L-赖氨酸生产能力得到提高的谷氨酸棒状杆菌突变株及利用其的L-赖氨酸的生产方法与流程
l-赖氨酸生产能力得到提高的谷氨酸棒状杆菌突变株及利用其的l-赖氨酸的生产方法
技术领域
1.本发明涉及l-赖氨酸生产能力得到提高的谷氨酸棒状杆菌突变株及利用其的l-赖氨酸的生产方法。
背景技术:
2.l-赖氨酸是人体或动物体内不能合成的必要氨基酸,必须从外部供给,一般情况下,是通过利用了细菌或酵母之类的微生物的发酵而生产的。l-赖氨酸的生产可以利用自然状态下获得的野生型菌株或者以提高其l-赖氨酸生产能力的方式转变的突变株。近年来,为了改善l-赖氨酸的生产效率,以多用于l-氨基酸和其它有用物质的生产的大肠杆菌、棒状杆菌等微生物为对象,适用基因重组技术,从而开发了具有优异的l-赖氨酸生产能力的各种重组菌株或突变株及利用其的l-赖氨酸生产方法。
3.根据韩国授权专利第10-0838038号和第10-2139806号,通过变更对包含与l-赖氨酸生产有关的酶的蛋白质进行编码的基因的碱基序列或氨基酸序列来增加该基因的表达或者剔除不必要的基因,从而可以提高l-赖氨酸生产能力。此外,在韩国公开专利第10-2020-0026881号中公开了为了增加对参与l-赖氨酸生产的酶进行编码的基因的表达而将基因的现有启动子变更为具有强活性的启动子的方法。
4.如上所述,开发了增加l-赖氨酸生产能力的各种方法,但与l-赖氨酸生产直接或间接相关的酶、转录因子、运输蛋白质等蛋白质的种类达到数十余种,因此关于根据这样的蛋白质的活性变化的l-赖氨酸生产能力是否增加,事实上仍需要大量的研究。
5.现有技术文献
6.专利文献
7.韩国授权专利第10-0838038号
8.韩国授权专利第10-2139806号
9.韩国公开专利第10-2020-0026881号
技术实现要素:
10.本发明的目的在于提供l-赖氨酸生产能力得到提高的谷氨酸棒状杆菌(corynebacterium glutamicum)突变株。
11.另外,本发明的目的在于提供利用了上述突变株的l-赖氨酸的生产方法。
12.本发明的发明人为了利用谷氨酸棒状杆菌菌株来开发l-赖氨酸生产能力得到提高的新型突变株而进行了研究,结果确认了在将编码作用于l-赖氨酸生物合成途径的最后步骤的二氨基庚二酸脱羧酶的lysa基因的启动子中的特定位置的碱基序列进行置换时,l-赖氨酸生产量增加,从而完成了本发明。
13.本发明的一个方式提供二氨基庚二酸脱羧酶的活性增强而l-赖氨酸生产能力得到提高的谷氨酸棒状杆菌突变株。
14.本发明中所使用的“二氨基庚二酸脱羧酶(diaminopimelate decarboxylase)”是指在l-赖氨酸生物合成途径的最后步骤中分解内消旋-二氨基庚二酸(meso-diaminoheptanedioate,mdap)的碳键而对生成二氧化碳和l-赖氨酸的反应进行催化的酶。
15.根据本发明的一具体例,上述二氨基庚二酸脱羧酶可以源于棒状杆菌(corynebacterium)属菌株。具体而言,上述棒状杆菌属菌株可以为谷氨酸棒状杆菌(corynebacterium glutamicum)、克氏棒状杆菌(corynebacterium crudilactis)、荒漠棒状杆菌(corynebacterium deserti)、帚石南棒状杆菌(corynebacterium callunae)、苏那棒状杆菌(corynebacterium suranareeae)、润滑脂棒状杆菌(corynebacterium lubricantis)、道桑棒状杆菌(corynebacterium doosanense)、高效棒状杆菌(corynebacterium