小麦UDP-Ara变位酶及编码基因与用途
本发明属于生物技术领域,具体涉及一种小麦udp-ara变位酶及编码基因与用途。
背景技术:
阿拉伯糖是植物特异性单糖,是包括多聚糖和糖蛋白在内的多种细胞壁成分的组成部分。该单糖有呋喃糖(arabinofuranose,araf)和吡喃糖(arabinopyranose,arap)两种形式,在植物中主要以araf形式存在。利用udp-ara变位酶(udp-arabinopyranosemutases,uams)的变位酶活性将udp-arap转化为udp-araf是植物中araf合成的唯一途径。因此,udp-ara变位酶在植物细胞壁合成相关过程中起非常重要的作用。
udp-ara变位酶也叫可逆糖基化多肽(reversiblyglycosylatedpolypeptides,rgps),是一类植物特有的蛋白,于1986年首次在胡萝卜细胞悬浮液中被发现,早期的研究主要集中在可逆糖基化多肽的可逆糖基化活性,对该蛋白生物学功能研究甚少。konishi等人于2007年从水稻幼苗中分离出一种能在体外催化udp-arap和udp-araf相互转化的蛋白,将其定名为udp-ara变位酶,并且证明udp-ara变位酶就是之前被报道过的可逆糖基化多肽。根据变位酶活性,udp-ara变位酶家族成员可分为两类。第一类变位酶具有变位酶活性,在不同植物中的数目为2-4个。第二类变位酶不具有变位酶活性,在多数植物中的数目为1个。
目前在许多植物中都鉴定出了udp-ara变位酶。拟南芥中(arabidopsisthaliana)共发现5个udp-ara变位酶基因,分别为atrgp1-atrgp5,其中atrgp1、atrgp2和atrgp5在花、幼苗和发育中的角果中表达量最高,而atrgp3和atrgp4只在角果中被检测到。大麦(hordeumvulgare)中共发现4个udp-ara变位酶基因,hvuam1、hvuam3和hvuam4在大麦组织中广泛分布,但hvuam2基因只在根、叶基、花药和胚乳中检测到。hvuam1和hvuam4在根尖和幼胚中表达量较高,而hvuam3在根尖和幼嫩花药中表达量较高。短柄草(brachypodiumdistachyon)中发现4个udp-ara变位酶基因,bdrgp1、bdrgp2和bdrgp4在不同时期的叶片、茎秆和叶鞘中表达丰度较高,但bdrgp3在不同组织中表达丰度都很低,甚至在叶片中不表达。水稻(oryzasativa)中发现3个udp-ara变位酶基因,其中osuam1和osuam2在花药、柱头、子房、叶片、叶鞘和节间表达水平较高,osuam3仅在生殖相关的组织,特别是花药中表达量较高。以上研究表明,不同类型udp-ara变位酶的组织分布有所不同,说明不同的udp-ara变位酶基因可能参与不同的生理代谢过程。udp-ara变位酶与细胞壁合成密切相关,而保持细胞壁完整性对植物适应逆境胁迫至关重要,所以udp-ara变位酶可能参与植物对逆境胁迫的响应。目前关于udp-ara变位酶基因家族与逆境胁迫关系的研究甚少。通过对水稻的胁迫研究发现,金属铬能诱导3个水稻udp-ara变位酶蛋白在根部表达。对棉花(gossypiumhirsutum)rgp启动子转基因烟草(nicotianatabacum)的研究表明,该启动子会被损伤和脱水胁迫诱导表达。udp-ara变位酶在其它物种应对逆境胁迫过程中的生物学功能研究亟待开展。
小麦(triticumaestivum)是全球范围内的主要粮食作物,挖掘小麦重要基因资源对小麦育种意义重大。本专利发明人长期从事小麦抗病机制方面的研究,在前期筛选小麦成株抗病性相关基因的过程中,得到一个注释为可逆糖基化多肽的表达序列标签(expressedsequencetag,est)片段xz9104_cy32_c235。该基因在条锈菌接种后诱导表达,说明其可能参与了小麦的成株期抗病性。将xz9104_cy32_c235est序列与小麦数据库进行比对,发现该基因所在家族包含四个基因。其中两个基因已被鉴定,分别被定名为targp1和targp2。另外,有关udp-ara变位酶家族基因在小麦中的生物学功能还未有报道。
技术实现要素:
