本氏烟分泌型腺毛调控基因NbJAZ3、其表达载体及应用
本氏烟分泌型腺毛调控基因nbjaz3、其表达载体及应用
技术领域
1.本发明涉及分子生物学技术领域,具体涉及一种本氏烟分泌型腺毛调控基因nbjaz3、其表达载体及应用。
背景技术:
2.表皮毛是植物表面的一种特化结构。根据表皮毛是否有无分支,可分为分支表皮毛和非分支表皮毛;根据腺体是否有无,可分为腺体表皮毛和非腺体表皮毛;根据无分泌能力存在与否,可分为分泌型和非分泌型;根据表皮毛的细胞数量,分为单细胞和多细胞表皮毛。植物表皮毛具有极其重要的作用,植物在其生长和发育过程中会受到许多不同类型的生物和非生物胁迫,植物表皮毛位于植物的表皮层和外部环境之间,增加植物表皮厚度,不仅为植物提供了天然的物理屏障,它还有效地减轻了有害生物的损害以及冰冻和紫外线对植物自身造成的损害和机械损伤,维持植物细胞离子稳态,减少植物的水分流失和热量损耗;另外,一些表皮毛可以分泌多种次生代谢物质,并与表皮上的气孔、角质和蜡质等共同发挥保护作用。例如,表皮毛还可以通过分泌驱虫剂、生物碱或有毒物质来防止生物损害,保护植物免受食草动物、昆虫和病原菌感染。
3.拟南芥作为常见的模式植物,其表皮毛通常是由一个细胞组成,是典型的单细胞结构。单细胞表皮毛的生长发育主要受wd40
‑
bhlh
‑
myb复合体控制。然而,大多数陆生植物产生的是多细胞表皮毛,例如,烟草、棉花、番茄、金鱼草和矮牵牛等。研究表明,多细胞表皮毛生长发育的分子调控机制与单细胞表皮毛不相同。
4.茉莉酸(jasmonic acid,ja)参与调控植物营养生长过程,组织分化发育,植株衰老和多种防御反应。其中,外源茉莉酸作为腺毛分化与发育过程中重要的调控因子。jaz(jasmonate zim
‑
domain)是一种植物特有的转录因子,在ja信号中作为重要的转录抑制子。jaz蛋白包含一个tify家族特有的tify保守结构域:tif[f/y]xg,其c端还有一个保守的jas结构域:s
‑
l
‑
x(2)
‑
f
‑
x(2)
‑
k
‑
r
‑
x(2)
‑
r。大量研究表明,在植物正常生长时,jaz蛋白与下游转录因子bhlhs,mybs,hds等结合,抑制其转录活性。当jaz蛋白被scf
coi1
复合体介导的泛素/26s蛋白酶体途径降解后,下游相关基因被释放,促进植株生长发育等过程。例如,jaz蛋白能够与myb21和myb24互作抑制它们的转录活性从而调控雄蕊的发育;jaz蛋白能与tt8、gl3、egl3、myb75和gl1互作并削弱它们的转录活性,从而抑制表皮毛的发生和花青素的积累;jaz4、jaz8通过与wrky57转录因子互作调控ja诱导的叶片衰老;jaz蛋白能与toe1和toe2互作从而调控植物开花时间;jaz通过与hd转录因子互作调控ja诱导的腺毛发生。
[0005]
jaz蛋白调控拟南芥单细胞表皮毛发育的分子机制较清晰,其具体调控模式为,当植物体内的ja处于低水平时,jaz蛋白能与myb类的转录因子gl1/myb75、bhlh类的转录因子gl3/egl3结合,从而抑制myb
‑
bhlh
‑
ttg1复合体的活性,使依赖于ja的表皮毛发生受到抑制;但是当植物体内ja水平升高时,会促使coi1受体与jaz蛋白结合并使jaz蛋白被scf
coi1
复合体泛素化,进而被26s蛋白酶体所降解,使原本受jaz蛋白抑制的myb
‑
bhlh
‑
ttg1复合体被释放出来并恢复其转录活性,激活下游参与表皮毛发育调控的基因的表达,最终促进拟
