一种调控杨树木质部发育的PagARR9基因及其应用的制作方法
一种调控杨树木质部发育的pagarr9基因及其应用
技术领域
1.本发明涉及一种调控杨树木质部发育的pagarr9基因及其应用,特别涉及一种调控杨树木质部发育的细胞分裂素响应调节因子pagarr9基因及其应用,属于植物基因工程技术领域。
背景技术:
2.现代林业担负着维护国土生态安全,满足木材需求等林产品战略物质供应的重要使命。随着经济的高速发展,我国对森林资源及林产品,特别是木材的消费需求量,持续大幅增长,现已成为世界第二大木材进口国。通过培育速生优质人工林新品种,提高有限造林地的木材产量和质量显得尤为重要。但是由于林木生长周期长、杂合度高等诸多自身生物学特性的限制,迄今对木材材性的遗传背景知之甚少,制约了人工林优质、高产目标的实现。
3.因此,揭示木材形成的遗传调控机理,不但可以丰富树木分子生物学理论,而且可以为采用分子育种技术对树木的材性改良提供支持。
4.林木形成层活动及次生维管组织发育受到多种外界环境因子及内源激素的协同作用,其中细胞分裂素对维管形成层的形成及分裂分化具有重要影响。通过调控细胞分裂素合成或代谢关键基因,增加或降低杨树形成层区域细胞分裂素浓度,显著增加或降低了形成层细胞活性和木材产量,但其调控形成层活动和木质部发育的分子机理尚不清楚。
5.细胞分裂素响应调节因子rrs是细胞分裂素信号转导过程中的关键因子,它们是细胞分裂素相关的调节基因中家族数目最多的,通过转录活性和蛋白活性的调节调控下游信号。
6.在拟南芥基因组中,编码所有的正向和负向细胞分裂素响应调节因子的arr基因家族一共24个,不同成员之间存在功能冗余,共同参与对植物生长发育的调控。其中,a类arr家族蛋白主要是作为负反馈调节细胞分裂素信号,b类arr是一类植物特异转录因子,细胞分裂素信号途径依赖于主要的b类arr的稳定性。
7.杨树作为木本模式植物,与拟南芥不同,顶端分生组织具有自我更新和持续分生能力,既可分化为顶芽又能形成花芽,维持多年生的特性,具有独特的分生组织发育调控机制。植物在长期进化过程中,许多重要调控因子在多年生木本植物中出现了同源基因家族数目的变化,基因功能可能也发生了分化。杨树中共发现33个rrs家族基因,不同成员之间有不同的表达模式,在不同的生物学过程中行使不同的功能。
8.因此,利用模式木本植物杨树,研究细胞分裂素响应调节因子rr调控木材形成的分子机制,筛选调控形成层活动和木材形成的特异arr基因,对了解细胞分裂素调控形成层活动和木材产量以及杨树基因工程育种具有重要意义。
技术实现要素:
9.本发明的目的在于提供一种能调控杨树木质部发育的基因pagarr9,并利用其表
达载体转化杨树,以期促进林木分子育种技术的发展,为优良树种的培育或筛选提供技术手段,同时为探索木质部发育的分子机理奠定基础。
10.为解决上述技术问题,本发明采用如下技术方案:
11.本发明基于对杨树细胞分裂素rrs家族成员的分析研究,鉴定出一个可以调控杨树木质部发育优势表达基因pagarr9,其编码区核苷酸序列和氨基酸序列分别如seq id no.3和seq id no.4所示。pagarr9 cds全长600bp,编码199个氨基酸和1个终止密码子。
12.在强启动子35s后插入木质部优势表达基因pagarr9,得到过表达载体,转化杨树,获得过表达植株。
13.利用保守序列设计sgrna引物,构建pagarr9基因的crispr/cas9载体,转化杨树,获得pagarr9敲除植株。
14.在具体研究过程中,利用上述载体通过农杆菌介导法转化银腺杨84k(populus alba
×
p.glandulosa),分别获得了pagarr9过表达株系(oe-32、oe-37、oe-38)和基因敲除植株(c9-2、c9-7、c9-21)。
15.对上述所得不同转基因株系进行表型及显微形态观察发现,pagarr9过表达株系显著变矮,木质部显著减少;而pagarr9敲除株系则显著增高,木质部显著增加。
16.以上研究结果证明,pagarr9对杨树株高及木质部发育具有关键调控作用,其在林木分子育种及优良品种选育中具有重要应用价值。
17.与现有技术相比,本发明的主要有益技术效果在于:
18.本发明以银腺杨84k为材料,筛选鉴定出了pagarr9基因,基于其过表达和敲除植株的表型鉴定表明,其能够调控杨树木质部发育,说明pagarr9基因是调控杨树木质部发育的关键调节因子,为木质部发育的调控手段提供了新的选择,在林木基因工程领域有重要应用价值。
19.下面通过具体实施方式和附图对本发明作进一步说明,但并不意味着对本发明保护范围的限制。
附图说明
20.图1为本发明实施例1中杨树细胞分裂素响应调节因子pagarr9基因的组织表达特性分析图。
21.图2为本发明实施例1中野生型84k杨树与过量表达pagarr9的转基因杨树的转录水平定量检测图。
22.图3-1为本发明实施例1中c9-2敲除转基因植株检测结果。
23.图3-2为本发明实施例1中c9-7敲除转基因植株检测结果。
24.图3-3为本发明实施例1中c9-21敲除转基因植株检测结果。
25.图4为本发明实施例1中野生型84k杨树与过量表达pagarr9和基因敲除的转基因杨树株高表型比较图。
26.图5-1为本发明实施例1中野生型84k杨树与过量表达pagarr9和基因敲除的转基因杨树地茎统计。
27.图5-2为本发明实施例1中野生型84k杨树与过量表达pagarr9和基因敲除的转基因杨树株高统计。
28.图6为本发明实施例1中野生型84k杨树与过量表达pagarr9和基因敲除的转基因杨树木质部组织切片观察。
29.图7为本发明实施例1中野生型84k杨树与过量表达pagarr9和基因敲除的转基因杨树木质宽度统计。
具体实施方式
30.下面结合具体实施例对本发明做进一步的说明,以下实施例中未进行详细说明的操作可参照分子克隆,相关试剂盒使用说明的操作实现。
31.除非特别说明,下面实施例中所涉及的试剂均为市场上可购的常规试剂,所用的方法均为本技术领域常用的方法。
32.实施例1:
33.一、杨树pagarr9基因组织表达分析
34.1.杨树pagarr9基因的克隆
35.以银腺杨84k(p.alba x p.glandulosa)为材料,使用rneasy plant mini试剂盒和rnase-free dnase i试剂盒(qiagen,hilden,germany)提取84k杨树总rna;每个样品取2.0μg rna,通过使用superscript iii first-strand synthesis system(life technologies,carlsbad,ca,usa)合成cdna第一链,参考已发表毛果杨基因组序列,使用primer3软件设计引物(扩增子包含起始密码子及终止密码子),进行基因全长扩增(引物中引入gateway接头)。
36.其中,pagarr9 orf正向引物pagarr9-cds-f如序列表中seq id no.1所示(表1),反向引物pagarr9-cds-r如seq id no.2(表2)所示。
37.表1
[0038][0039]
表2
[0040][0041]
pcr反应体系如下:takara高保真扩增酶混合液primestar 12.5μl,正向引物(10μm)1μl,反向引物(10μm)1μl,模板(84k杨cdna)1μl,无菌ddh2o补足至25μl;反应程序:预变性98℃,5min;98℃,30s;56℃,30s;72℃,3min,10个循环;98℃,30s;60℃,30s;72℃,3min,25个循环;72℃10min;最终获得基因全长cdna序列为600bp,命名为pagarr9基因,序列如序列表seq id no.3所示(表3),其所编译表达蛋白序列如seq id no.4所示(表4)。
[0042]
表3
[0043][0044]
表4
[0045][0046]
2.组织表达特性分析
[0047]
植物材料来自温室生长2个月的84k杨野生植株的各个组织和器官,包括以下几个部分,顶端分生组织(sam):包含植物顶端未展开的第一和第二片叶子,还包括叶子中间的茎段;叶片(leaf):从第3片叶子开始到第7片叶子为止的连续叶片;皮(bark):从第15节间到植株基部的树皮,这部分组织主要涵盖了形成层区域和韧皮部区域;木质部(dix):从第15节间到植株基部的茎段,去掉皮之后,刮取的正在发育的木质部区域;茎段(stem):第2-3节间、第4、6、8、10、15、20节间的所有组织,包括了皮、形成层区域和木质部区域等。