本发明的目的在于提供一种小麦udp-ara变位酶及编码基因与用途。本发明将小麦udp-ara变位酶的编码基因命名为tauam3。本发明克隆了tauam3基因的cdna全长序列和dna全长序列,并对该序列进行了生物信息学分析。利用rt-pcr技术对该基因的组织特异性表达及不同胁迫条件下的表达特征进行了分析,为进一步研究该基因的功能、进而选育抗逆小麦品系提供了理论依据。
具体地,本发明提供一种小麦udp-ara变位酶,所述udp-ara变位酶的氨基酸序列如序列表seqidno.1所示。
具体地,本发明提供所述小麦udp-ara变位酶的编码基因tauam3的核苷酸序列如序列表seqidno.2所示。
基于本发明的技术方案,扩增所述基因tauam3开放阅读框全长的引物设计为:
f:5′atggcgccgctgctcaaggacgagctg3′;
r:5′ctacttccccttgagtttgccg3′。
进一步地,所述基因tauam3开放阅读框全长为1071bp,编码356个氨基酸。
利用graingenes数据库对tauam3进行预测分析,所述基因tauam3位于小麦3bl染色体上。
经序列进行比对分析,所述基因tauam3含有2个内含子,分别长131和88bp。
实时定量pcr分析表明,所述基因tauam3在小麦根、茎、叶和穗部均有表达。
另外,本发明提供所述小麦udp-ara变位酶,或权利要求2-6中任一项所述基因tauam3在激素信号转导途径及逆境胁迫中的用途。
进一步地,植物激素sa、meja、aba和eth及nacl盐胁均能诱导基因tauam3表达udp-ara变位酶。
还有,本发明提供所述小麦udp-ara变位酶,或权利要求2-6中任一项所述基因tauam3在选育抗逆小麦品系中的用途。
与现有技术相比,本发明所述小麦udp-ara变位酶及编码基因,具有下述的有益效果或优点。
本发明利用rt-pcr方法,从小麦中克隆到一个udp-ara变位酶基因tauam3。该基因orf全长1071bp,编码356个氨基酸。同源性分析显示,tauam3与大麦hvuam3具有最高的同源性。tauam3与短柄草、高粱、水稻和玉米同源蛋白有83.2%-87.3%的相似性,与拟南芥atrgp1和蓖麻rcrgp4的同源性分别为82.1%与57.5%,表明udp-ara变位酶基因氨基酸序列在单子叶植物内部相对保守。组织特异性表达分析结果显示,tauam3在检测的小麦组织中都有表达,说明该基因在小麦组织中的分布比较广泛。这与其他物种中同源基因的研究结果类似。另外,tauam3在根中表达丰度最高、在茎和子叶中表达丰度较低的趋势与hvuam3在大麦中分布一致,说明该类基因的组织特异性分布在近源属种中具有较高相似性。
现有研究表明,sa和ja处理会明显增加短柄草叶片中阿拉伯糖的合成,而阿拉伯糖的合成与udp-ara变位酶的表达量直接相关,可以推测sa和ja可能会诱导udp-ara变位酶基因的表达。本发明发现,多种激素处理小麦叶片后,tauam3都有不同程度的诱导表达。sa和meja对小麦tauam3有强烈的诱导作用,并且持续时间较长。aba和eth也能诱导tauam3的上调表达,但是诱导程度小于sa和meja。众所周知,水杨酸作为一种能激活植物抗病防卫反应的重要内源信号分子,与植物抗病性密切相关]。ja参与植物的发育和对生物及非生物胁迫的反应,可以独立调节多种抗菌蛋白的表达,并且能和sa、eth和aba等相互作用,调控植物的防御反应。本发明中,tauam3的转录表达显著受sa、meja、eth和aba的诱导,表明tauam3可能参与了依赖sa、meja、eth和aba的信号途径。另外,高盐胁迫能明显诱导tauam3的表达,暗示tauam3基因可能在植物对盐胁迫响应中发挥重要作用。tauam3可能通过参与依赖sa、meja、eth和aba的信号途径来参与小麦对生物及非生物逆境胁迫的响应。
综上,本发明从小麦中克隆了一个udp-ara变位酶基因tauam3,并对其在不同胁迫条件下的表达特征进行了研究,这不仅丰富了植物udp-ara变位酶研究的资料库,也为进一步揭示tauam3基因在小麦胁迫应答及抗病过程中的具体功能及作用机制奠定了基础。
附图说明
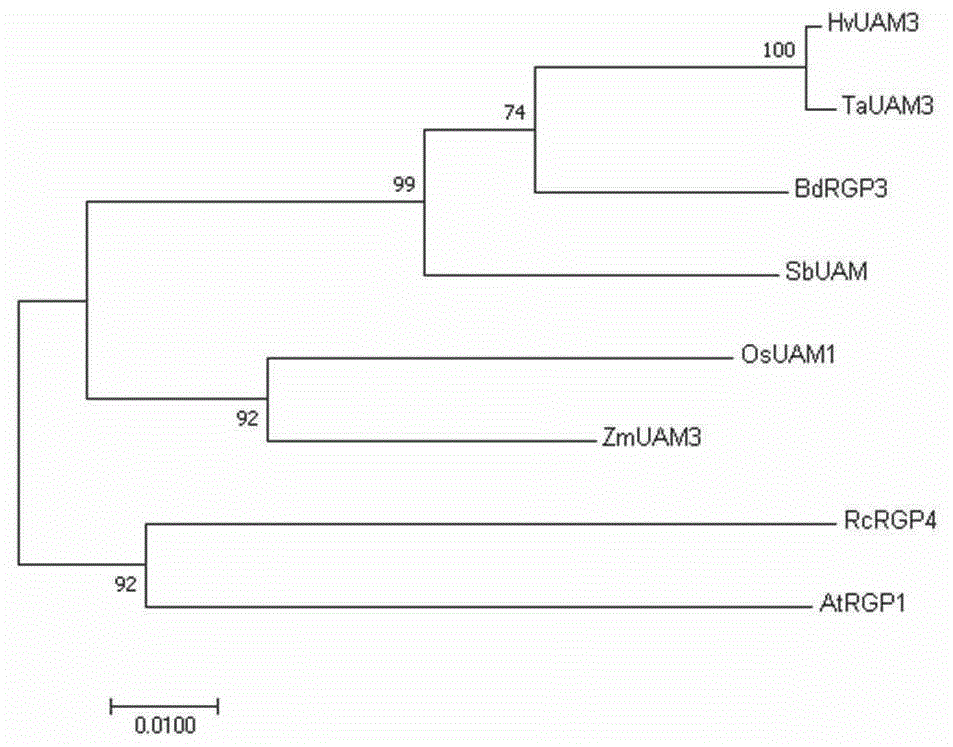
图1为几种植物udp-ara变位酶蛋白同源序列的系统进化树分析。图1中,标尺代表每单位氨基酸的变化,0.01代表两个序列之间1%的差异。