efficiens)、乌氏棒状杆菌(corynebacterium uterequi)、停滞棒状杆菌(corynebacterium stationis)、帕氏棒状杆菌(corynebacterium pacaense)、奇棒状杆菌(corynebacterium singulare)、腐殖质还原棒状杆菌(corynebacterium humireducens)、海洋棒状杆菌(corynebacterium marinum)、耐盐棒状杆菌(corynebacterium halotolerans)、蝶科棒状杆菌(corynebacterium spheniscorum)、弗氏棒状杆菌(corynebacterium freiburgense)、纹带棒状杆菌(corynebacterium striatum)、犬棒状杆菌(corynebacterium canis)、产氨棒状杆菌(corynebacterium ammoniagenes)、肾棒状杆菌(corynebacterium renale)、污染棒状杆菌(corynebacterium pollutisoli)、亚胺棒状杆菌(corynebacterium imitans)、里海棒状杆菌(corynebacterium caspium)、睾丸棒状杆菌(corynebacterium testudinoris)、假性棒状杆菌(corynebacaterium pseudopelargi)或者微黄棒状杆菌(corynebacterium flavescens),但并不限定于此。
16.本发明中所使用的“活性增强”是指新导入或增大对作为目标的酶、转录因子、运输蛋白质等蛋白质进行编码的基因的表达,从而与野生型菌株或转变前的菌株相比,表达量增加。这样的活性的增强包括以下情况:通过编码基因的核苷酸置换、插入、缺失或者它们的组合,蛋白质本身的活性与原始微生物具有的蛋白质的活性相比增加的情况,以及通过编码它的基因的表达增加或者翻译增加等,细胞内的整体酶活性程度与野生型菌株或转变前的菌株相比高的情况,也包括它们的组合。
17.根据本发明的一具体例,上述二氨基庚二酸脱羧酶的活性增强可以是对编码二氨基庚二酸脱羧酶的基因的启动子诱导位置特异性突变。
18.根据本发明的一具体例,上述编码二氨基庚二酸脱羧酶的基因的启动子可以由seq id no:1的碱基序列表示。
19.本发明中所使用的“启动子”是指调节基因转录的dna的特定位点,包括起始目标基因的mrna转录的rna聚合酶(polymerase)的结合位点,通常位于以转录起始点为基准的上游(upstream)。原核生物中的启动子定义为rna聚合酶结合的转录起始点周围的位点,通常由从转录起始点向前的-10区域和-35区域的碱基对分开的两个短的碱基序列构成。本发明中的启动子突变是被改良为与野生型启动子相比具有高活性,通过在位于转录起始点的上游的启动子区域内诱导突变,从而可以增加位于下游(downstream)的基因的表达。
20.根据本发明的一具体例,上述二氨基庚二酸脱羧酶的活性增强可以是从编码二氨基庚二酸脱羧酶的基因的启动子序列内的转录起始点向前的-25至-10区域中的一个以上
的碱基被置换。
21.更具体而言,本发明中的启动子突变可以是-25至-10区域中的1个以上的碱基,优选为-20至-15区域、-19至-16区域、或者-18和-17区域中的1个、2个、3个、4个、或者5个碱基被连续或非连续地置换。
