间作玉米下花生叶片基因差异表达的比较转录组分析方法
间作玉米下花生叶片基因差异表达的比较转录组分析方法
【技术领域】
1.本发明涉及生物技术领域,特别涉及间作玉米下花生叶片基因差异表达的比较转录组分析方法。
背景技术:
2.花生是我国重要油料作物和经济作物,花生间作是目前我国广泛采用的一种在时空上和生态上绿色高效高产的种植模式,且不同农作物之间的间作栽培模式非常丰富。其中,玉米间作花生具有传统单作不可比拟的优势,玉米和花生的生长发育在时空上互补,两者间作不仅可以通过改变农田生态结构,在时空上充分利用光、温、水、热及养分等资源,提高了间作胁迫下玉米对强光及花生对弱光的高效利用能力,促进间作作物对胁迫的应激能力、能量和物质的转化与积累,最终实现作物产量和生物量的增加以及土地总产出率。
3.在玉米间作花生的基础理论研究方面,研究表明,间作体系通过影响玉米和花生的根系生长与分布,提高两者对地下资源的充分利用效率,从而实现二者的间作优势,还可能是间作体系促进了豆科作物固定更多的氮,同时释放大量h+来促进根系活化与吸收磷元素,而磷素营养在植物光合作用系统中发挥重要调控作用;此外,玉米根系分泌物能高效活化土壤中难溶性铁以利于花生吸收而改善花生的铁营养;还有研究表明,玉米在强光胁迫下叶片对光能传递和转化的效率并没有明显增强,而间作花生改善玉米叶片适应强光的关键在于增强功能叶对co2的固定羧化能力。
4.相比单作,玉米间作花生优势明显,但该间作体系也会因玉米花生间对地下资源产生竞争性吸收和利用而引发各种胁迫(在此称之为“间作胁迫”),比如水分、养分和光资源等胁迫。在玉米花生间作体系中,花生对地上地下资源的竞争吸收和利用在整个生育期中一直处于劣势,尤其是对n、p和光照等资源的吸收和利用。虽然株高差异明显的玉米与花生的间作复合群体的产量明显提高,但高秆作物的玉米对矮秆作物的花生会引发时空上的生境胁迫和种间竞争,而由此引起的生境胁迫尤其是弱光胁迫将会严重影响矮秆作物的生长发育,并导致矮秆作物的生产力受到比较严重的影响。
5.前人就间作玉米胁迫对花生形态发育、光合特性和生理特性方面的影响研究已有相对较多报道,但对花生应答间作玉米胁迫的深层机理知之甚少,且花生应答间作玉米胁迫以调控其荚果膨大的机理也缺乏研究,更未见利用转录组测序对荚果膨大期花生功能叶应答间作玉米胁迫的关键差异表达基因及代谢通路进行发掘和分析的研究报道。为了深入地分析花生功能叶响应间作玉米胁迫以影响其荚果膨大的转录调控,有必要深入挖掘间作玉米胁迫下花生荚果产量形成的重要基因和关键代谢通路,为利用定向化学调控技术以减少间作玉米胁迫对花生产量造成的不利影响提供理论依据。
技术实现要素:
6.鉴于上述内容,有必要深入挖掘间作玉米胁迫下花生荚果产量形成的重要基因和关键代谢通路,为利用定向化学调控技术以减少间作玉米胁迫对花生产量造成的不利影响
提供理论依据。
7.为达到上述目的,本发明所采用的技术方案是:
8.对间作玉米下花生叶片基因差异表达的比较转录组分析的引物组,所述引物组的核苷酸序列如seq id no.1
‑
seq id no.36所示;所述核苷酸序列seq id no.1
‑
seq id no.2为内参基因引物对,所述核苷酸序列seq id no.3
‑
seq id no.36为17个重要代谢基因引物对;所述花生叶片选自花生荚果膨大期的功能叶。
9.本发明还包括所述的引物组在分析玉米和花生间作和/或花生单作的荚果膨大期花生功能叶基因差异表达分析中的应用。
10.进一步的,所述花生荚果膨大期为:花生种植后90d
‑
110d;花生种植区为广西全境。
11.本发明还包括所述对间作玉米下花生叶片基因差异表达的比较转录组分析的引物组的获取方法,所述方法为:
12.(1)培育花生参试材料:参试材料包括玉米与花生间作的实验组花生功能叶和单独种植花生的对照组花生功能叶;
13.(2)提取步骤(1)实验组和对照组两组花生功能叶的rna;构建cdna文库;
14.(3)对构建的文库进行测序、质量控制获得高质量的序列信息;
15.(4)将步骤(3)获得的序列信息与两个二倍体祖先野生种花生的全基因组序列进行比对,获取在参考基因组上的位置信息,获得mapped reads和非mapped reads,基于mapped reads进行后续生物信息学分析;
16.(5)通过mapped reads在参考基因组上的位置信息,使用cufflinks软件的cuffquant和cuffnorm组件,采用fpkm法对样品转录本和基因的表达水平进行定量;
17.(6)基于参考基因组序列,使用cufflinks软件对mapped reads进行拼接,寻找未被注释的转录区,发掘新基因;使用blast软件将发掘的新基因与nr、swiss
‑
prot、go、cog、kog、pfam和kegg数据库进行序列比对,使用kobas2.0得到新基因的kegg orthology结果;预测完新基因的氨基酸序列之后使用hmmer软件与pfam数据库比对,获得新基因的注释信息;
18.(7)根据比较转录组测序结果,有针对性地挑选与碳氮代谢、氨基酸代谢、脂类代谢、糖代谢和叶绿素代谢及植物激素信号转导和植物与病原菌互作相关的差异表达基因以及核糖磷酸
‑3‑
差向异构酶和甘油
‑3‑
磷酸
‑2‑
o
‑
酰基转移酶这两个代谢重要基因,以actin11为内参基因,设计得到如权利要求1所述的引物组。
