平板上打出菌饼,接种在中蔗1号的叶片上,保湿24h。分别在接种后0h,12h,24h,48h,72h,120h,144h,216h采样。将上述时间点样品和菌丝,参照takara minibest universal rna extraction kit(takara)试剂盒提取总rna,反转录得到cdna。使用lightcycler 96定量pcr仪器,以反转录得到的cdna为模板,选取基因fusarium verticillioides(f.verticillioides)actin作为内参基因,使用tb green(takara)染料法进行pcr扩增,每个反应生物学重复3次。反应结束后,按照2
‑△△
ct
法分析基因在甘蔗镰孢菌侵染甘蔗不同时间点的相对表达量。
[0073]
2结果与分析
[0074]
2.1序列分析结果
[0075]
fs_00548orf全长708bp,编码235个氨基酸,n端有19aa的信号肽,无跨膜区。fs_03159全长807bp,编码268个氨基酸,n端有18aa的信号肽,无跨膜区。fs_06646全长906bp,
used as negative controls.yeast growth on cmd
‑
w(minus trp)medium confirmed that the vector had transformed into the yeast strain.the growth on ypraa medium and change in color of ttc confirmed the invertase secretion.
[0083]
2.4 fs_00548在甘蔗镰孢菌侵染过程中诱导表达
[0084]
通过提取菌丝以及侵染过程中各个时间点甘蔗叶片的rna,反转录为cdna,以真菌菌丝中的表达量作为对照(相对表达量为1),分析fs_00548基因在甘蔗镰孢菌侵染甘蔗叶片过程中的表达模式。qrt
‑
pcr结果显示:与菌丝相比,fs_00548在甘蔗镰孢菌侵染甘蔗的12h出现显著的诱导表达,直到72h达到表达高峰(高达48倍)。随后表达量逐步下降。图5 expression parttern of fs_00548 in the interaction between pathogen and sugarcane.the asterisks indicate statistical difference relative to mycelium.my:mycelium.hpi:hour past infection.(“﹡”indicates p<0.05,“﹡﹡”indicates p<0.01)
[0085]
3结论与讨论
[0086]
目前在已经公布的众多真菌,细菌,卵菌基因组中都发现了nlp基因,说明这类基因广泛存在各种病原菌中,并且在病原菌与植物互作中发挥重要功能。在1995年,镰刀菌中的npp1基因就已经被报道,随后在核盘菌(sclerotinia sclerotiorum)、芽孢杆菌(bacillus)、大丽轮枝菌(verticillium dahliae)、马铃薯晚疫病菌(phytophthora infestans)、稻瘟病菌(magnaporthe oryzae)等原核和真核病原菌中均发现含有npp1保守结构域基因。本实验中从基因组中分析得到4个含有npp1保守结构域的nep1
‑
like基因,但是只克隆到了3个基因(fs_00548,fs_03159和fs_11062)。通过瞬时表达本氏烟草叶片,发现只有fs_00548可以在烟草叶片上诱导细胞坏死。与fs_00548蛋白的“ghrhdwen”不同,fs_03159蛋白的保守肽段为“ghthdwen”,并且fs_03159蛋白的信号肽具有分泌活性,推测该基因不能诱导坏死可能是由突变引起的,但是需要进一步的氨基酸突变实验验证。parthasarathy等发明发现引起枯萎病的病原菌v.dahliae中含有7个nlp基因,但是只有2个基因可以诱导本氏烟草叶片坏死。chen等发明发现辣椒疫霉病原菌p.capsic中有10个nlp基因,但是只有3个基因可以诱导烟草细胞和辣椒叶片坏死。根据这些结果,同种病原菌的nlp基因可能在真菌侵染植物过程中发挥不同的功能。
[0087]
病原菌的分泌蛋白都可能作为效应蛋白发挥功能,因为效应蛋白通过内质网
‑
高尔基体的分泌途径进行,为确保分泌功能的正常进行,所以需要n端的信号肽发挥分泌功能。由于信号肽可以被分析软件识别,所以才能在软件分析效应蛋白的时候缩小范围。fs_00548,fs_03159和fs_11062序列的n端均有一段信号肽,通过酵母信号肽分泌功能验证系统,发现fs_00548和fs_03159的信号肽具有分泌功能,fs_11062的信号肽区域没有分泌活性,说明fs_00548和fs_03159是典型的分泌蛋白基因。而fs_11062蛋白的信号肽没有分泌活性可能是该基因不能在烟草叶片上诱导细胞坏死的原因之一,需要进一步的实验探究。当fs_00548蛋白缺失信号肽时,发现该基因的诱导坏死功能减弱,说明信号肽区域对其发挥功能具有重要作用,由于信号肽是分泌蛋白的重要组成部分,推测信号肽的缺失可能影响了植物对该蛋白的识别,需要进一步实验探究。
[0088]
本实验中采用侵染甘蔗叶片的方法,通过qrt
‑
pcr发明fs_00548基因在侵染过程中的表达情况。结果显示,fs_00548基因在整个侵染过程中均有表达量,在72h表达量最高,
是菌丝中的48倍,随后表达量持续下降。nlp基因的表达模式以前也报道过,大豆疫霉的ps nlp1在接种植物48h后表达量有显著的上调,致病疫霉pinpp1.1只在侵染的后期才能检测到。致病疫霉接种马铃薯叶片后pitg_10839的表达量随着接种时间的不同呈现不同的上调趋势,且在48h时上调倍数最高,达到350倍左右[25]。病原菌mycosphaerella graminicola中的mg nlp在侵染的末期被检测到高表达。说明这一类基因可能在整个真菌侵染过程中都有表达,但是主要在后期发挥功能。真菌在侵染阶段分泌诱导细胞坏死的蛋白,不仅可以破坏宿主防御系统,帮助病原菌成功定殖甘蔗,并且可能为了使其营养方式从寄生转化为腐生。虽然该类基因在侵染早期也有表达,但是其活性可能被其他效应蛋白抑制。该基因在侵染过程中发挥的具体功能还需要进一步实验探究。
[0089]
尽管已经示出和描述了本发明的实施例,对于本领域的普通技术人员而言,可以理解在不脱离本发明的原理和精神的情况下可以对这些实施例进行多种变化、修改、替换和变型,本发明的范围由所附权利要求及其等同物限定。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1