图2为tauam3基因在小麦不同组织器官中的表达。
图3为小麦tauam3基因在激素处理下的表达模式。
图4为小麦tauam3基因在高盐和干旱胁迫下的表达模式。
具体实施方式
下面结合实施例对本发明提供的一种小麦udp-ara变位酶及编码基因与用途进行详细的说明。
1材料与方法
1.1材料
供试小麦品种“水源11”由西北农林科技大学植物病理研究所提供。
1.2方法
1.2.1胁迫处理及取样
小麦幼苗的培育参照康振生等(康振生等.洛夫林10常温致病新菌系的发现[j].西北农学院学报,1984,12(4):18-28.)的方法进行,于光照培养箱(percival,e-41l2)中14-16℃培养。获取小麦的根所用小麦培育方法为,种子用清水萌发后置于hogland营养液中,于光照培养箱中生长,条件同上。成株期小麦于人工气候室中培养,光照强度10000lux,每日光照16h,光照条件下23℃,黑暗条件下13℃。分别于小麦一叶展平期、二叶期和扬花初期剪取小麦子叶、第二叶、茎杆、旗叶及穗部,在小麦一叶展平期剪取小麦的根,液氮速冻后于-80℃保存,用于不同小麦组织中tauam3表达量的检测。对小麦幼苗分别进行植物激素水杨酸(sa)、乙烯(eth)、茉莉酸甲酯(meja)、脱落酸(aba)及nacl和peg6000处理,激素处理及环境胁迫处理的对照样品分别为喷清水处理及正常条件生长的小麦。分别于处理后0、2、6、12和24h剪取处理和对照叶片,液氮速冻后于-80℃保存备用。
1.2.2总rna提取与cdna合成
采用通用植物rna提取试剂盒(华越洋)提取小麦样品的总rna,提取过程中用dna酶ⅰ去除基因组dna。通过琼脂糖凝胶电泳检测rna的完整性,并利用核酸蛋白检测仪(life,nanodroptm1000)测定rna的浓度。用反转录试剂盒(thermoscientific)合成cdna,rna用量为2μg。合成的cdna模板稀释后用于基因的克隆和实时定量pcr分析。
1.2.3小麦tauam3基因的cdna全长克隆
以xz9104_cy32_c235est序列作为种子序列,在ensembleplant网站针对小麦数据库进行比对,查找并获得一条udp-ara变位酶基因的序列。利用ncbi网站的orffinder程序进一步分析所得序列的开放阅读框(openreadingframe,orf)。应用primerpremier5.0软件,对获得的序列设计特异引物,得到1对能有效扩增小麦tauam3orf全长的引物:
f:5′atggcgccgctgctcaaggacgagctg3′;
r:5′ctacttccccttgagtttgccg3′。
将不同生长阶段小麦样品的总rna等量混合,经反转录得到cdna,以其为模板进行pcr扩增。反应体系为:2μl10×taqmastermix(novoprotein),上游引物和下游引物(10μmol/l)各1μl,10倍稀释后的模板cdna2.0μl,补去离子水至总体积20μl。pcr程序为:95℃4min;95℃1min,60℃1min,72℃1min,33个循环;72℃延伸10min;4℃保存。pcr产物回收后克隆至pgem-teasy载体,采用通用引物t7和sp6送交测序。
1.2.4序列分析
利用expasy网站的protparam工具预测编码蛋白的等电点、分子质量、分子式及不稳定指数。利用protscale在线分析蛋白的疏水性。利用signalp-5.0server进行氨基酸序列信号肽分析。利用tmhmmserverv.2.0进行跨膜蛋白结构分析。利用netphos3.1server进行磷酸化位点分析。利用sopma软件预测编码蛋白的二级结构。利用结构域分析软件对编码蛋白的结构域进行分析。利用psort工具进行亚细胞定位预测。利用graingenes数据库进行基因的染色体定位分析。通过ncbi网站进行blastp检索,获得其它植物中与小麦tauam3基因同源性较高的氨基酸序列。利用dnaman对小麦tauam3氨基酸序列与其他植物同源序列进行同源性比对,并利用mega7.0软件构建系统发育树。
1.2.5小麦tauam3基因的dna全长克隆
利用ctab法提取小麦叶片基因组dna。采用orf全长引物进行pcr扩增、克隆并送交测序,方法同1.2.3。