19.进一步的,所述玉米与花生间作的实验组其所用玉米品种为桂单0810;其所用花生品种为桂花836。
20.本发明还包括所述对间作玉米下花生叶片基因差异表达进行分析的方法,所述方法为:提取花生功能叶的rna,合成cdna第一链,稀释20倍后的cdna第一链作为qrt
‑
pcr的扩增模板,使用2
‑
δδct
法计算样品差异基因的相对表达量;
21.所述qrt
‑
pcr反应中,核苷酸序列seq id no.1
‑
seq id no.2为内参基因actin 11的引物对;
22.核苷酸序列seq id no.3
‑
seq id no.36为17个重要代谢基因引物对。
23.上述17个重要代谢基因引物对的对应关系如下:
24.其中,核苷酸序列seq id no.3
‑
seq id no.4为araip.816xh基因的引物对;
25.其中,核苷酸序列seq id no.5
‑
seq id no.6为araip.1z65w基因的引物对;
26.其中,核苷酸序列seq id no.7
‑
seq id no.8为araip.a909a基因的引物对;
27.其中,核苷酸序列seq id no.9
‑
seq id no.10为araip.jq4v7基因的引物对;
28.其中,核苷酸序列seq id no.11
‑
seq id no.12为araip.i2m0y基因的引物对;
29.其中,核苷酸序列seq id no.13
‑
seq id no.14为araip.dlj08基因的引物对;
30.其中,核苷酸序列seq id no.15
‑
seq id no.16为araip.r1dvq基因的引物对;
31.其中,核苷酸序列seq id no.17
‑
seq id no.18为araip.gtw9x基因的引物对;
32.其中,核苷酸序列seq id no.19
‑
seq id no.20为araip.ugn58基因的引物对;
33.其中,核苷酸序列seq id no.21
‑
seq id no.22为aradu.nr4mv基因的引物对;
34.其中,核苷酸序列seq id no.23
‑
seq id no.24为aradu.0603j基因的引物对;
35.其中,核苷酸序列seq id no.25
‑
seq id no.26为aradu.qg8f6基因的引物对;
36.其中,核苷酸序列seq id no.27
‑
seq id no.28为aradu.wb4gb基因的引物对;
37.其中,核苷酸序列seq id no.29
‑
seq id no.30为aradu.z9z80基因的引物对;
38.其中,核苷酸序列seq id no.31
‑
seq id no.32为aradu.y1fv5基因的引物对;
39.其中,核苷酸序列seq id no.33
‑
seq id no.34为aradu.f99cn基因的引物对;
40.其中,核苷酸序列seq id no.35
‑
seq id no.36为newgene.2049基因的引物对;
41.本发明具有如下有益效果:
42.1、本发明通过对单作花生和玉米花生间作花生的转录组进行比较分析,研究花生功能叶应答玉米间作胁迫的分子机制,利用rna
‑
seq技术对花生单作与间作玉米胁迫下荚果膨大期花生功能叶的差异表达基因进行了比较分析;以代谢通路相关的差异基因为主,尤其是碳氮代谢、氨基酸代谢、脂类代谢、糖代谢和叶绿素代谢、核糖磷酸
‑3‑
差向异构酶和甘油
‑3‑
磷酸
‑2‑
o
‑
酰基转移酶代谢相关基因对花生种仁中的物质积累和荚果膨大期的叶片进行研究,挖掘出了间作玉米胁迫下花生荚果产量形成的重要基因和关键代谢通路,为利用定向化学调控技术以减少间作玉米胁迫对花生产量造成的不利影响提供理论依据;并通过qrt
‑
pcr检测17个候选差异表达基因(degs)的相对表达水平进行验证,最终表明qrt
‑
pcr检测结果与转录组测序分析中的倍数变化大体一致,采用转录组测序分析的方法准确、可行、有效、简单。
【附图说明】
43.图1是花生单作和玉米间作花生田间种植图示意图;
44.图2是花生样品之间的差异基因表达量相关性热图;
45.图3是间作玉米胁迫下花生功能叶转录组的差异表达基因聚类热图;
46.图4是间作玉米胁迫下花生功能叶转录组的差异表达基因火山图;
47.图5是间作玉米胁迫下花生功能叶转录组的差异表达基因ma图;
48.图6是间作玉米胁迫下花生功能叶转录组差异表达基因的cog注释图;
49.图7是间作玉米胁迫下花生功能叶转录组差异表达基因的eggnog注释图;
50.图8是间作玉米胁迫下花生功能叶转录组差异表达基因的go注释图;
51.图9是间作玉米胁迫下花生功能叶转录组差异表达基因的kegg注释图;
52.图10是间作玉米胁迫下花生功能叶转录组差异表达基因的通路富集图;
53.图11是代谢通路上17个关键差异基因的整体表达谱热图;
54.图12是碳代谢通路上的基因差异表达热图;
55.图13是植物激素信号转导通路上的基因差异表达热图;
56.图14是氨基酸的生物合成通路上的基因差异表达热图;
57.图15是卟啉与叶绿素代谢通路上的基因差异表达热图;
58.图16是黄酮类化合物的生物合成通路上的基因差异表达热图;
59.图17是植物与病原体互作通路上的基因差异表达热图;
60.图18是基于花生功能叶转录组测序的花生新基因的go注释图;