南芥单细胞表皮毛的发生;另外,棉花ghjaz2通过抑制ghmyb25
‑
like、ghgl1、ghmyc2、ghwd40和ghji1负调控棉花单细胞表皮毛的发育,这与拟南芥表皮毛发育的调控机制相似。
[0006]
烟草腺毛分泌物是烟叶香气成分的重要前体物,而烟草腺毛的类型和结构、腺毛发育、腺毛的密度变化等都会影响到分泌物的产生和组成,因此烟草腺毛基因调控研究在烟草栽培学研究上具有重要的现实意义。但当前关于jaz蛋白调控多细胞腺毛发育的研究较少(如青蒿aajaz与一个新的转录因子aahd1共同参与调控青蒿多细胞腺毛的发育;番茄jaz2过量表达后参与调控表皮毛发生的关键基因的表达量显著下调,叶面多细胞腺毛的数量显著减少,但其具体调控机制还不清楚),而关于烟草的此类研究尚未见相关报道。
技术实现要素:
[0007]
本发明目的在于提供一种本氏烟分泌型腺毛调控基因nbjaz3,并利用其构建相应的表达载体、转化植物,以期解决当前对于烟草分泌型腺毛发生机制不明及缺乏有效的调控手段的技术问题,进而达到对烟草腺毛类型的定向改良和/或对腺毛发生密度进行调节的目的。
[0008]
为解决上述技术问题,本发明采用如下技术方案:本发明研究从本氏烟基因组中鉴定出10个jaz基因,并对这10个基因进行了进化树和共线性分析,分别命名为nbjaz1
‑
nbjaz10,并完成nbjaz基因家族的生物信息学分析。本发明首次克隆到nbjaz3基因,分别利用组织表达分析、亚细胞定位和转基因植株构建等方法明确nbjaz3负调控烟草分泌型腺毛的发生的功能,为多细胞分泌型腺毛的发生提供重要的理论基础。
[0009]
本发明首次构建了过量表达载体pcamia2300
‑
nbjaz3
‑
oe和基因沉默载体nbjaz3
‑
rnai,并通过叶盘转化法将其导入本氏烟;研究了过量表达株系、基因沉默株系和野生型对照株系的腺毛密度。
[0010]
所述本氏烟分泌型腺毛发生负调控基因nbjaz3能明显降低烟草腺毛的密度。研究结果表明,nbjaz3在腺毛中大量表达,并受meja诱导后上调表达。亚细胞定位分析发现该基因定位于细胞核和细胞膜。nbjaz3过量表达株系的分泌型腺毛密度显著降低,腺毛发生相关基因nbcycb2和nbmixta2的表达水平均显著降低,而基因沉默株系的腺毛密度没有明显改变,nbcycb2和nbmixta2基因的表达水平也没有显著变化。
[0011]
本发明研究运用酵母双杂交、双分子荧光互补、双荧光素酶报告系统等试验验证了nbjaz3与nbwo存在体内体外蛋白互作关系,并得到一个新的ja介导的调控分泌型腺毛发生的分子模型nbjaz3
‑
nbwo
‑
nbcycb2。
[0012]
与现有技术相比,本发明的主要有益技术效果在于:1. 本发明筛选、鉴定出了本氏烟分泌型腺毛调控基因nbjaz3,基于其过表达和基因沉默株系的表型和生理特征鉴定、检测研究表明,其能够负调控烟草分泌型腺毛的发生,具体而言,其在调控腺毛发生密度、定向改良腺毛类型等方面具有重要的应用价值。
[0013]
2. 本发明有利于进一步研究揭示烟草分泌型腺毛的分子调控机制,并在烟草生产中加以利用。
附图说明
[0014]
图1为本氏烟腺毛负调控基因nbjaz3的编码区序列图。
[0015]
图2为本氏烟nbjazs进化树和组织表达热图分析图谱;其中,a图为采用邻接法进行nbjazs进化树分析;b图为10个nbjazs在子叶、幼叶、成熟叶片、腺毛、茎、去毛茎和根系中的表达热图分析,其中,nbjaz3在腺毛中大量表达;rt