1.2.6实时定量pcr分析
利用primerquesttool在线引物设计软件,根据tauam3全长cdna序列设计特异引物:
f:5′gctgataagggacgagaaggtg3′;
r:5′gtgcccaaaggcgaagatg3′。
以小麦延伸因子基因taef-1a和小麦gapdh基因为内参基因分别设计引物:
f:5′aacacttcaccggaccattca3′;
r:5′cgtccctgccctttgtacac3′;和
f:5′ttcaacatcattccaagcagc3′;
r:5′cgtaacccaaaatgcccttg3′。
应用实时定量pcr仪(bio-rad,cfxconnect)进行基因表达分析。反应体系为:2×sybrqpcrmastermix(vazyme)10μl,上游引物和下游引物(10μmol/l)各0.5μl,30倍稀释后的模板cdna3μl,加灭菌去离子水补足体积至20μl。反应程序为:95℃3min;95℃10s,60℃10s,72℃30s,40个循环。反应结束后分析扩增产物的熔解曲线。每个反应含3次重复,试验设3次生物学重复。每对引物的扩增设未加模板对照,以检验pcr扩增的背景。所有引物进行扩增效率检测,扩增效率在90%-110%的引物符合要求。目标基因在样品中的表达量分别除以相应时间点对照样品的表达量,即得到目标基因的表达量。
2结果与分析
2.1tauam3的克隆与序列分析
以xz9104_cy32_c235est序列为母序列,通过在ensembleplant网站与小麦数据库进行比对,找到该est片段所在基因的全长序列。利用ncbi网站的orffinder程序对此序列进行预测分析,发现该序列含有1个完整的orf区。在此orf区两端设计全长序列的上、下游引物,经rt-pcr扩增、克隆验证后得到1条长度为1071bp的片段,与预期片段大小一致。起始密码子为atg,终止密码子为tga,编码356个氨基酸。
2.2tauam3基因的生物信息学分析
采用protparam工具对其编码蛋白分析发现,tauam3蛋白相对分子量为40.56kda,等电点6.01,分子式为c1850h2830n466o516s22,不稳定指数为26.89,表明该蛋白是稳定性蛋白质。该蛋白亲水性平均数为-0.212,与利用protscale在线分析的疏水性结果一致。通过signalp-5.0server、tmhmmserverv.2.0和netphos3.1server对tauam3氨基酸序列进行在线分析,结果表明tauam3无信号肽和跨膜结构,含有12个丝氨酸(ser)磷酸化位点、7个苏氨酸(thr)磷酸化位点和6个酪氨酸(tyr)磷酸化位点。二级结构预测发现tauam3由无规则卷曲、α-螺旋、延伸链结构和β-转角组成。其中无规则卷曲居多,占41.3%(约147个氨基酸),其次是α-螺旋占39.0%(约139个氨基酸),延伸链结构占16.0%(约57个氨基酸),β-转角占3.7%(约13个氨基酸)。经过对氨基酸序列进行保守结构域预测,结果显示该蛋白在氨基酸序列的1-340位有一个高度保守的rgp超家族结构域,表明其属于rgp家族成员。
2.3tauam3的染色体定位与dna全长克隆
利用graingenes数据库对tauam3进行预测分析,初步将该基因定位于小麦3bl染色体上。利用tauam3的orf全长引物,以小麦基因组dna为模板进行pcr扩增,得到1条有效扩增产物。经克隆、测序获得1290bp的dna序列。将此序列与该基因cdna序列进行比对,显示该基因含有2个内含子,分别长131和88bp。
2.4同源性及进化树分析
通过dnaman分析tauam3与其他植物udp-ara变位酶氨基酸序列的同源性。结果表明,tauam3与大麦hvuam3、短柄草bdrgp3以及高粱同源蛋白具有很高的同源性,同源性分别达到95.1%、87.3%和85.9%,与水稻osuam1和玉米zmuam3同源性均为83.2%,与拟南芥atrgp1和蓖麻rcrgp4的同源性分别为82.1%与57.5%。进一步采用mega7.0软件构建系统发育树发现,8个udp-ara变位酶蛋白可以被划分为两个分支,atrgp1和rcrgp4聚在同一个分支上,而hvuam3、bdrgp3、tauam3、sbuam、osuam1和zmuam3聚在另一分支上(图1),其中tauam3和hvuam3的亲缘关系最近,两者应该具有相似的功能。