61.图19是基于花生功能叶转录组测序的花生新基因的eggnog注释图;
62.图20是基于花生功能叶转录组测序的花生新基因的nr注释图;
63.图21是17个候选基因的rna
‑
seq结果图;
64.图22是17个候选基因的qrt
‑
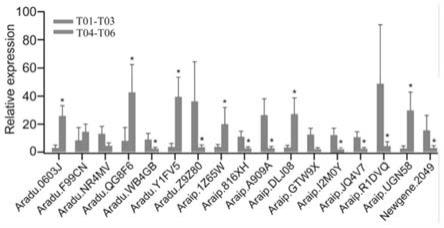
pcr的表达结果比较图。
【具体实施方式】
65.为使本发明的上述目的、特征和优点能够更加明显易懂,下面结合附图对本发明的具体实施方式做详细的说明。在下面的描述中阐述了很多具体细节以便于充分理解本发明。但是本发明能够以很多不同于在此描述的其它方式来实施,本领域技术人员可以在不违背本发明内涵的情况下做类似改进,因此本发明不受下面公开的具体实施的限制。
66.实施例1:
67.荚果膨大期花生功能叶应答间作玉米胁迫的比较转录组分析:
68.1、玉米间作花生体系建立:
69.花生品种:桂花836;该品种为广西主栽抗病高产适合间作的花生品种;
70.玉米品种:桂单0810;该品种为广西高产玉米品种;
71.实验地址:广西农业科学院武鸣里建基地;
72.花生种植时间:3月10日,花生荚果膨大期取样时间:6月15日。
73.土壤状况:全氮、全磷和全钾含量分别为0.113%、0.053%、0.24%,水解性氮88mg/kg,有效磷11.4mg/kg,速效钾110mg/kg,有机质18.5g/kg;沙红壤土,ph值7.1。
74.实验设计:采用随机区组设计,3次重复,设置花生单作(对照)和玉米间作花生(处理)两种种植模式:单作和间作的种植方向均为东西走向,间作采用4行玉米/6行花生的种植模式,单作和间作均采用宽窄行起垄种植,垄宽90cm(包两边沟),相同规格,即宽行距55cm、窄行距35cm,玉米穴距0.2m,每穴1株,花生穴距16.65cm,每穴2株,每个小区面积约为20m2;具体如图1所示:图1中的上行为花生单作种植的示意图;图1中下行为花生和玉米间作的示意图。
75.植玉米时,以450kg/hm2的复合肥(n:p2o5:k2o为15:15:15)为基肥,以300kg/hm2的复合肥和150kg/hm2的尿素(n为46%)为追肥;种植花生时,额外施用67.5kg/hm2的p2o5、67.5kg/hm2的k2o和750kg/hm2的钙镁磷肥。
76.在花生荚果膨大期,从花生单作种植模式每个小区中随机选取植株长势相对均一的花生功能叶10片混合为一个重复样品,分别记为t01、t02和t03;从玉米间作花生种植模
式每个小区中靠近玉米的4行花生中随机选取植株长势相对均一的花生功能叶10片混合为一个重复样品,分别记为t04、t05和t06。将收集到的花生功能叶样品用液氮迅速冷冻后保存于
‑
80℃超低温冰箱中并用于转录组测序。
77.2、cdna文库构建、rna
‑
seq测序与数据质控:
78.采用天根rna试剂盒提取花生功能叶的总rna,利用nanodrop检测rna的纯度和浓度,利用agilent2100仪器精确检测rna的完整性。样品检测合格后,进行文库构建,其主要流程如下:
79.①
用带有oligo(dt)的磁珠富集mrna;
80.②
加入fragmentation buffer将mrna进行随机打断;
81.③
以mrna为模板,用六碱基随机引物(random hexamers)合成第一条cdna链,然后加入缓冲液、dntps、rnase h和dna polymerase i合成第二条cdna链,利用ampure xp beads纯化cdna;
82.④
纯化的双链cdna再进行末端修复、加a尾并连接测序接头,然后用ampure xp beads进行片段大小选择;
83.⑤
最后通过pcr富集得到cdna文库。
84.在测序之前,先使用qubit2.0进行初步定量,接着利用agilent 2100对文库的insert size进行检测,然后通过q
‑
pcr方法对文库的有效浓度进行准确定量,此后,不同文库按照目标下机数据量进行pooling,利用illumina hiseq平台进行测序。获得的原始测序数据需要进行严格的质量控制,采用以下过滤标准:
85.①
去除含有接头的reads;
86.②
去除低质量的reads(包括去除n的比例大于10%的reads;去除质量值q≤10的碱基数占整条read的50%以上的reads),经过质量控制之后得到的高质量的clean data。
87.3、转录组数据与参考基因组序列比对:
88.本发明以两个二倍体祖先野生种花生的全基因组序列(https://peanutbase.org/)作为参考基因组,利用tophat2将过滤后得到的clean reads与参考基因组进行序列比对,获取在参考基因组上的位置信息,获得比对上的读长数目和非比对上的读长数目,基于比对上的读长数目进行后续生物信息学分析。
89.4、差异基因表达分析与功能注释:
90.通过比对上的读长数目在参考基因组上的位置信息,使用cufflinks软件的cuffquant和cuffnorm组件,采用fpkm法对样品转录本和基因的表达水平进行定量,将差异倍数(fold change,fc)≥2且错误发现率(false discovery rate,fdr)<0.01作为差异表达基因的筛选标准,并根据相对表达量将其划分为上调基因和下调基因,同时对差异基因进行go、kegg和cog等多种数据库功能注释和富集分析。其中,fpkm计算公式如下:
[0091][0092]
5、新基因发掘与功能注释:
[0093]
基于参考基因组序列,使用cufflinks软件对比对上的读长数目进行拼接,通过过滤掉编码少于50个氨基酸残基的过短肽链或只包含单个外显子的序列,并与参考基因组注释信息进行比较,寻找未被注释的转录区,发掘样品的新转录本和新基因,并使用blast软
件将发掘的新基因与nr、swiss
‑
prot、go、cog、kog、pfam和kegg数据库进行序列比对,使用kobas2.0得到新基因的kegg orthology结果,预测完新基因的氨基酸序列之后使用hmmer软件与pfam数据库比对,获得新基因的注释信息。
[0094]
对测序结果进行注释信息的分析如下:
[0095]
1、花生功能叶转录组测序数据评估:
[0096]
单作和间作下的三个样本的测序原始数据组成如表1所示:
[0097]
表1测序原始数据组成
[0098][0099]
注:干净读长百分比为:过滤后的reads数目占总reads数目的百分比;含有接头读长百分比为:含有接头reads数目占总reads数目的百分比;低质量读长百分比为:过滤掉的低质量reads数目占总reads数目的百分比。
[0100]
单作下的三个样本的测序数据指标参数统计结果如表2所示:
[0101]
表2测序数据指标参数统计结果
[0102][0103]
注:干净读长数目,指clean data中pair
‑
end reads数目;干净读长碱基数目,指clean reads的总碱基数目;gc含量(%),指clean data中g和c数目占总碱基数目的百分比;≥q20:clean data中质量值大于或等于20的碱基所占的百分比;≥q30:clean data中质量值大于或等于30的碱基所占的百分比。
[0104]
表1
‑
表2的结果表明,测序获得的序列有效性和可利用比列较高,整体质量优良,测序碱基错误率很低、测序质量高,测序结果可靠,所获数据合格、结果可靠,可用于后续生物信息学分析。
[0105]
2、花生功能叶转录组测序数据与参考基因组序列的比对:
[0106]
利用tophat2软件将过滤获得的样品单末端clean reads与两个二倍体祖先野生种花生基因组(https://peanutbase.org/)进行比对分析。结果如表3所示:
[0107]
表3样品测序reads与花生参考基因组序列的比对结果统计
[0108][0109]
注:单端干净读长数目指clean reads数目,按单端计(
×
2);比对上的读长数目指比对到花生参考基因组上的reads数目及其在占单端干净读长数目的百分比(包括单端比对和双端比对);单一比对上的读长数目指比对到花生参考基因组唯一位置的reads数目及其占单端干净读长数目的百分比;多重比对上的读长数目指比对到花生参考基因组多处位置的reads数目及其占单端干净读长数目的百分比;比对上正链的读长数目指比对到花生参考基因组正链上的reads数目及其占单端干净读长数目的百分比;比对上负链的读长数目指比对到花生参考基因组负链上的reads数目及其占单端干净读长数目的百分比。
[0110]
由表3可知,单作下的三个样本的单端干净总读长数目为147450600,平均每个样本的单端干净读长数目为49150200;间作下的三个样本的单端干净总读长数目为142836012,平均每个样本的单端干净读长数目为47612004。单作下的三个样本的比对上的总读长数目为115903364,平均每个样本的比对上的读长数目为38634455,其平均比对率为78.61%;间作下的三个样本的比对上的总读长数目为112349639,平均每个样本的比对上的读长数目为37449880,其平均比对率为78.66%。单作下的三个样本的单一比对上的总读长数目为97017431,平均每个样本的单一比对上的读长数目为32339144,其平均比对率为65.81%;间作下的三个样本的单一比对上的总读长数目为93803465,平均每个样本的单一比对上的读长数目为31267822,其平均比对率为65.67%。单作下的三个样本的多重比对上的总读长数目为18885933,平均每个样本的多重比对上的读长数目为6295311,其平均比对率为12.80%;间作下的三个样本的多重比对上的总读长数目为18546174,平均每个样本的
多重比对上的读长数目为6182058,其平均比对率为12.99%。单作下的三个样本的比对上正链的总读长数目为54748957,平均每个样本的比对上正链的读长数目为18249652,其平均比对率为37.13%;间作下的三个样本的比对上正链的总读长数目为53050645,平均每个样本的比对上正链的读长数目为17683548,其平均比对率为37.14%。单作下的三个样本的比对上负链的总读长数目为54712596,平均每个样本的比对上负链的读长数目为18237532,其平均比对率为37.11%;间作下的三个样本的比对上负链的总读长数目为52987776,平均每个样本的比对上负链的读长数目为17662592,其平均比对率为37.10%。总的来说,各样品的比对上的读长数目与参考基因组的比对效率在78.41%~78.93%之间,尚有约22%的新转录序列待开发。
[0111]
统计比对上的读长数目在花生参考基因组不同区域(外显子、内含子和基因间区)中的分布。结果如表4所示:
[0112]
