一种用于调节VEGF基因表达的microRNA及其应用
stat3/dnmt1/ezh2蛋白复合物的结合促进了dnmt1与mir-374b-5p启动子的绑定是mir-374b-5p被mcp-1抑制的原因。这一系列过程为mcp-1刺激vegf基因表达的分子机制提供了关键联系。这些发现不仅有助于更好的理解mcp-1和vegf在肿瘤血管发生和肿瘤生长中的作用,同时还提供了一种潜在的治疗手段。通过外源性补充mir-374b-5p解除mcp-1对vegf的作用,抑制由mcp-1造成的肿瘤快速生长和转移。“阻断-解除阻断”机制关键在于vegf基因的大量表达可被mir-374b-5p的加入所阻断,而该阻断被mcp-1的作用解除。这种机制对于调节必需的多功能生长因子(如vegf)很有意义,因此mir-374b-5p在治疗vegf相关疾病中同样具有潜在的价值。从另一方面来看,在与冠状动脉疾病相关的缺血等情况下需要vegf的高活性来促进血管生成时,mir-374b-5p也可以作为治疗靶点,以缓解其对vegf基因表达的阻碍,从而达到促进血管新生改善缺血状态等。
附图说明
13.图1为mcp-1对小鼠体内血管生成的影响。
14.图2为mcp-1对小鼠体内基质胶中vegf mrna和蛋白表达的影响。其中a为vegf的mrna表达水平;b为vegf的蛋白表达水平。
15.图3为mcp-1对小鼠乳腺肿瘤体积的影响。
16.图4为mcp-1对小鼠乳腺肿瘤内部血管生成的影响。
17.图5为mcp-1对小鼠乳腺肿瘤内部vegf表达的影响。其中a为vegf的mrna表达水平;b为vegf的蛋白表达水平。
18.图6为mcp-1对小鼠体内基质胶及乳腺肿瘤中mir-374b-5p表达的影响。其中a为在基质胶中;b为在乳腺肿瘤内部。
19.图7为在huvec细胞中,加入mir-374b-5p对vegf表达的影响。其中a为vegf的mrna表达水平;b为vegf的蛋白表达水平。
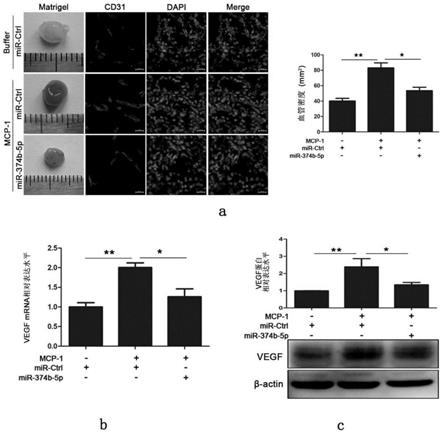
20.图8为在小鼠体内基质胶内注射mir-374b-5p对mcp-1诱导的血管新生及vegf的影响。其中a为基质胶血管生成情况;b、c分别为基质胶中vegf的mrna和蛋白表达情况。
21.图9为在小鼠皮下乳腺肿瘤中注射mir-374b-5p后对mcp-1诱导的肿瘤生长的影响。其中a为肿瘤照片及生长体积变化、小鼠体重变化情况;b为肿瘤内部血管生成情况。
22.图10为在小鼠皮下乳腺肿瘤中注射mir-374b-5p对瘤内vegf表达的影响。其中a、b分别为vegf的mrna和蛋白表达变化。
23.图11为mir-374b-5p启动子重组质粒设计示意图。
24.图12为mir-374b-5p启动子重组质粒荧光素酶活性测定。
25.图13为在huvec细胞中,5-aza对mir-374b-5p表达的影响。
26.图14为在huvec细胞中mcp-1增加dnmt1与mir-374b-5p启动子的结合。
27.图15为在小鼠体内基质胶和乳腺肿瘤中,mcp-1对stat3相关蛋白表达的影响。其中a为在基质胶中;b为在乳腺肿瘤内部。
28.图16为在huvec细胞中,pf-cbp1对mir-374b-5p表达的影响。
29.图17为在huvec细胞中,通过免疫共沉淀分析ac-stat3/dnmt1/ezh2相互作用情况。其中a为mcp-1对复合物结合的促进作用;b为pf-cbp1对mcp-1所致复合物结合增加的抑制作用。
具体实施方式
30.下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明的范围。
31.实施例1细胞培养及刺激
32.huvec细胞和bend.3细胞分别用含10%胎牛血清、1%亲链霉素(双抗)的1640或dmem培养基,在37℃5%co2培养箱中培养;取生长状态良好的对数期细胞进行实验。mcp-1(peprotech)100ng/ml刺激细胞48h建立体外模型;10μm pf-cbp1盐酸盐(medchemexpress)、4μm 5-aza-2
’‑
deoxycytidine(jena bioscience)预处理细胞2h后再与mcp-1共刺激48h。