2.5tauam3蛋白的亚细胞定位分析
对tauam3蛋白进行亚细胞定位预测结果显示,该蛋白定位在细胞质、线粒体、细胞核、质膜和液泡上的概率分别为60.9%、21.7%、8.7%、4.3%和4.3%,结果表明定位在细胞质的概率最大,说明该蛋白很可能存在于细胞质中。
2.6tauam3的组织特异性表达
利用实时定量pcr技术检测tauam3在幼苗期小麦根、子叶、第二叶及成株期小麦茎、旗叶和穗中的表达量,结果表明该基因在所有检测的组织中都能被检测到。tauam3在茎部表达水平较低,在穗和旗叶中表达水平较高,分别为茎中表达量的5.8倍和6.1倍,在根和第二叶中高丰度表达,分别为茎中该基因表达量的8.1和8.0倍(图2)。该结果表明,tauam3基因在小麦中的表达丰度存在组织特异性。
2.7胁迫处理后小麦tauam3的表达
tauam3在sa处理后2h即被检测到上调表达,在6h时表达量达到峰值,此时表达量为未处理对照的7.8倍,之后tauam3的表达量开始降低,直至24h,其表达量恢复到对照水平。tauam3在meja处理后表现出强烈的上调表达,处理后2htauam3的表达量开始上升,其表达量在处理后6h达到未接种对照的8.2倍,直至24h一直持续在较高水平。tauam3在aba和eth处理后表达模式比较一致,表达量均在处理12h后达到峰值,分别为未处理对照的4.1倍和5.8倍(图3)。该结果表明,sa、meja、aba和eth处理均能诱导tauam3的表达,但是诱导表达的程度有所不同。
环境胁迫处理结果(图4)表明,nacl处理后tauam3基因表达呈上升趋势,至24h达到未处理对照的3.7倍。peg模拟干旱胁迫处理后tauam3基因的表达量变化不明显,仅在处理后12h达到对照的1.6倍。该研究结果表明,高盐胁迫能诱导tauam3基因的上调表达。
以上所述仅为本发明的较佳实施例,并不用于限制本发明,凡在本发明的技术启示和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
序列表
<110>西北农林科技大学
<120>小麦udp-ara变位酶及编码基因与用途
<130>2100371
<141>2021-06-04
<160>2
<170>siposequencelisting1.0
<210>1
<211>356
<212>prt
<213>小麦变位酶(udp-araudp-ara)
<400>1
metalaproleuleulysaspgluleuaspilevalileprothrile
151015
argasnleuasppheleuglumettrpargprophepheglnprotyr
202530
hisleuileilevalglnaspglyaspprothrargthrvalmetval
354045
progluglypheasptyrgluleutyrasnargasnaspileasnarg
505560
ileleuglyprolysalasercysileserphelysaspseralacys
65707580
argcyspheglytyrmetvalserlyslyslystyrvaltyrthrile
859095
aspaspaspcysphevalalalysaspprothrglylysaspileasp
100105110
alaleualalyshisileglnasnleuleucysproserthrproleu
115120125
phepheasnthrleutyraspprotyrglngluglyalaasppheval
130135140
argglytyrpropheserleuarggluglyvalprothralavalser
145150155160
hisglyleutrpleuasnileproasptyraspalaprothrglnleu
165170175
vallysproarggluargasnglyargtyrvalaspalavalmetthr
180185190