[0048]
上述每一个样品都包含三个生物学重复,所有的植物材料均用液氮速冻后保存于-80℃冰箱中备用,提取样品rna,反转录合成cdna,以杨树actin作为内参基因,采用定量pcr分析pagarr9基因的组织表达特性,扩增引物为pagarr9-rt-f(seq id no.5,表5)和pagarr9-rt-r(seq id no.6,表6)。
[0049]
表5
[0050]
名称序列5(seq id no.5)pagarr9-rt-fgtggaaatggctctggctac
[0051]
表6
[0052]
名称序列6(seq id no.6)pagarr9-rt-rtcaagcccaaaaactccaag
[0053]
实时荧光定量pcr反应体系如下(10μl反应体系):
[0054][0055]
反应程序为:95℃预变性5min;然后进行45个循环的扩增,包含:95℃变性20sec,58℃退火20sec和72℃延伸20sec;最后72℃延伸5min;同时采取如下程序获得溶解曲线:从60℃开始升温到95℃的过程中,采取0.06℃/sec的速度升温(5acquisitions per℃),直至升温结束,完成整个程序并结束运行;采用2-δδct
计算方式进行构建荧光定量结果分析。
[0056]
如图1所示,为本发明实施例1中杨树细胞分裂素响应调节因子pagarr9基因的组织表达特性分析图;通过荧光定量pcr分析发现,pagarr9基因在不同组织中表达量存在差异,在根中有相对较高的表达丰度,而在茎中随着茎段的木质化程度增加,相对表达量增加,而且木质部的表达量明显高于韧皮部,因此,推测该基因在次生木质部形成调控方面可能发挥重要的作用。
[0057]
二、pagarr9基因植物表达载体构建
[0058]
1.过量表达载体的构建
[0059]
利用gateway技术,构建pagarr9基因的过量表达载体,使用特异pcr引物,以84k cdna为模板,进行pcr扩增,将pagarr9基因orf构建到入门载体pdnor222.1;反应体系为pcr产物80ng;pdnor222.1 vector 0.4μl;bp酶(货号:invitrogen 11789020)0.6μl;无菌ddh2o补足至5μl,反应程序为:25℃反应5h。
[0060]
从筛选培养板上挑取阳性克隆进行pcr检测及测序验证,带pagarr9基因的入门载体通过mlui限制性内切酶线性化后,与植物表达载体pmdc32进行lr反应,反应体系为:线性化已连接pagarr9基因的pdnor222.1载体50ng;pmdc32载体75ng;lr酶(货号:invitrogen 11791020)0.6μl;水补足至5μl,反应条件:25℃反应5h;经lr反应后,pagarr9基因导入植物表达载体pmdc32中,得到过量表达载体(pmdc32-pagarr9),在pagarr9基因的5’端组装有强表达启动子camv35s,它能使pagarr9基因在杨树体内高效表达;在pagarr9基因的3’端组装有强终止子nos,可有效终止pagarr9基因的转录。
[0061]
2.基因敲除载体的构建
[0062]
(1)引物设计:84k杨pagarr9基因组序列来源,根据毛果杨ptarr9的基因组序列设计全长基因组引物,以84k杨基因组dna为模板,克隆pagarr9基因组序列,选择在没有snp(single nucleotide polymorphism,单核苷酸多态性)位点的位置,选取一段20bp的随机核苷酸序列(pam)作为靶序列,合成目的引物序列atu3dt1f(seq id no.7)(表7)、atu3dt1r(seq id no.8)(表8)、atu6-1t1f(seq id no.9)(表9)、atu6-29t1r(seq id no.10)(表10)。
[0063]
表7
[0064]
名称序列7(seq id no.7)atu3dt1fgtcacagtggaattgtaatggctg
[0065]
表8
[0066]
名称序列8(seq id no.8)atu3dt1raaaccagccattacaattccactg
[0067]
表9
[0068]