33.实施例2体内血管生成实验
34.将bend 3.细胞消化计数,每5
×
105细胞中加入100μl基质胶,并混入相应的试剂(mcp-1 30ng,mir-ctrl/mir-374b-5p 100nμ)4℃轻摇4h使细胞与基质胶溶液彻底混匀;将100μl混悬液注射到雌性c57bl/6j小鼠腹部皮下,第3、5天胶内补充试剂,在第10天对小鼠实施安乐死后取出基质胶拍照并进行后续实验。
35.实施例3小鼠乳腺癌4t1细胞同种异体移植瘤模型
36.将2
×
106个4t1细胞注入雌性balb/c小鼠背部皮下,当肿瘤达到约100mm3时随机分组,记为第0天,每3天瘤内注射100μl对应试剂(mcp-1300n,mir-ctrl/mir-374b-5p 100nm);在第15天对小鼠实施安乐死后取出肿瘤拍照并进行后分析。
37.实施例4双荧光素酶报告基因检测
38.用构建的mir-374b-5p启动子重组质粒转染hek-293t细胞,24h后用双荧光素酶报告基因检测试剂盒(翊圣生物)检测细胞荧光值来评价mir-374b-5p启动子的主要区域。
39.实施例5rna提取和qpcr检测
40.基因总rna和microrna分别用trizol试剂(invitroge)及mirneasy mini kit(qiagen)提取;使用first-strand cdna synthesis supermix kit和top green qpcr supermix kit(全式金)进行反转录和扩增。引物序列为:h-vegf,5
’‑
cgcaagaaat cccggtataa and 5
’‑
aaatgctttc tccgctctga;h-β-actin,5
’‑
gcgagagatg acccagatc and 5
’‑
ccagtggtac ggccagagg。mirna引物购自广州瑞博生物有限公司。用2-δδct法分析相关mrna的相对表达量。
41.实施例6染色质免疫沉淀测定(chip)
42.采用chip检测试剂盒(碧云天)进行实验。细胞先后进行37%甲醛交联、细胞裂解液裂解、超声裂解后在上清中加入对应抗体4℃孵育过夜、加入protein g磁珠沉淀抗体-dna复合物、洗脱、解交联、dna提取与纯化;使用pcr试剂盒对dna片段进行扩增分析;mir-374b-5p启动子引物序列为5'-ggtcatttca ccttttattt ctctg和5'-agaggaagaa actgaagttc aacaa。得到的产物进行琼脂糖凝胶电泳。
43.实施例7免疫共沉淀(co-ip)与蛋白印迹
44.用np-40裂解缓冲液(碧云天)在冰上裂解细胞,离心留取上清液备用;在50μl蛋白a/g磁珠(bimake)中加入200μl稀释的抗体(ezh2、dnmt1、igg)溶液,室温混合翻转15min后用磁力吸附弃上清液;将裂解液加入与抗体结合的磁珠中,室温翻转1h形成蛋白-抗体-磁复合物;用1
×
上样缓冲液煮沸洗脱蛋白、弃磁珠;sds-page免疫印迹分析样品。
45.实施例1-7的试验结果讨论如下:
46.1.mcp-1促进vegf表达和肿瘤血管生成
47.本发明通过动物模型评估mcp-1对vegf所致的肿瘤血管生成的作用。在小鼠血管生成模型(基质胶实验)中发现,注射mcp-1的基质胶中bend.3细胞形成的血管数量约为对照组的2倍(图1),且vegf的mrna和蛋白表达水平也比对照组明显提高(图2)。在balb/c小鼠皮下种植小鼠乳腺癌4t1细胞所形成的肿瘤模型中我们发现,与对照组相比,瘤内注射mcp-1会导致肿瘤体积增大约2倍(图3),且肿瘤切片后内皮细胞标记物cd31免疫荧光染色显示,mcp-1瘤内注射组的肿瘤内部血管密度是对照组肿瘤的2倍以上(图4),并伴随着瘤内vegf表达的增加(图5)。这些数据表明mcp-1促进vegf表达以及肿瘤血管新生。
48.2.mir-374b-5p抑制mcp-1所致vegf基因的高表达
49.与对照组相比,在mcp-1刺激的基质胶及乳腺肿瘤中mir-374b-5p表达量明显减低(图6)。当在huvec细胞中加入mir-374b-5p时,发现mir-374b-5p组细胞内vegf的mrna和蛋白质表达较mir-ctrl组明显减少;不仅如此,mir-374b-5p的加入还能阻止mcp-1造成的vegf的高表达(图7)。这些发现提示mir-374b-5p可能在mcp-1调控vegf表达中发挥关键作用,通过加入外源性mir-374b-5p可抑制vegf的表达。