22.根据本发明的一个实施例,编码二氨基庚二酸脱羧酶的lysa基因构成了编码精氨酰-trna合成酶(arginyl-trna synthetase)的args基因和操纵子(operon),args-lysa操纵子通过一个启动子来调节。因此,在谷氨酸棒状杆菌菌株的args-lysa操纵子的启动子序列中,将-17和-18区域的碱基序列由cg置换为ct,从而获得了具有lysa基因的新型启动子序列的谷氨酸棒状杆菌突变株。这样的谷氨酸棒状杆菌突变株可以包含由seq id no:2的碱基序列表示的lysa基因的突变的启动子序列。
23.另外,根据本发明的一个实施例,在谷氨酸棒状杆菌菌株的args-lysa操纵子的启动子序列中,将-17和-18区域的碱基序列由cg置换为ga,从而获得了具有lysa基因的新型启动子序列的谷氨酸棒状杆菌突变株。这样的谷氨酸棒状杆菌突变株可以包含由seq id no:3的碱基序列表示的lysa基因的突变的启动子序列。
24.根据本发明的一个实施例,在谷氨酸棒状杆菌菌株的args-lysa操纵子的启动子序列中,将-17和-18区域的碱基序列由cg置换为gt,从而获得了具有lysa基因的新型启动子序列的谷氨酸棒状杆菌突变株。这样的谷氨酸棒状杆菌突变株可以包含由seq id no:4的碱基序列表示的lysa基因的突变的启动子序列。
25.本发明中所使用的“生产能力得到提高”是指与亲本菌株相比,l-赖氨酸的生产率增加。上述亲本菌株是指成为突变对象的野生型或突变株,包括直接成为突变的对象或通过重组的载体等转化的对象。在本发明中,亲本菌株可以是野生型谷氨酸棒状杆菌菌株或由野生型突变的菌株。
26.根据本发明的一具体例,上述亲本菌株是在参与赖氨酸生产的基因(例如,lysc、zwf和hom基因)的序列上诱导了突变的突变株,可以是在韩国微生物保藏中心(korean culture center of microorganisms)于2021年4月2日以保藏号kccm12969p保藏的谷氨酸棒状杆菌菌株(以下称为“谷氨酸棒状杆菌ds1菌株”)。
27.根据本发明的一个实施例,上述l-赖氨酸生产能力得到提高的谷氨酸棒状杆菌突变株通过包含编码二氨基庚二酸脱羧酶的lysa基因的启动子突变,从而与亲本菌株相比,显示出增加的l-赖氨酸生产能力,特别是,与亲本菌株相比,l-赖氨酸生产量增加2%以上,具体而言,增加2至40%,更具体而言,增加3至30%,从而每1l的菌株培养液可以生产60~80g的l-赖氨酸,优选地,可以生产65~75g的l-赖氨酸。
28.根据本发明的一具体例的谷氨酸棒状杆菌突变株可以通过包含亲本菌株中的编码二氨基庚二酸脱羧酶的基因的启动子序列被部分置换的突变体的重组载体而实现。
29.本发明中所使用的“部分”是指不是碱基序列或多核苷酸序列的全部,可以为1至300个,优选可以为1至100个,更优选可以为1至50个,但并不限定于此。
30.本发明中所使用的“突变体”是指参与l-赖氨酸的生物合成的二氨基庚二酸脱羧酶基因的启动子序列内的-25至-10区域中的一个以上的碱基被置换的启动子突变体。
31.根据本发明的一具体例,上述二氨基庚二酸脱羧酶基因的启动子序列内的-17和-18区域的碱基序列被置换为ct的突变体可以具有seq id no:2的碱基序列,被置换为ga的
突变体可以具有seq id no:3的碱基序列,被置换为gt的突变体可以具有seq id no:4的碱基序列。
32.本发明中所使用的“载体”作为可以在合适的宿主细胞中表达目标蛋白质的表达载体,是指为了表达基因插入物而包括可操作地连接的(operably linked)必需的调控元件的基因产物。在这里,“可操作地连接的”是指需要表达的基因与其调节序列彼此功能性地结合并以能够进行基因表达的方式连接,“调控元件”包括用于实施转录的启动子、用于调节转录的任意的操纵子序列、编码合适的mrna核糖体结合位点的序列、以及调节转录和翻译的终止的序列。这样的载体包括质粒载体、粘粒载体、噬菌体载体、病毒载体等,但并不限定于此。