‑
pcr结果采用2
‑
δδct
分析法;l25和nbef作为内参基因;子叶的表达水平最低,设置为1;采用单因素anova的fisher
’ꢀ
lsd法进行显著性差异分析,**:p<0.01,*:0.05<p<0.01。
[0016]
图3为本发明nbjaz3在不同浓度外源meja处理后的表达热图分析图谱;以ms培养基上生长的无菌苗为阴性对照,分别采用mock(0.8%乙醇)、20、50、100和200 μμ meja处理本氏烟叶片24 h后分析nbjaz3的表达水平。rt
‑
pcr结果采用2
‑
δδct
分析法;l25和nbef作为内参基因;采用单因素anova的fisher
’ꢀ
lsd法进行显著性差异分析,**:p<0.01,*:0.05<p<0.01。
[0017]
图4为本发明nbjaz3亚细胞定位结果图谱;其中,a图为nbjaz3定位于烟草细胞核和细胞膜;绿色荧光信号(gfp)在509nm波长处进行收集;b图为亚细胞定位载体pcxdg
‑
gfp
‑
nbjaz3和细胞核定位序列载体共表达转化烟草细胞的结果,表示了nbjaz3确实定位于细胞核;红色荧光信号(rfp)在532nm波长处进行收集。
[0018]
图5为本发明nbjaz3启动子活性分析图谱;其中,a图为pcxgus
‑
p
‑
gus
‑
nbjaz3载体构建示意图;b
‑
g图分别为nbjaz3启动子瞬时表达后烟草不同组织(幼苗、成熟叶、根、茎、花瓣和花托)的gus染色活性分析;成熟叶、茎和花瓣中的gus活性较高,幼苗中的gus活性较低。
[0019]
图6为本发明nbjaz3转基因株系腺毛表型分析对比图;a图为nbjaz3过量表达株系(oe
‑
2和oe
‑
6),nbjaz3基因编辑株系(rnai
‑
3和rnai
‑
6)以及对照株系的腺毛形态观察;b图为oe
‑
2、oe
‑
6、rnai
‑
3、rnai
‑
6和对照株系的腺毛密度统计分析;c图为oe
‑
2、oe
‑
6、rnai
‑
3、rnai
‑
6和对照株系中nbjaz3、nbcycb2和nbmixta2的表达水平分析;l25和nbef作为内参基因;采用单因素anova的fisher
’ꢀ
lsd法进行显著性差异分析,**:p<0.01。
[0020]
图7为本发明实施例中外源meja处理缓解nbjaz3对长柄分泌型腺毛发生的抑制作用的验证图谱;其中,a图是1 μm meja处理oe
‑
2、oe
‑
6、rnai
‑
3、rnai
‑
6和对照株系两周后的腺毛形态观察;b图是1 μm meja处理oe
‑
2、oe
‑
6、rnai
‑
3、rnai
‑
6和对照株系两周后腺毛密度统计分析;l25和nbef作为内参基因;采用单因素anova的fisher’lsd法进行显著性差异分析,**:p<0.01。
[0021]
图8为本发明nbjaz3作为nbwo上游唯一直接抑制因子的验证试验图谱;其中,a图是nbwo在20、50、100和200 μm meja处理24 h后的热图分析;b图是nbwo在子叶、幼叶、成熟叶、腺毛、茎、去毛茎和根中的表达水平分析,采用单因素anova的fisher
’ꢀ
lsd法进行显著性差异分析,**:p<0.01;c是10个nbjazs与nbwo酵母双杂交的结果,nbjazs的编码区序列与pgbkt7载体连接(标记为bd
‑