50.3.mir-374b-5p抑制mcp-1所致的血管新生和肿瘤生长
51.在小鼠基质胶内注射mir-374b-5p会使mcp-1刺激下胶内血管生成减少约60%(图8a),并且mir-374b-5p的加入会抑制基质胶中mcp-1刺激下vegf的大量产生(图8b,c)。同样,mir-374b-5p瘤内注射会明显抑制肿瘤的生长,且不影响小鼠体重,说明对小鼠无毒性;并会使mcp-1刺激下的平均肿瘤体积减小约一半(图9a);极大地抑制了肿瘤内部血管的生成密度(图9b)。瘤内vegf的表达量也因为外源性mir-374b-5p的加入而有所减少(图10)。这些发现表明mir-374b-5p的加入能够显着抑制肿瘤内部血管生成及肿瘤生长,同时阻碍mcp-1对血管生成和肿瘤生长的促进作用。
52.4.mcp-1增加dnmt1与mir-374b-5p启动子的结合由于vegf基因的表达可以被加入的mir-374b-5p阻断,说明在mcp-1诱导vegf表达时需要阻断mir-374b-5p的作用。发明人根据mir-374b-5p启动子区域序列(如seq id no:1所示)构建了多个重组质粒(图11),即将人基因组中mir-374b转录起始位点上游2000bp序列(chrx:74217547-74219546),进行划分,选取:promoter1(p1):chrx:74218937-74219546(610bp);promoter2(p2):chrx:74218937-74219228,(292bp);promoter3(p3):chrx:74219229-74219546(318bp),将其分别插入pgl3-basic-vector质粒中,共得到三个重组质粒。通过测定重组质粒的荧光素酶活性发明人找到mir-374b-5p启动子功能区主要集中在chrx:74218937-74219228(p2,图12)。当使用dna甲基转移酶抑制剂5-aza-2'-脱氧胞苷(5-aza)时,5-aza会阻碍huvec细胞中mcp-1对mir-374b-5p表达的下调(图13)。由于dnmt1是哺乳动物基因组表观遗传修饰中dna甲基化的关键基因,发明人接下来探究dnmt1是否与mir-374b-5p表达量的变化相关。通过染色质共沉淀(chip)分析发现dnmt1能与mir-374b-5p启动子结合,且mcp-1可以促进了二者的结合(图14)。综上所述,dnmt1参与mcp-1对mir-374b-5p表达的调控,通过增加与mir-374b-5p启动子的结合阻碍mir-374b-5p的表达。
53.5.mcp-1通过增加ac-stat3/dnmt1/ezh2复合物的形成促进dnmt1与mir-374b-5p启动子的结合
54.已知乙酰化stat3(ac-stat3)和dnmt1的复合物可协同甲基化许多肿瘤抑制基因,同时dnmt1/ezh2可以共同催化mirna甲基化。通过检测体内小鼠基质胶及乳腺肿瘤中stat3的变化发现,mcp-1刺激并不影响小鼠基质胶中及瘤内stat3磷酸化(活性形式)和stat3总量的变化,而会增加stat3在lys685位点处的乙酰化水平(图15)。我们进一步发现加入pf-cbp1(stat3乙酰化形式的特异性抑制剂)可有效地解除mcp-1对mir-374b-5p表达的抑制作用(图16)。此外,免疫共沉淀分析显示mcp-1促进huvec细胞中的ac-stat3/ezh2/dnmt1复合物的相互作用,而pf-cbp1可抑制这些相互作用(图17)。这些发现表明mcp-1通过增加stat3在lys685位点处的乙酰化水平促进ac-stat3/ezh2/dnmt1复合物的形成,从而导致dnmt1与mir-374b-5p启动子结合增多,最终抑制mir-374b-5p的表达。
55.以上所述仅为本发明的较佳实施例而已,并不用以限制本发明,凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1