表4样品测序mapped reads在花生参考基因组不同区域中的分布
[0113][0114]
由表4可知,单作下的三个样本分布在外显子中的比对上的读长数目占总数的78.36%~80.41%,平均79.63%,间作下的三个样本分布在外显子中的比对上的读长数目占总数的79.64%~80.69%,平均80.09%;单作下的三个样本分布在基因间区中的比对上的读长数目占总数的12.79%~13.53%,平均13.06%,间作下的三个样本分布在基因间区中的比对上的读长数目占总数的12.90%~13.25%,平均13.02%;单作下的三个样本分布在内含子中的比对上的读长数目占总数的6.79%~8.11%,平均7.31%,间作下的三个样本分布在内含子中的比对上的读长数目占总数的6.40%~7.15%,平均6.89%。
[0115]
3、花生功能叶响应间作玉米胁迫的差异基因表达分析:
[0116]
本发明采用fpkm法计算基因的表达量,并以|log2fc|≥1且fdr<0.01的条件来筛选差异表达基因。所筛选差异表达基因的层次聚类显示,单作荚果膨大期花生功能叶的三个生物学重复因具有相同或相似表达模式的差异基因而被聚为一类,间作玉米胁迫下荚果膨大期花生功能叶的三个生物学重复因具有相同或相似表达模式的差异基因而被聚为另一类(具体可参见图2)。经过rsem软件包筛选鉴定出167个差异表达基因,上调和下调表达基因数分别为111个和56个,在8个差异表达的新基因中,有3个上调表达,有5个下调表达结果如表4:
[0117]
表4间作玉米胁迫下花生功能叶转录组的差异表达基因概况
[0118][0119][0120]
由表4可知,对167个差异表达基因进行多种数据库功能注释(go、cog、kog、eggnog、kegg、pfam、swiss
‑
prot和nr),结果表明,有57个在cog数据库中得到了功能注释,以一般功能预测(r,17.59%)、无机离子转运与代谢(p,12.96%)、氨基酸转运与代谢(e,11.11%)、碳水化合物转运与代谢(g,9.26%)、转录(k,9.26%)、信号转导机制(t,8.33%)和复制、重组与修复(l,6.48%)为主(具体可参见图3
‑
5);有149个在eggnog数据库中得到了功能注释,以一般功能预测(r,25.49%)、未知功能(s,18.95%)、转录(k,11.11%)和无机离子转运与代谢(p,9.15%)为主(具体可参见图6
‑
7);有74个在kog数据库中得到了功能注释;有147个在pfam数据库中得到了功能注释,可编码氨基酸;有137个在swiss
‑
prot数据库中得到了功能注释,可翻译表达为蛋白质;有159个在nr数据库中得到了功能注释。
[0121]
4、花生功能叶响应间作玉米胁迫的差异表达基因的go注释与富集:
[0122]
对116个差异表达基因在go数据库中得到了功能注释,其中,有24个参与了不同生物学过程(biological process),有68个执行了不同分子功能(molecular function),剩下的24个参与形成了不同细胞组分(cellular component)(如图8所示)。为了进一步明确这些差异表达基因的具体功能,通过ks<0.05筛选,发现有22个差异表达基因显著富集在13个go条目上,主要涉及叶绿素结合(go:0016168)、细胞铁离子稳态与绑定(go:0006879和go:0005506)、植物细胞壁纤维素代谢过程(go:0052541)以及电子载体活性、信号转导与受体活性(go:0009055、go:0004871和go:0038023),还有定位于质体(go:0009536)、叶绿体(go:0009507)和线粒体(go:0005739)上的(表9)。此外,核糖磷酸
‑3‑
差向异构酶(go:0004750)和甘油
‑3‑
磷酸
‑2‑
o
‑
酰基转移酶(go:0090447)很可能是花生功能叶响应间作玉米胁迫的关键酶如表5所示:
[0123]
表5间作玉米胁迫下花生功能叶转录组差异表达基因的go注释与显著富集
[0124]
[0125][0126]
5、花生功能叶响应玉米间作胁迫的差异表达基因的kegg注释与富集:
[0127]
有63个差异表达基因在kegg数据库中得到了功能注释,为了进一步弄清间作玉米胁迫引发的花生功能叶差异表达基因的具体功能,经过kegg数据库对基因的生物代谢通路进行了系统分析,结果显示,63个差异表达基因中的34个参加了37条具体代谢通路,涉及到新陈代谢(metabolism)、遗传信息处理(genetic information processing)、环境信息处理(environmental information processing)、生物体系统(organismal systems)和细胞过程(cellular processes),其中,以代谢通路为主(表6和图9
‑
图10所示)。37条具体代谢通路包括23个差异表达基因参加的32条代谢通路、4个差异表达基因参加的间作玉米胁迫
引发的植物激素信号转导通路(ko04075)、3个差异表达基因参加的间作玉米胁迫触发花生叶片机体系统的植物与病原体互作通路(ko04626)、2个差异表达基因参加的细胞自噬通路(ko04145)、1个差异表达基因参加的植物生理节律通路(ko04712)及1个差异表达基因参加的rna转运通路(ko03013)具体如表6所示;另外,基于p<0.05的代谢通路显著性富集分析发现,黄酮类生物合成(ko00941)、戊糖和葡萄糖醛酸相互转化(ko00040)和卟啉和叶绿素代谢(ko00860)等3个代谢通路得到了显著富集。