nbjazs),nbwo的编码区序列与pgadt7载体连接(标记为ad
‑
nbwo),含bd
‑
nbjazs与ad
‑
nbwo的酵母菌液共培养后分别涂布于sd/
‑
leu/
‑
trp(ddo)和sd/
‑
ade/
‑
his/
‑
leu/
‑
trp/x/a(x:x
‑
α
‑
gal,a:aba;qdo/x/a)培养基上,结果表明只有nbjaz3与nbwo存在蛋白互作关系;d图是nbjaz3与nbwo双分子荧光互补结果,nbjaz3与nbwo的编码区序列分别与psat6
‑
ceyfp
‑
c1和psat6
‑
neyfp
‑
c1载体连接构成重组质粒nbjaz3
‑
cyfp和
nbwo
‑
nyfp;两个重组质粒共转化入烟草原生质体,同时以nbjaz3
‑
cyfp与nyfp、cyfp与
‑
nyfp为对照组。
[0022]
图9为本发明nbjaz3抑制nbwo调控nbcycb2的转录活性的验证图谱;a是双荧光素酶报告系统中的报告因子和效应因子的载体示意图,nbcycb2启动子片段连接入报告载体pgreen
‑
0800
‑
ii,nbjaz3和nbwo编码区序列连接入效应载体pcxdg
‑
ha;b图是报告因子和效应因子共转化后的luc相对活性分析;c图是pronbcycb2::gfp
‑
gus稳定转化烟草叶片的gus染色和活性分析,nbwo转化后叶片呈现深蓝色,且gus活性显著增加,而nbjaz3与nbwo共转化后叶片颜色呈现部分浅蓝色,且gus活性显著降低,这表明nbjaz3抑制nbwo的表达;采用单因素anova的fisher
’ꢀ
lsd法进行显著性差异分析,**:p<0.01。
具体实施方式
[0023]
下面结合附图和实施例来说明本发明的具体实施方式,但以下实施例只是用来详细说明本发明,并不以任何方式限制本发明的范围。
[0024]
在以下实施例中所涉及的质粒载体等如无特别说明,均为常规商业载体;所涉及的试剂如无特别说明,均为市售常规试剂;所涉及的试验、检测方法,如无特别说明,均为常规方法。
[0025]
实施例一:本氏烟分泌型腺毛调控基因nbjaz3的克隆及功能验证路线本氏烟的表皮毛全部是分泌型,按照表皮毛腺柄长度可分为长柄和短柄分泌型腺毛,因此,利用本氏烟为材料,探索ja调控分泌型腺毛发生的分子机制具有重要的研究价值。
[0026]
本发明研究采用同源克隆技术,利用茄科基因组网站本氏烟(nicotiana benthamiana)基因组序列进行反转录pcr(reverse transcription
‑
polymerase chain reaction,rt
‑
pcr),以本氏烟叶片总rna反转录合成cdna第一链为模板,首次扩增了本氏烟腺毛发生负调控基因nbjaz3,该基因完整开放阅读框序列全长1044 bp,编码347个氨基酸。
[0027]
采用rt
‑
pcr技术验证了nbjaz3基因在腺毛中大量表达,并受meja诱导后上调表达。构建pcxdg
‑
gfp
‑
nbjaz3亚细胞定位载体,通过瞬时转化烟草叶片,发现nbjaz3定位于细胞核和细胞膜。
[0028]
为了一步研究本氏烟nbjaz3在腺毛发生过程中的具体功能,本发明研究构建了pcamia2300
‑
nbjaz3
‑
oe过量表达载体和nbjaz3
‑
rnai基因沉默载体,通过叶盘转化法将其导入本氏烟。结果发现,nbjaz3过量表达株系的分泌型腺毛密度显著降低,而nbjaz3基因沉默株系的分泌型腺毛密度没有改变。rt