33.本发明中所使用的“重组载体”在转化到合适的宿主细胞内后,可以与宿主细胞的基因组无关地进行复制,或者可以缝合于基因组其自身。这时,上述“合适的宿主细胞”能够进行载体的复制,可以包括作为开始复制的特定碱基序列的复制起点。
34.在上述转化中,根据宿主细胞而选择合适的载体导入技术,从而可以在宿主细胞内表达作为目标的基因。例如,载体导入可以通过电穿孔法(electroporation)、热冲击(heat-shock)、磷酸钙(capo4)沉淀、氯化钙(cacl2)沉淀、显微注射法(microinjection)、聚乙二醇(peg)法、deae-葡聚糖法、阳离子脂质体法、乙酸锂-dmso法、或者它们的组合而实施。转化的基因只要可以在宿主细胞内表达,就可以被包括在内,而不限制于将其插入宿主细胞的染色体内或者位于染色体外。
35.上述宿主细胞包括在生物体内或试管内用本发明的重组载体或多核苷酸转染、转化、或者感染的细胞。包含本发明的重组载体的宿主细胞为重组宿主细胞、重组细胞或重组微生物。
36.另外,根据本发明的重组载体可以包括选择标记(selection marker),上述选择标记用于选择利用载体转化的转化体(宿主细胞),在经上述选择标记处理的培养基中只有表达选择标记的细胞可以生存,因此能够选择转化的细胞。作为代表性的例子,上述选择标记有卡那霉素、链霉素、氯霉素等,但并不限定于此。
37.插入到本发明的转化用重组载体内的基因由于同源重组交叉而可以被置换到如棒状杆菌属微生物等的宿主细胞内。
38.根据本发明的一具体例,上述宿主细胞可以为棒状杆菌属菌株,例如,可以为谷氨酸棒状杆菌ds1菌株。
39.另外,本发明的另一方式提供l-赖氨酸的生产方法,其包括以下步骤:a)在培养基中培养上述谷氨酸棒状杆菌突变株的步骤;以及b)从上述突变株或者培养突变株的培养基中回收l-赖氨酸的步骤。
40.上述培养可以根据该领域中已知的合适的培养基和培养条件而进行,只要是本领域技术人员就可以容易地调整培养基和培养条件来使用。具体而言,上述培养基可以是液体培养基,但并不限定于此。培养方法可以包括例如分批式培养(batch culture)、连续式培养(continuous culture)、补料分批式培养(fed-batch culture)或者它们的组合培养,但并不限定于此。
41.根据本发明的一具体例,上述培养基必须以合适的方式满足特定菌株的要求,可以由本领域技术人员适当地进行变形。关于棒状杆菌属菌株的培养基,可以参考公知的文
献(manual of methods for general bacteriology.american society for bacteriology.washington d.c.,usa,1981),但并不限定于此。
42.根据本发明的一具体例,培养基中可以包含各种碳源、氮源和微量元素成分。作为可以使用的碳源,包括葡萄糖、蔗糖、乳糖、果糖、麦芽糖、淀粉、纤维素之类的糖及碳水化合物;大豆油、向日葵油、蓖麻籽油、椰子油等油及脂肪;棕榈酸、硬脂酸、亚油酸之类的脂肪酸;甘油、乙醇之类的醇;乙酸之类的有机酸。这些物质可以单独使用或以混合物的形式使用,但并不限定于此。作为可以使用的氮源,可以包括蛋白胨、酵母提取物、肉汤、麦芽提取物、玉米浆、豆粕和尿素或者无机化合物,例如,硫酸铵、氯化铵、磷酸铵、碳酸铵和硝酸铵。氮源同样可以单独使用或以混合物的形式使用,但并不限定于此。作为可以使用的磷的供应源,可以包括磷酸二氢钾或磷酸氢二钾或相应的含钠的盐,但并不限定于此。此外,培养基可以含有生长所需要的硫酸镁或硫酸铁之类的金属盐,但并不限定于此。除此以外,可以包含氨基酸和维生素之类的必要生长物质。此外,可以使用合适培养基的前体。上述培养基或单个成分可以在培养过程中通过适当的方式,分批或连续添加到培养液中,但并不限定于此。