名称序列9(seq id no.9)atu6-1t1fgtcattcaaacccaaaaactcca
[0069]
表10
[0070]
名称序列10(seq id no.10)atu6-1t1raaactggagtttttgggtttgaa
[0071]
(2)靶序列接头的制备:将靶序列引物用te buffer溶液或者双蒸水稀释成1μm母液,然后上下游靶序列引物各取10μl,混匀后,采取90℃变性30s,然后移至室温条件下,逐渐冷却退火,形成双链。
[0072]
(3)配制10μl体系的1
×
bsaⅰ的酶切连接反应液(10μl体系):
[0073][0074]
用pcr仪反应5个循环:37℃,5min;20℃,5min。
[0075]
(4)扩增grna表达盒,采用2轮pcr扩增,以获得更稳定的特异性目标产物以及避免空载产物扩增。
[0076]
1)第一轮pcr扩增,针对正向靶序列和反向互补靶序列,分别做两个pcr反应扩增,取1μl步聚(3)得到的酶切连接产物作为模板,使用表7、表8、表9、表10中的引物进行pcr反应,引物浓度均为10μm:
[0077][0078]
反应1和反应2的反应条件均为:95℃,15sec;60℃,15sec;72℃,15sec;循环数为25-28个循环,取5μl pcr产物,进行2%琼脂糖凝胶电泳检测;并各取反应1和反应2的pcr产物各1μl,用ddh2o稀释10倍后,从中吸取2μl,作为第二轮pcr反应的模板。
[0079]
表11
skoog(ms)基本培养基添加0.5mg/l 6-苄基氨基嘌呤(6-benzyl aminopurine)(6-ba)和0.05mg/l萘乙酸(naphthaleneacetic acid)(naa))上,在温度为22
±
2℃的黑暗条件下共培养3天,共培养后的叶片转移至含有3mg/l潮霉素(hygromycin b)和200mg/l特美汀(timentin)的sim上,在培养温度为23-25℃、光照为16/8h(白天/黑夜)、光照强度为50μm m-2s-1的条件下,诱导和筛选抗性不定芽,经过30天的诱导培养,将抗性不定芽转移至含有3mg/l潮霉素(hygromycin b)和200mg/l特美汀(timentin)的生根培养基中(rim,1/2ms基本培养基添加0.05mg/l iba和0.02mg/l naa),直至诱导生根,提取已生根植株叶片dna进行pcr验证。
[0095]
2.过表达转基因植株检测
[0096]
获得抗性的pagarr9基因过表达84k杨和野生型植株,提取基因组dna,利用pcr扩增表达载体上抗性基因,能够扩增得到清晰条带,即为转基因植株;选取转基因植株的茎段,以野生型茎段为对照,提取总rna并反转录,进行pagarr9基因定量分析,以确定转基因植株目的基因表达量,定量引物为pagarr9-rt-f(seq id no.5),pagarr9-rt-r(seq id no.6)。
[0097]
如图2所示,为本发明实施例1中野生型84k杨树与过量表达pagarr9的转基因杨树的转录水平定量检测图,其中,pagarr9基因在过量表达转基因株系oe-32、oe-37、oe-38中的表达量分别是野生型84k中的14.4、12.9、12.3倍。
[0098]
3.crispr-cas9介导的pagarr9基因编辑突变体植株鉴定
[0099]
设计突变体植株检测引物9-c9-f617(seq id no.14,表14)、9-c9-r617(seq id no.15,表15),目的条带包含pagarr9基因pam位置;提取转基因植株dna为模板,用高保真酶(宝日医,prime starmax dna聚合酶)进行pcr扩增,所得pcr产物连接至t载体(艾德莱零背景ptopo-ta/blunt通用克隆试剂盒,艾德莱,北京)并转化至大肠感菌感受态细胞,经菌液pcr鉴定为阳性的克隆,送测序公司测序,选择30个以上单克隆进行序列比对分析,测序结果如图3-1至图3-3所示,其中,如图3-1所示,为本发明实施例1中c9-2敲除转基因植株检测结果,由测序结果可知,该株系两条染色体上均有274个碱基(表16序列)被编码不同的275个碱基(表17序列)替换;如图3-2所示,为本发明实施例1中c9-7敲除转基因植株检测结果,由测序结果可知,该株系中两条染色体上均存在目标基因序列缺失1个碱基的编辑方式;如图3-3所示,为本发明实施例1中c9-21敲除转基因植株检测结果;由测序结果可知,该株系中两条染色体上均存在目标基因序列缺失2个碱基的编辑方式。