[0128]
表6间作玉米胁迫下花生功能叶转录组差异表达基因的kegg注释与富集
[0129]
[0130]
[0131][0132]
根据上述通路构建基因差异表达热图,其中,图11为对23条代谢通路上的17个关键差异基因的整体表达谱热图;图12为碳代谢通路(ko01200)的表达谱热图;图13植物激素信号转导通路(ko04075)的表达谱热图;图14为氨基酸的生物合成通路(ko01230)的表达谱热图;图15为卟啉与叶绿素代谢通路(ko00860)的表达谱热图;图16为黄酮类化合物的生物合成通路(ko00941)的表达谱热图;图17为植物与病原体互作通路(ko04626)的表达谱热图。
[0133]
6、基于花生功能叶转录组测序的新基因发掘与功能注释:
[0134]
过滤掉编码少于50个氨基酸残基的过短肽链或只包含单个外显子的序列,使用cufflinks软件发掘到了单作和间作玉米胁迫下花生功能叶中总共有2299个未被注释的转录区,并对这些新基因进行了多种数据库功能注释(go、cog、kog、eggnog、kegg、pfam、swiss
‑
prot和nr)。结果如表7所示:
[0135]
表7基于花生功能叶转录组测序的花生新基因功能注释
[0136]
注释数据库gocogkogeggnogkeggpfamswiss
‑
protnrall新基因数目740227824146844586893615941624
[0137]
由表7可知,有445个新基因在kegg数据库中得到了功能注释,参与了新陈代谢(metabolism)、遗传信息处理(genetic information processing)、环境信息处理(environmental information processing)、生物体系统(organismal systems)和细胞过程(cellular processes),以新陈代谢通路为主;有227个新基因在cog数据库中得到了功能注释;有824个新基因在kog数据库中得到了功能注释;有868个新基因在pfam数据库中得到功能注释,可编码氨基酸;有936个新基因在swiss
‑
prot数据库中得到了功能注释,可翻译表达成蛋白质具体如表7所示
[0138]
具体结果如图18
‑
图20所示,如图18所示,总共有1624条新基因被注释到其中的一个或多个数据库,其中,有740个新基因在go数据库中得到了功能注释,分别参与了不同生物学过程(biological process)、执行了不同分子功能(molecular function)和参与形成了不同细胞组分(cellular component);如图19所示,有1468个新基因在eggnog数据库中得到了功能注释,以未知功能(s,28.08%)、一般功能预测(r,27.68%)、转录(k,6.97%)、信号转导机制(t,6.9%)和翻译后的修饰、蛋白周转、分子伴侣(o,5.56%)为主;如图20所示,有1594个新基因在nr数据库中得到了功能注释,以大豆(glycine max,23.40%)、菜豆
(phaseolus vulgaris,11.92%)、鹰嘴豆(cicer arietinum,10.79%)、蒺藜苜蓿(medicago truncatula,10.60%)和野生大豆(glycine soja,9.72%)为主。
[0139]
上述结果表明,在确保花生植株各时期水肥正常需求的情况下,相对于花生单作种植模式,间作玉米胁迫致使荚果膨大期花生功能叶响应基因在转录水平上发生了明显变化,且基因差异表达以上调为主(111/167),但在重要代谢通路上的关键基因差异表达上,以下调为主,这些基因差异表达很可能影响着间作花生的生物量或者干物质积累;同时,本发明发现,定位于质体和线粒体上的差异基因的表达均显著下调,叶绿体上的差异基因的表达也以显著下调为主(7/9),这很可能是间作遮荫胁迫下花生功能叶的细胞器结构、数目和功能下降的转录证据,尤其是叶绿体。叶绿素是植物进行光合作用的保障,负责吸收、传递和转化光能,基因aradu.m64bq(go:0016168)的表达下调引起了结合态叶绿素含量的降低,且卟啉和叶绿素代谢通路(ko00860)上的差异基因表达也下调,引起游离态叶绿素含量增加,进而造成细胞光氧化损伤,不利于光合作用。虽然信号受体活性(go:0038023)和信号转导活性(go:0004871)的差异基因表达上调,但该基因的上调表达还影响了生色团结合蛋白(go:0018298)和天线蛋白(ko00196)的含量和活性,而它们与植物光系统ⅱ中光能捕获及光能转化的正常运转密切相关,这或许是间作玉米胁迫虽然提高了花生叶绿素含量及其对弱光利用能力,但单产始终不如花生单作种植的内在原因。此外,影响花生功能叶中铁离子状态的基因表达也下调(go:0006879和go:0005506),而铁和镁在鳌合酶的作用下参与叶绿素a的合成,且花生体内铁离子状态也可能是间作玉米改善花生铁营养重要原因之一。总之,间作胁迫下花生叶绿素代谢紊乱,尤其发生了降解代谢而引起细胞光氧损伤可能是叶片净光合速率下降的主要原因,间作玉米胁迫通过抑制叶绿素代谢来影响花生功能叶的光合作用。