43.根据本发明的一具体例,在培养过程中,可以将氢氧化铵、氢氧化钾、氨、磷酸和硫酸之类的化合物以适当的方式添加到微生物培养液中来调整培养液的ph。此外,在培养过程中,可以使用脂肪酸聚乙二醇酯之类的消泡剂来抑制气泡产生。进一步而言,为了维持培养液的有氧状态,可以向培养液内注入氧气或者含氧气的气体(例如空气)。培养液的温度通常可以为20℃至45℃,例如,可以为25℃至40℃。培养时间可以持续到有用物质获得期望的生产量为止,例如,可以为10至160小时。
44.根据本发明的一具体例,在从上述培养的突变株和培养突变株的培养基中回收l-赖氨酸的步骤中,可以根据培养方法并利用该领域中公知的合适的方法,从培养基中收集或回收所生产的l-赖氨酸。例如,可以使用离心分离、过滤、提取、喷雾、干燥、蒸发、沉淀、结晶化、电泳、分级溶解(例如,硫酸铵沉淀)、色谱法(例如,离子交换、亲和性、疏水性和尺寸排阻)等方法,但并不限定于此。
45.根据本发明的一具体例,在回收赖氨酸的步骤中,可以将培养基低速离心分离而去除生物质,可以将得到的上清液通过离子交换色谱法而分离。
46.根据本发明的一具体例,在上述回收l-赖氨酸的步骤中,可以包括纯化l-赖氨酸的工序。
47.根据本发明的谷氨酸棒状杆菌突变株通过使编码二氨基庚二酸脱羧酶的基因的表达增加或增强,从而与亲本菌株相比,可以提高l-赖氨酸的生产收率。
附图说明
48.图1表示根据本发明的一实施例的包含启动子-17和-18位点由cg置换为ct的args-lysa操纵子的启动子的pcgi(pm1-args+lysa)载体的结构。
49.图2表示根据本发明的一实施例的包含启动子-17和-18位点由cg置换为ga的args-lysa操纵子的启动子的pcgi(pm2-args+lysa)载体的结构。
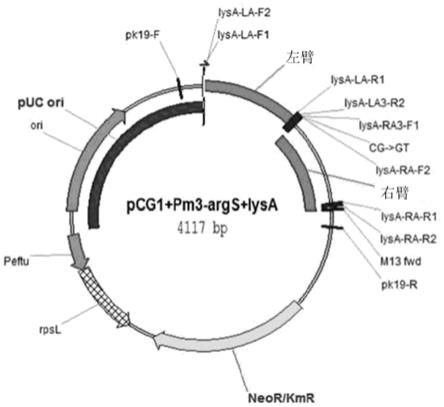
50.图3表示根据本发明的一实施例的包含启动子-17和-18位点由cg置换为gt的args-lysa操纵子的启动子的pcgi(pm3-args+lysa)载体的结构。
具体实施方式
51.下面,对本发明更详细地进行说明。但是,这样的说明只是为了帮助理解本发明而例示性地提出的,本发明的范围并不限定于这样的例示性的说明。
52.实施例1.谷氨酸棒状杆菌突变株的制造
53.为了制造二氨基庚二酸脱羧酶的活性增强的谷氨酸棒状杆菌突变株,使用了谷氨酸棒状杆菌ds1菌株和e.coli dh5a(hit competent cells
tm
,cat no.rh618)。
54.上述谷氨酸棒状杆菌ds1菌株在cm-肉汤(cm-broth)培养基(ph 6.8)中以30℃的温度进行了培养,上述cm-肉汤培养基的组成是在1l的蒸馏水中有5g的葡萄糖、2.5g的nacl、5.0g的酵母提取物、1.0g的尿素(urea)、10.0g的聚蛋白胨和5.0g的牛肉(beef)提取物。
55.上述e.coli dh5a在lb培养基上以37℃的温度进行了培养,上述lb培养基的组成是在1l的蒸馏水中有10.0g的胰蛋白胨、10.0g的nacl和5.0g的酵母提取物。
56.抗生素卡那霉素(kanamycin)和链霉素(streptomycin)使用了sigma公司的制品。tdna测序分析委托马克罗基因株式会社进行了分析。
57.1-1.重组载体的制作