‑
pcr分析腺毛发生调控基因表达水平,结果显示nbcycb2和nbmixta2表达量在nbjaz3过量表达株系均显著下降,而在nbjaz3基因沉默株系中未发生明显改变。另外,发明人验证了外源meja能缓解nbjaz3对分泌型腺毛发生的抑制作用。
[0029]
采用酵母双杂交、双分子荧光互补、双荧光素酶报告系统等试验验证了nbjaz3与nbwo存在体内体外蛋白互作关系,并提出一个新的ja介导的分泌型腺毛发生的分子模型nbjaz3
‑
nbwo
‑
nbcycb2。
[0030]
本发明所公开的应用是基因nbjaz3负调控烟草分泌型腺毛发生的应用以及nbjaz3通过抑制nbwo调控nbcycb2的转录活性影响分泌型腺毛发生的应用。
[0031]
以下是本氏烟腺毛发生负调控基因nbjaz3的编码区序列以及负调控腺毛发生的实验验证的步骤方法。
[0032]
a、本发明课题组前期研究分析发现,外源meja处理烟草后诱导腺毛大量发生。在此基础上,利用同源克隆技术,以本氏烟叶片总rna反转录合成cdna第一链为模板,扩增得到了本氏烟腺毛负调控基因nbjaz3序列,该本氏烟腺毛负调控基因nbjaz3的编码区序列如图1所示,图中方框分别表示tify和jas保守结构域,红色字母表示两个结构域的典型氨基酸特征:b、采用rt
‑
pcr技术验证了nbjaz3基因在腺毛中大量表达,并受meja诱导后上调表达,参见图2
‑
3。
[0033]
c、构建pcxdg
‑
gfp
‑
nbjaz3亚细胞定位载体,通过瞬时转化烟草叶片,发现nbjaz3定位于细胞核和细胞膜,参见图4。
[0034]
d、构建pcxgus
‑
p
‑
gus
‑
nbjaz3启动子gus载体,通过瞬时转化烟草叶片,发现nbjaz3在成熟叶、茎和花瓣中的大量表达,参见图5。
[0035]
e、将本氏烟腺毛负调控基因nbjaz3序列的完整开放阅读框插入camv35s启动子下游,构建了植株过量表达载体;利用nbjaz3序列中特异性较好的324bp(105 bp
‑
428 bp)片段进行基因干扰试验,构建了基因沉默载体。将这两个重组载体通过农杆菌介导的叶盘转化法导入野生型本氏烟,筛选获得了表现型良好的t2代nbjaz3过量表达和基因沉默株系。
[0036]
f、鉴定了nbjaz3过量表达株系(oe
‑
2和oe
‑
6)的腺毛密度显著降低,而nbjaz3基因编辑株系(rnai
‑
3和rnai
‑
6)的腺毛密度没有明显增加。rt
‑
pcr分析发现nbcycb2和nbmixta2表达量在nbjaz3过量表达株系中均显著下降,而在nbjaz3基因沉默株系中未发生明显改变(参见图6
‑
7);另外,验证了外源meja能缓解nbjaz3对分泌型腺毛发生的抑制作用。以上结果都表明本氏烟腺毛发生负调控基因nbjaz3在植物分泌型腺毛发生过程中起重要作用。
[0037]
g、构建bd
‑
nbjazs和ad
‑
nbwo酵母双杂交载体,以及nbjaz3
‑
cyfp和nbwo
‑
nyfp载体,酵母双杂交和双分子荧光互补试验结果表明nbjaz3与nbwo存在蛋白互作关系,参见图8。
[0038]
h、构建报告因子和效应因子载体,双荧光素酶报告系统显示nbjaz3抑制nbwo调控nbcycb2的转录活性;另外,发明人将nbwo农杆菌菌液、nbjaz3与nbwo混合农杆菌菌液分别注射到pronbcycb2::gfp
‑