[0140]
花生是以收获荚果为主的重要油料作物,其氨基酸、脂质和碳水化合物等物质含量丰富,而它们的生物合成与代谢对花生产量和品质形成尤为重要。质体、叶绿体和线粒体是植物物质代谢和能量转化的最重要场所,其生理特性易受环境条件的影响。
[0141]
定位于质体、叶绿体和线粒体上的转录基因表达受间作胁迫影响,其中,叶绿体光合碳代谢重要酶核糖磷酸
‑3‑
差向异构酶(go:0004750)的编码转录基因表达下调,脂质合成代谢途径中的限速酶甘油
‑3‑
磷酸
‑2‑
o
‑
酰基转移酶(go:0090447)的编码转录基因表达上调。核糖磷酸
‑3‑
差向异构酶是卡尔文循环再生阶段的一种重要酶类,在co2同化中发挥着重要作用,有证据表明,超表达rpease基因增加了植物体内碳水化合物的含量,促进植物的生长发育、光合速率以及生物量的积累,也增强了植株对逆境的抵抗力。在本发明中,间作玉米胁迫限制了花生叶片光合作用,这主要归因于叶绿体中碳固定(ko00710)和氮代谢(ko00910)通路上的基因表达下调,同时作为氮代谢主要底物的氨基酸合成与代谢也受间作玉米胁迫抑制,且研究发现一个新转录基因(newgene.2049)下调了丙氨酸、天冬氨酸、谷氨酸、精氨酸和脯氨酸等氨基酸代谢,且下调脯氨酸不利于提高花生的遮荫耐受性,然而线粒体中氧化磷酸化反应的调控转录基因上调提供了atp,使得遮阴下的光合作用得到缓解,也为荚果膨大期花生的源—库提供保障。由此可见,作物可以通过碳氮代谢的协同作用来适应外界环境胁迫。
[0142]
油脂是花生最重要的营养物质,甘油
‑3‑
磷酸
‑2‑
o
‑
酰基转移酶(gpat)的活性受基因表达的影响,定位于叶绿体和内质网,催化脂质合成代谢的第一步,对花生种子含油量意
义重大。本发明发现,在间作玉米遮阴胁迫下,影响gpat酶活的转录基因表达上调,且该酶主导的脂质代谢也以上调为主,尤其是甘油脂质和甘油磷脂代谢途径,其中定位于叶绿体的代谢产物是叶绿体膜脂的基本骨架成分,而定位于内质网的代谢产物则经去磷酸化后转运到叶绿体与pi作用合成膜脂成分以补充和维持膜系统的完整性,这对维持花生叶片光系统的正常功能至关重要,这可能是间作遮阴下磷改善花生光合特性以提高产量的重要证据。此外,有证据表明gpat是一种含铁酶,而花生叶片中铁离子状态和平衡受间作遮阴胁迫的不利影响(go:0006879和go:0005506),这又在一定程度上限制了gpat的活性,进而导致叶片光合作用和脂质代谢受不同程度的影响,最终不利于油脂的积累。由gpat调节的脂质代谢还影响作物花粉育性和种子结实率,对植株的抗逆性和生长发育也发挥重要作用。
[0143]
除上述叶绿体上、碳氮代谢及脂质代谢上的差异表达基因响应间作玉米胁迫外,还有植物激素信号转导通路(ko04075)、植物与病原体互作通路(ko04626)、黄酮类生物合成通路(ko00941)上的差异表达基因也响应了间作玉米胁迫。值得一提的是,苯丙素生物合成通路(ko00940)上的差异表达基因也参与了应答间作玉米胁迫,苯丙素是响应非生物逆境的重要物质,参与了植物细胞抗氧化物活性的调控,很可能在间作花生生长过程中具有重要作用。
[0144]
实施例2:
[0145]
差异表达基因的qrt
‑
pcr验证
[0146]
为了验证转录组测序的可靠性,有针对性地挑选17个差异表达基因(degs)进行实时荧光定量pcr检验,根据实施例1比较转录组测序结果,挑选17个与碳氮代谢、氨基酸代谢、脂类代谢、糖代谢和叶绿素代谢以及植物激素信号转导和植物与病原菌互作相关的差异表达基因,加上核糖磷酸
‑3‑
差向异构酶和甘油
‑3‑
磷酸
‑2‑
o
‑
酰基转移酶这两个重要基因,选用actin11为内参基因。设计特异引物,引物序列如表8:
[0147]
表8重要差异表达基因引物对(具体参见seq id no.1
‑
seq id no.36)
[0148][0149][0150]
采用上述引物对玉米间作花生和单作花生进行扩增、测序检验的方法如下:
[0151]
提取样品rna,采用大连宝生物公司的逆转录试剂盒合成cdna第一链,稀释20倍后的cdna第一链作为qrt
‑
pcr的扩增模板,qrt
‑
pcr根据sybr premix ex taq
tm kit(takara,dalian,china)的使用说明书进行。扩增反应体系:sybr premix ex taq(1
×
)5μl、cdna 2μl、正反向引物各0.5μl(10μm),补水至总体积10μl。扩增反应程序:95℃,2min;40个循环的95℃,10s;60℃,30s。使用2
‑
δδct
法计算样品差异基因的相对表达量。所用仪器为abi 7500real
‑
time pcr系统。每个样品每个基因均采用三次生物学重复和三次技术性重复。得到结果如图21
‑
22所示:
[0152]
qrt
‑
pcr的结果与rna
‑
seq结果在表现上大体一致,不仅说明了转录组测序获得的基因差异表达信息具有较高的可重复性和准确性,是可信的,而且说明转录组测序数据能较为准确地反映荚果膨大期花生功能叶对间作玉米胁迫的反应。另外,核糖磷酸
‑3‑
差向异构酶和甘油
‑3‑
磷酸
‑2‑
o
‑
酰基转移酶也是花生应答间作玉米胁迫的重要调控酶,并与花生种仁中的物质积累和荚果膨大密切相关。qrt
‑