58.为了增加赖氨酸生产率,在菌株中导入作用于赖氨酸生物合成途径的最后步骤的二氨基庚二酸脱羧酶的增强。在本实施例中利用的方法为了增加编码二氨基庚二酸脱羧酶的lysa基因的表达,对args-lysa操纵子的启动子诱导了特异性突变。将args-lysa操纵子的启动子的-17和-18位点的碱基序列由cg置换为ct,在谷氨酸棒状杆菌基因组上,以args-lysa操纵子的正中间为中心,用pcr扩增左臂510bp部分和右臂480bp部分,通过重叠pcr(overlap pcr)方法连接后,克隆到作为重组载体的pcgi(参考文献[kim et al.,journal of microbiological methods 84(2011)128-130])中。将上述质粒命名为pcgi(pm1-args+lysa)(参考图1)。为了制作上述质粒,使用了下述表1的引物扩增各自的基因片段。
[0059]
【表1】
[0060]
[0061]
利用以上的引物并在以下的条件下实施pcr。利用热循环仪(thermocycler)(tp600,takara bio inc.,日本),在添加有各个三磷酸脱氧核苷酸(datp、dctp、dgtp、dttp)100μm的反应液中,将1pm的寡核苷酸、10ng的谷氨酸棒状杆菌ds1菌株的染色体dna,利用模板(template),在1单位的pfu-x dna聚合酶混合物(solgent)的存在下,实施了25~30周期(cycle)。pcr实施条件是在(i)变性(denaturation)步骤:在94℃30秒,(ii)退火(annealing)步骤:在58℃30秒,以及(iii)延伸(extension)步骤:在72℃1~2分钟(每1kb赋予2分钟的聚合时间)的条件下实施的。
[0062]
利用自组装克隆(self-assembly cloning),将如上所述制造的基因片段克隆到pcgi载体中。将上述载体转化到e.coli dh5a中,涂抹在含有50μg/ml的卡那霉素的lb-琼脂板上,在37℃培养24小时。分离最终形成的菌落,确认插入物(insert)是否正确存在于载体后,分离该载体,用于谷氨酸棒状杆菌菌株的重组。
[0063]
作为上述方法中共同进行的过程,该基因的扩增是由谷氨酸棒状杆菌ds1菌株基因组dna用pcr方法进行扩增的,根据策略,通过自组装克隆(self-assembled cloning)方法插入到e.coli dh5a中选择。染色体的基因碱基置换(chromosomal base substitution)是分别扩增各个片段的基因,用层叠pcr制造了目标dna片段。在基因操作时,作为pcr扩增酶,利用了ex taq聚合酶(takara)、pfu聚合酶(solgent),各种限制酶和dna修饰酶使用了neb制品,根据供给的缓冲液和方案进行了使用。
[0064]
1-2.突变株的制造
[0065]
利用上述pcgi(pm1-args+lysa)载体制造了作为突变菌株的ds3菌株。以上述载体的最终浓度成为1μg/μl以上的方式准备,对谷氨酸棒状杆菌ds1菌株使用电穿孔法(参考文献[tauch et al.,fems microbiology letters 123(1994)343-347])而诱导了1次重组。这时,将电穿孔的菌株涂抹在含有20μg/μl的卡那霉素的cm-琼脂平板上,分离菌落后,通过pcr和碱基序列分析确认了是否正确插入到了基因组的诱导位置。为了将这样分离的菌株再次诱导2次重组,接种在含有链霉素的cm-琼脂液体培养基中,培养一夜以上后,涂抹在含有相同浓度的链霉素的琼脂培养基中,分离菌落。在最终分离的菌落中确认了是否存在对卡那霉素的耐性后,通过碱基序列分析确认了在没有抗生素耐性的菌株中是否向lysa基因导入了突变(参考文献[schafer et al.,gene 145(1994)69-73])。最终,获得了导入有突变lysa基因的谷氨酸棒状杆菌突变株(ds3)。