gus稳定表达烟草叶片上,gus染色与活性分析结果显示nbjaz3抑制nbwo的表达,参见图9。
[0039]
实施例二:nbjaz3基因的组织表达和应答meja处理的表达模式分析研究本发明前期研究表明1和5 μmmeja处理2周后,本氏烟植株逐渐矮化,10 μm meja处理2周后导致本氏烟植株死亡。因此,本发明研究采集1 μm meja处理2周后的本氏烟叶片为材料进行腺毛形态观察。结果发现长柄分泌型腺毛数量显著增加,而短柄型分泌型腺毛数量没有明显变化;另外,腺毛长度也没有发生显著变化。这些结果表明ja信号在烟草分泌型腺毛发生过程中具有重要作用。
[0040]
jaz蛋白作为ja信号核心抑制子通过调控下游相关基因的表达参与植物多种生长发育过程。因此,本发明研究筛选获得了17个本氏烟jaz蛋白,对其进行进化树分析发现,本氏烟jaz蛋白可聚类成10组,分别命名为nbjaz1
‑
nbjaz10(图2a)。采用rt
‑
pcr分析10个
nbjazs在子叶、幼叶、成熟叶、腺毛、茎、去毛茎和根系中的表达水平,结果显示nbjaz1,nbjaz3,nbjaz6,nbjaz8,nbjaz9和nbjaz10在腺毛中表达量较高。nbjaz1,nbjaz3,nbjaz5,nbjaz6,nbjaz8,nbjaz9和nbjaz10在叶片和茎中表达量较高。其中,nbjaz3在腺毛中的表达水平显著高于叶片和茎。
[0041]
对nbjaz3进行meja应答反应分析,结果如图3所示,以不同浓度meja处理本氏烟叶片24 h期间,nbjaz3的表达水平均高于对照,并在100 μm处理6 h时达到最大值。这些结果表明nbjaz3受meja诱导后能快速表达。
[0042]
实施例三:nbjaz3定位验证研究构建了pcxdg
‑
gfp
‑
nbjaz3亚细胞定位载体,通过瞬时转化将载体转化本氏烟叶片中,利用激光共聚焦显微镜观察发现,绿色荧光聚集在细胞核和细胞膜(见图4),这表明nbjaz3定位于细胞核和细胞膜。为了进一步验证试验结果,将pcxdg
‑
gfp
‑
nbjaz3载体和细胞核定位载体共表达转化烟草叶片,发现绿色荧光和红色荧光能重叠定位于细胞核,这表明nbjaz3确实定位于细胞核。
[0043]
实施例四:nbjaz3启动子在成熟组织中大量表达的验证研究构建pcxgus
‑
p
‑
gus
‑
nbjaz3启动子gus载体,通过瞬时转化本氏烟不同组织(幼苗、成熟叶、根、茎、花瓣和花托),gus染色分析发现nbjaz3在成熟叶、茎和花瓣中的gus活性较高,幼苗中的gus活性较低(见图5)。
[0044]
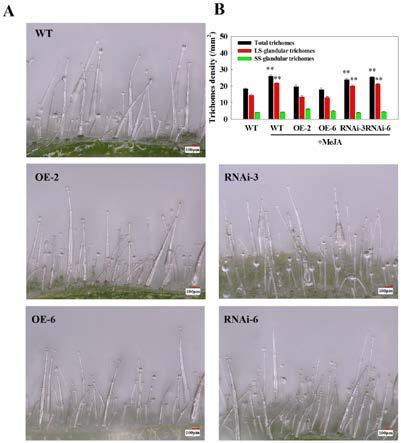
实施例五:nbjaz3负调控分泌型腺毛的发生的验证研究为了进一步验证nbjaz3在烟草分泌型腺毛发生中的作用,利用超景深显微镜分别观察nbjaz3过量表达株系(oe
‑
2和oe
‑
6),nbjaz3基因编辑株系(rnai
‑
3和rnai
‑
6)和野生型本氏烟(wt)的腺毛生长情况(见图6a、图6b);数据分析显示oe
‑
2和oe
‑
6的腺毛总数显著降低,分别是wt的0.64和0.67倍;长柄分泌型腺毛的密度也明显降低,分别是wt的0.55和0.45倍;然而rnai
‑
3和rnai
‑
6的腺毛总数和长柄分泌型腺毛数量没有明显变化。
[0045]
通过qrt
‑
pcr检测了腺毛发生相关基因在nbjaz3过量表达株系(oe
‑
2和oe
‑