ileprolysglyserleupheprometcysglymetasnleualaphe
195200205
asparggluleuileglyproalamettyrpheglyleumetglyasp
210215220
glyglnproileglyargtyraspaspmettrpalaglytrpcysval
225230235240
lysvalilecysasphisleuglyleuglyvallysthrglyleupro
245250255
tyriletrphisserlysalaserasnprophevalasnleulyslys
260265270
glutyrlysglyilephetrpglngluaspileileprophephegln
275280285
alaalalysleuthrlysglucysaspthrvalglnlyscystyrile
290295300
serleuserglnglnvallysglulysleuglylysileaspprotyr
305310315320
phevallysleualaaspalametvalthrtrpileglualatrpasp
325330335
metleuasnserlysaspserlysglualaaspvalalaasnglylys
340345350
leulysglylys
355
<210>2
<211>1071
<212>dna
<213>小麦udp-ara变位酶编码基因tauam3()
<400>2
atggcgccgctgctcaaggacgagctggacatcgtcatcccgacgatccggaacctggac60
ttcctcgagatgtggcggcccttcttccagccgtaccacctcatcatcgtgcaggacggc120
gaccccacgaggaccgtcatggtgcccgagggcttcgactacgagctctacaaccgcaac180
gacatcaaccgcatcctcggccccaaggcctcctgcatctccttcaaggactccgcctgc240
cgctgcttcggctacatggtctccaagaagaagtacgtctacaccatcgacgacgattgc300
ttcgtggccaaggaccccacggggaaggacatcgacgcgctggccaagcacatccagaac360
ctgctctgcccgtccacgccgctcttcttcaacaccttgtacgacccctaccaggagggc420
gccgacttcgtgcgcggctacccgttcagcctccgggagggcgtgcccaccgccgtgtcg480
catggcctctggctcaacatccccgactatgacgcgcccacgcagctggtcaagccgcgg540
gagcgcaacggcaggtacgtcgacgccgtcatgaccatccccaagggctccctcttcccc600
atgtgcggcatgaacctcgccttcgaccgcgagctcatcggcccggccatgtacttcggc660
ctcatgggcgacggccagcccatcggacgctacgacgacatgtgggccggctggtgcgtc720
aaggtgatctgcgaccaccttgggctgggagtcaagacggggctgccctacatctggcac780
agcaaggccagcaacccgttcgtgaacctgaagaaggagtacaagggcatcttctggcag840
gaggacatcatccccttctttcaggccgcgaagctgaccaaggaatgcgacaccgtgcag900
aagtgctacatctcgctctcgcagcaggtcaaggagaagctcggcaagatcgacccctac960
ttcgtcaagctcgccgacgccatggtcacctggatcgaggcctgggacatgctcaactcc1020
aaggactccaaggaggccgacgtcgccaacggcaaactcaaggggaagtag1071
- 还没有人留言评论。精彩留言会获得点赞!