[0066]
实施例2.谷氨酸棒状杆菌突变株的制造
[0067]
将args-lysa操纵子的启动子的-17和-18位点的碱基序列由cg置换为ga,除此以外,通过与实施例1相同的方法制造了谷氨酸棒状杆菌突变株。
[0068]
在这里,为了制作质粒,使用了下述表2的引物扩增各个基因片段,利用制作的质粒pcgi(pm2-args+lysa)载体(参照图2)而制造了作为突变菌株的ds3-1菌株。最终,获得了导入有突变lysa基因的谷氨酸棒状杆菌突变株(ds3-1)。
[0069]
【表2】
[0070][0071]
实施例3.谷氨酸棒状杆菌突变株的制造
[0072]
将args-lysa操纵子的启动子的-17和-18位点的碱基序列由cg置换为gt,除此以外,通过与实施例1相同的方法制造了谷氨酸棒状杆菌突变株。
[0073]
在这里,为了制作质粒,使用了下述表3的引物扩增各个基因片段,利用制作的质粒pcgi(pm3-args+lysa)载体而制造了作为突变菌株的ds3-2菌株。最终,获得了导入有突变lysa基因的谷氨酸棒状杆菌突变株(ds3-2)。
[0074]
【表3】
[0075][0076]
实验例1.突变株的l-赖氨酸生产率比较
[0077]
将亲本菌株谷氨酸棒状杆菌ds1菌株和实施例1至3中制造的作为生产赖氨酸的突
变株的ds2菌株、ds2-1菌株和ds2-2菌株的l-赖氨酸的生产率进行了比较。
[0078]
在含有10ml的具有下述表4所述的组成的赖氨酸培养基的100ml的烧瓶中,分别接种菌株,在30℃以180rpm的条件震荡培养48小时。培养结束后,赖氨酸分析用hplc(岛津,日本)测定了l-赖氨酸的生产量,将其结果示于下述表5。
[0079]
【表4】
[0080]
组成含量(1l蒸馏水基准)葡萄糖(glucose)100g硫酸铵(ammonium sulfate)55gkh2po41.1gmgso4·
h2o1.2gmnso4·
h2o180mgfeso4·
h2o180mg硫胺素(thiamine)
·
hcl9mg生物素(biotin)1.8mgcaco35%ph7.0
[0081]
【表5】
[0082]
菌株od610l-赖氨酸(g/l)每单位菌体的l-赖氨酸生产量(g/gdcw)亲本菌株(ds1)24.064.86.4突变株(ds3)23.065.26.7突变株(ds3-1)23.065.86.8突变株(ds3-2)22.066.97.2
[0083]
如上述表5所示的那样,确认了在谷氨酸棒状杆菌突变株ds3、ds3-1和ds3-2中,为了增强赖氨酸生物合成途径,args-lysa操纵子的启动子序列的特定位置(-17和-18区域)被置换成最佳的碱基序列(ct、ga或gt),与亲本菌株谷氨酸棒状杆菌ds1菌株相比,l-赖氨酸的生产率分别增加约4.7%、6.3%、12.5%。通过这样的结果,可知通过lysa基因的表达增强促进赖氨酸前体的碳键分解,从而提高了菌株的l-赖氨酸生产能力。
[0084]
到目前为止,对本发明围绕其优选实施例进行了研究。本发明所属技术领域的技术人员可以理解本发明在不脱离本发明的本质性的特性的范围内可以以变形的形态实现。因此,公开的实施例应从说明性的角度进行考虑而不是从限制性的角度进行考虑。本发明的范围显示在权利要求书中而不是上述的说明中,并且应被解释为在与其等同范围内的所有的差异包括在本发明中。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1