6),nbjaz3基因编辑株系(rnai
‑
3和rnai
‑
6)和野生型本氏烟(wt)中的表达量(见图6c),结果表明nbcycb2的表达水平在oe
‑
2和oe
‑
6中显著降低,分别是对照的0.51和0.21倍;nbmixta2的表达水平在oe
‑
2和oe
‑
6中也显著降低,分别是对照的0.26和0.41倍;但是,nbcycb2和nbmixta2的表达水平在rnai
‑
3和rnai
‑
6中都没有显著改变。这些结果表明nbjaz3通过抑制nbcycb2和nbmixta2的表达,从而负调控烟草分泌型腺毛的发生。
[0046]
另外,对nbjaz3过量表达株系(oe
‑
2和oe
‑
6),nbjaz3基因编辑株系(rnai
‑
3和rnai
‑
6)和野生型本氏烟(wt)喷施1 μm meja,发现所有植株的腺毛数量均增加,其中,oe
‑
2和oe
‑
6的腺毛密度与未处理的wt的腺毛密度相近。这些结果表明外源meja有效缓解nbjaz3对分泌型腺毛发生的抑制作用(见图7)。
[0047]
实施例六: nbjaz3通过抑制nbwo调控nbcycb2的转录活性影响分泌型腺毛的发生的验证研究发明人课题组前期研究表明,nbwo基因沉默株系的腺毛密度显著降低,nbwo过量表达株系的腺毛密度没有明显改变,并验证了nbwo与nbcycb2蛋白之间存在负反馈调节关系;因此,本发明对nbwo是否参与ja信号进行进一步研究分析,结果显示nbwo的表达水平在20、50、100和200 μm meja处理后均增加(见图8);为了进一步验证nbwo在ja介导的腺毛发
生的过程中的作用,本发明研究构建bd
‑
nbjazs和ad
‑
nbwo酵母双杂交载体,将含bd
‑
nbjazs与ad
‑
nbwo的酵母菌液共培养后分别涂布于sd/
‑
leu/
‑
trp(ddo)和sd/
‑
ade/
‑
his/
‑
leu/
‑
trp/x/a(x:x
‑
α
‑
gal,a:aba;qdo/x/a)培养基上,发现只有nbjaz3与nbwo存在蛋白互作关系。构建nbjaz3
‑
cyfp和nbwo
‑
nyfp载体进行双分子荧光互补试验,将重组质粒共转化入烟草原生质体后发现nbjaz3与nbwo确实存在蛋白互作关系。
[0048]
前期研究表明,nbwo与nbcycb2蛋白之间存在负反馈调节关系,且nbcycb2负调控烟草腺毛的发生。为了进一步明确nbjaz3影响nbwo与nbcycb2之间的转录调控活性,发明人利用nbcycb2启动子片段构建报告载体,利用nbjaz3和nbwo的编码区序列构建效应载体,将两种载体分别转化烟草原生质体,发现含nbjaz3和nbwo的载体共转化后,luc相对活性显著降低。发明人利用nbcycb2启动子稳定表达烟草植株为材料,将含nbjaz3和nbwo菌液分别转化烟草叶片中,发现仅nbwo转化后叶片呈现深蓝色,gus活性显著增加,而nbjaz3与nbwo共转化后叶片颜色呈现部分浅蓝色,gus活性显著降低,这表明nbjaz3抑制nbwo的表达(参见图9)。
[0049]
上述实施例为本发明较佳的实施方式,但是,所属技术领域的技术人员能够理解,本发明的实施方式并不受上述实施例的限制,其它任何未背离本发明构思的精神实质与原理下所作的改变、修饰、替代、组合、简化,均应为等效的置换方式,都包含在本发明的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1