pcr验证差异表达基因的结果与转录组测序结果是基本一致的。
[0153]
实施例3:
[0154]
按实施例1的方法设计实验后,在花生荚果膨大期,从花生单作(对照)的每个小区选择正常生长的花生植株3株,从玉米间作花生(处理)的每个小区的每一行选择正常生长的花生植株3株(共18株),分别调查和测定花生单作和玉米间作花生两种种植模式下花生植株的株高和侧枝长以及花生功能叶叶绿素spad值和净光合速率,花生主茎倒三叶功能叶的叶绿素spad值使用日本产spad
‑
502型叶绿素仪测定,花生主茎倒三叶功能叶的净光合速
率pn值使用美国cid公司生产的ci
‑
340便携式光合测定仪于上午9:30~12:30时间段内测定。
[0155]
另外,在饱果成熟期,每个小区选正常生长的花生植株6株,分别调查花生单作(对照)和玉米间作花生(处理)两种种植模式下花生植株的结果性状和经济性状。得到的结果如下:
[0156]
表9间作玉米胁迫对花生植株形态和功能叶光合作用的影响
[0157]
处理株高(cm)侧枝长(cm)叶绿素spad值净光合速率pn(μmol.m
‑2.s
‑1)单作39.39ba42.22bb38.79a23.83a间作54.78aa57.17aa37.10a22.58a
[0158]
由表9可知,花生植株平均株高和平均侧枝长在间作玉米胁迫处理下比对照单作分别增加了39.07%和35.41%,平均株高在处理与对照之间的差异达到显著水平(p<0.05),侧枝长在处理与对照之间的差异达到极显著水平(p<0.01),表明间作玉米胁迫处理可明显促进花生植株的伸长。间作玉米胁迫处理下花生的平均叶绿素spad值和净光合速率pn值略低于对照单作花生,间作玉米胁迫处理下花生的平均叶绿素spad值和净光合速率pn分别比对照单作花生降低了4.36%和5.25%,处理与对照之间的差异不显著,表明间作玉米胁迫处理对花生功能叶叶绿素合成和净光合速率影响不大。
[0159]
本实施例还研究间作玉米胁迫对花生相关结果性状和经济性状的影响,结果如表10:
[0160]
表10间作玉米胁迫对花生相关结果性状和经济性状的影响
[0161]
处理平均饱果数(个)百果重(g)百仁重(g)小区产量(kg)单作93.15a203.36a80.29a5.05a间作88.64a195.78a74.31b4.31b
[0162]
注:小区面积为13.34m2。
[0163]
由表10可知,间作玉米胁迫处理下和对照单作下花生的平均饱果数差异不显著;平均百果重差异不显著;间作玉米胁迫处理下和对照单作下花生的平均百仁重的差异达到显著水平;平均小区产量的差异达到显著水平。结果表明,间作玉米胁迫处理对花生饱果数和百果重影响不大,但对花生百仁重和小区产量影响较大,一定程度上抑制花生种仁中的物质积累和荚果膨大。
[0164]
上述结果表明,在玉米间作花生种植模式下,苗期花生与玉米共生,但随着玉米和花生的生长发育,后期玉米株高明显高于花生,花生冠层的透光率降低,导致花生出现避荫反应,具体表现为花生植株增高,茎秆变纤细,节间过度伸长,抗折力下降,易倒伏,叶片变薄,干物质积累量显著降低,最终在一定程度上影响了花生的生长发育及其产量和品质。相比单作花生,玉米
‑
花生间作可以明显促进花生植株伸长,轻微降低其叶片叶绿素spad值和净光合速率pn值但影响不大,表明间作玉米胁迫处理可明显促进花生植株的伸长,但由于在4行玉米/6行花生的种植模式下花生的生长空间足够大,间作花生所在的生长空间透光和通气效果好,玉米对花生的遮荫胁迫不明显,进而对花生功能叶叶绿素合成和光合作用影响不大。另外,间作玉米胁迫对花生饱果数和百果重影响不大,但对百仁重影响较大,一定程度上可以抑制种仁中的物质积累和荚果膨大,这可能由于间作玉米胁迫影响了花生荚果膨大过程中的碳氮代谢、氨基酸代谢、脂类代谢和糖代谢。另外,间作玉米胁迫对花生饱
果数和百果重影响不大,但对百仁重影响较大,一定程度上可以抑制种仁中的物质积累和荚果膨大,这可能由于间作玉米胁迫影响了花生荚果膨大过程中的碳氮代谢、氨基酸代谢、脂类代谢和糖代谢。
[0165]
综上所述,本申请对花生和玉米间作的花生果实膨大期功能叶的代谢路径进行比较转录组测序分析研究,并通过qrt
‑
pcr对分析结果进行验证,最终发现在花生荚果膨大期,响应间作玉米胁迫的167个差异表达基因尤其是显著富集在13个go条目的28个差异表达基因及显著富集在32条kegg代谢通路上的23个差异表达基因影响花生功能叶的形态和生理特性,主要涉及叶绿素代谢、碳氮代谢、脂质代谢和氨基酸代谢等生物途径及其关键酶,这些差异基因组成的的代谢调控网络积极参与了间作玉米胁迫下花生荚果膨大的发育进程,另外,核糖磷酸
‑3‑
差向异构酶和甘油
‑3‑
磷酸
‑2‑
o
‑
酰基转移酶也是花生应答间作玉米胁迫的重要调控酶,并与花生种仁中的物质积累和荚果膨大密切相关;上述结论为后期后人利用定向化学调控技术以减少间作玉米胁迫对花生产量造成的不利影响提供理论依据。
[0166]
以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对本发明范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明的保护范围应以所附权利要求为准。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1