[0100]
表14
[0101]
名称序列14(seq id no.14)9-c9-f617agtccctcaccttcccttctttc
[0102]
表15
[0103]
名称序列15(seq id no.15)9-c9-r617gccacatagtcagatttctagcata
[0104]
表16
[0105][0106]
表17
[0107][0108][0109]
四、pagarr9转基因植株表型观察
[0110]
将pagarr9突变体植株和过表达植株以及野生型84k杨,作为对照,同时种植在温室中,共种三批,并分别对不同批次的植株进行表型测定和拍照,如图4所示,为本发明实施例1中野生型84k杨树与过量表达pagarr9和基因敲除的转基因杨树株高表型比较图,图4显示了移栽两个月后的表型照片,与非转基因84k杨植株相比,突变体植株整体较为高大粗壮,而过表达植株则表现出纤细弱小表型;如图5-1所示,为本发明实施例1中野生型84k杨树与过量表达pagarr9和基因敲除的转基因杨树地茎统计;如图5-2所法,为本发明实施例1中野生型84k杨树与过量表达pagarr9和基因敲除的转基因杨树株高统计;在株高上,突变体c9-21植株增高极显著,增加了24%,c9-7和c9-2植株增高显著,分别增高了12%和14%;过表达植株的高度降低也达到了极显著水平,oe-32、oe-37、oe-38高度分别降低了20%、22%、24%,结果如图5-1所示;在地径的粗度变化上,c9-21、c9-7、c9-2植株分别增粗了31%、27%和25%,达到了极显著水平,而过表达植株oe-32变细了12.5%,达到了极显著水平,oe-37和oe-38均变细了6%,达到显著水平,结果如图5-2所示。因此,综合表型和统计数据,初步推测pagarr9具有抑制茎段细胞分裂增殖和分化的功能。
[0111]
五、pagarr9转基因木质部切片观察
[0112]
1.取0.5cm长的土培苗杨树地面以上2cm处茎段,经loctite 495胶水固定在振荡切片机leica vt1200s切片槽内后,切成50μm厚度的横切面切片,切片保存于70%酒精中。
[0113]
2.组织化学染色及拍照观察分析:将新鲜切片进行0.05%tbo染色1min后,用水清洗三次,以清除浮色和多余的染色液,盖上盖玻片,采用olympus bx51普通光学显微镜,对
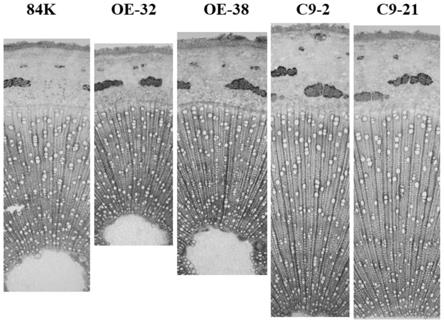
经tbo染色的切片进行观察,拍照,以分析茎段横截面的形态变化,结果如图6所示,为本发明实施例1中野生型84k杨树与过量表达pagarr9和基因敲除的转基因杨树木质部组织切片观察;与84k杨对照相比,pagarr9突变体植株木质部区域变宽和直径加粗现象;相反地,超量表达转基因植株表现出了木质部变窄和变细;统计c9-21和c9-2植株的木质部宽度,发现均分别增宽了45%和50%,而oe-32和oe-38植株,则减少了25%和12%,结果如图7所示,为本发明实施例1中野生型84k杨树与过量表达pagarr9和基因敲除的转基因杨树木质宽度统计。以上这些结果说明了pagarr9基因影响次生木质部的发育,即径向生长。
[0114]
对上述所得不同转基因株系进行表型及显微形态观察发现,pagarr9过表达株系株高显著变矮,木质部显著减少;而pagarr9敲除株系则株高显著增高,木质部显著增加。以上研究结果证明,pagarr9对杨树株高及木质部发育具有关键调控作用,其在林木分子育种及优良品种选育中具有重要应用价值。
[0115]
虽然以上对本发明目的构思和实施例作了详尽说明,但本领域普通技术人员可以认识到,在没有脱离权利要求限定范围的前提条件下,仍然可以对本发明作出各种改进和变换,而这种改进和变换仍然应当属于本发明的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1