一种检测腐霉利的方法及试剂盒
1.本发明涉及杀菌剂的检测技术领域,具体地,涉及一种检测腐霉利的方法及试剂盒。
背景技术:
2.腐霉利(procymidone,prm)是一种低毒、廉价、高效、广谱的二甲酰亚胺类内吸 性杀菌剂,作用机理是抑制菌体内甘油三酯的合成,对葡萄孢属和核盘菌属真菌具有特效 作用,主要用于防治果树、蔬菜、花卉的灰霉病、菌核病以及其他作物或食物如三七、人 参等的黑斑病。虽然腐霉利毒性不高,但是其在果蔬中的残留可以通过食物链进入动物或 人体,其代谢物二氯苯胺会对机体产生一定的毒性,严重时可导致哺乳动物性器官畸形、 萎缩和病变等。近年来,由于腐霉利在果蔬上的广泛使用,使其成为农药残留检出率和超 标率较高的农药品种之一,尤其是韭菜中腐霉利超标事件在各地监管抽检中频频曝光,因 此加强对腐霉利农药残留的检测十分必要。
3.目前,对腐霉利残留的检测方法多为气相色谱法、气质联用法和液相色谱法。尽管这 些仪器方法具有实验准确可靠、灵敏度高和重复性好等优点,但是存在样品前处理过程复 杂、检测时间长、仪器昂贵等不足,只能用于小批量样品抽检,无法满足对大批量样品进 行现场、快速检测的需要。
4.基于抗原和抗体的特异性反应的酶联免疫检测方法具有高通量、高灵敏度、快速、易 操作、低成本等优势,可以制成商品化的试剂盒以便于现场检测,已在农兽药残留及微量 毒素检测中被广泛应用。免疫分析方法成功的关键在于制备灵敏度高特异性的抗体,目前 应用最为广泛的是单克隆抗体与多克隆抗体,但是这些传统抗体具有开发周期长、稳定性 较低、保存条件苛刻等缺陷,制备得到一种对腐霉利具有强特异性、高灵敏度的稳定抗体, 对建立快速、灵敏、准确的腐霉利检测方法具有重要意义。
5.纳米抗体(variable domain of heavy chain of heavy chain antibody,vhh)是一种新型 的基因工程抗体,来源于骆驼及鲨鱼类动物体内重链抗体的可变区,具有低免疫原性、高 稳定性、高亲和力和高特异性等特点。与常规抗体相比,纳米抗体更易于储藏和运输,甚 至能够在高温、化学和压力等极端条件下变性后仍可有效地重折叠并恢复其抗原亲和力。 此外,纳米抗体可以通过原核生物进行大量表达,制备周期短,可有效地避免传统抗体的 批间差异问题,具有更高的经济效益与应用优势。
技术实现要素:
6.本发明要解决的技术问题是克服现有检测腐霉利的方法的缺陷和不足,提供一种腐霉 利纳米抗体及其制备方法和应用。
7.本发明的第一个目的是提供一种抗腐霉利的纳米抗体。
8.本发明的第二个目的是提供一种编码所述纳米抗体的基因。
9.本发明的第三个目的是提供一种重组载体,此重组载体含有本发明第二个目的所
述基 因的核苷酸序列。
10.本发明的第四个目的是提供一种重组细胞,所述重组细胞转染有本发明第三个目的所 述表达载体。
11.本发明的第五个目的是提供所述纳米抗体、所述基因、所述重组载体、或所述重组细 胞中的一种或几种在检测腐霉利的免疫学检测分析中的应用。
12.本发明的第六个目的是提供一种腐霉利的检测方法。
13.本发明的第七个目的是提供一种检测腐霉利的试剂盒。
14.为了实现上述目的,本发明是通过以下方案予以实现的:
15.本发明通过构建双峰驼免疫抗体文库,用噬菌体展示技术,将检测抗原固相包被于酶 标板上,投入双峰驼免疫抗体文库进行亲和淘筛,获得一种抗腐霉利特异性结合的纳米抗 体,其氨基酸序列如seq id no.1所示。并应用于腐霉利的免疫学检测分析,通过此免疫 检验分析建立了一种快速、灵敏并且稳定的腐霉利的检测方法。
16.因此本发明要求保护以下内容:
17.一种特异性识别腐霉利的纳米抗体,其氨基酸序列如seq id no.1所示。
18.其中,所述纳米抗体氨基酸序列的框架区(fr1-fr4)依次为如seq id no.2,seq idno.3,seq id no.4,seq id no.5所示;互补决定区(cdr1-cdr3)依次如seq id no.6, seq id no.7,seq id no.8所示。
19.一种编码特异性识别腐霉利纳米抗体的基因,其核苷酸序列如seq id no.9所示。
20.具体地:
21.所述纳米抗体的vhh的氨基酸序列如seq id no.1所示:
22.evqlldsgggsvqaggslrlscevftskiatgymgwfrqapgkereavaithtgsg npwyadsvkgrftisqdnakltvylqmnnlrpedtatytcaareagswelrrlspewy tywgqgtlvtvss;
23.所述纳米抗体的框架区fr1的氨基酸如seq id no.2所示:
24.evqlldsgggsvqaggslrlscevf;
25.所述纳米抗体的框架区fr2的氨基酸如seq id no.3所示:
26.mgwfrqapgkereavai;
27.所述纳米抗体的框架区fr3的氨基酸如seq id no.4所示:
28.wyadsvkgrftisqdnakltvylqmnnlrpedtatytc;
29.所述纳米抗体的框架区fr4的氨基酸如seq id no.5所示:
30.wgqgtlvtvss;
31.所述纳米抗体的互补决定区cdr1的氨基酸如seq id no.6所示:
32.tskiatgy;
33.所述纳米抗体的互补决定区cdr2的氨基酸如seq id no.7所示:
34.thtgsgnp;
35.所述纳米抗体的互补决定区cdr3的氨基酸如seq id no.8所示:
36.aareagswelrrlspewyty;
37.所述编码特异性纳米抗体的基因的核苷酸序列如seq id no.9所示:
38.gaggtgcagctgctggattctgggggaggctcggtacaggctggagggtctct gagactctcgtgtgaagtttttacaagcaagatagcaaccggctacatgggctggtt ccggcaggctccgggtaaggagcgcgaggccg
tcgcaattactcataccggtagtgg aaacccatggtatgccgactccgtgaagggccgcttcaccatctcccaagacaacgc caagctcacggtgtatctgcaaatgaacaacctgagacctgaggacactgccacgt acacctgtgcggcccgggaggcagggtcttgggagctcaggcggttatctccagaa tggtatacctactggggccaggggaccctggtcaccgtctcctca。
39.一种重组载体,连接有所述编码基因。
40.一种重组细胞,含有所述表达载体,或能够表达所述纳米抗体。
41.所述纳米抗体、所述基因、所述重组载体和/或所述重组细胞中的一种或几种在检测 腐霉利和/或制备腐霉利免疫学检测试剂盒中的应用,也属于本发明的保护范围。
42.本发明要求保护一种腐霉利的非诊断目的检测方法,利用所述纳米抗体。
43.优选地,基于间接竞争elisa方法进行检测,用式(ⅰ)所述的腐霉利半抗原与载 体蛋白偶联得到的腐霉利完全抗原作为检测抗原,用所述纳米抗体作为检测抗体进行检 测,
[0044][0045]
优选地,所述载体蛋白为载体蛋白为钥孔血蓝蛋白(klh)、牛血清蛋白(bsa)或 鸡卵白蛋白(ova)。
[0046]
更优选地,所述载体蛋白为牛血清蛋白(bsa),所述检测抗原结构式如(ⅲ)所示,
[0047][0048]
更优选地,包被有检测抗原的固相载体中先后加入待测样品与氨基酸序列如seq idno.1所示的纳米抗体,充分反应后弃去液体并洗涤;加入酶标记抗二体充分反应后弃去 液体并洗涤,充分反应后弃去液体并洗涤;进行显色反应,终止反应;读450nm od值。
[0049]
在一个具体的实施例中,显色液a与显色液b混合的tmb显色液进行显色反应,10% h2so4(v/v)终止反应。
[0050]
本发明还要求保护一种检测腐霉利的试剂盒,含有所述纳米抗体。
[0051]
优选地,还包括式(ⅰ)所述腐霉利半抗原与载体蛋白偶联得到的腐霉利完全抗做检 测抗原,用所述纳米抗体作为检测抗体,
[0052][0053]
更优选地,所述载体蛋白为牛血清蛋白(bsa),所述检测抗原结构式如(ⅲ)所示,
[0054][0055]
优选地,检测抗原包被于固相载体。
[0056]
优选地,还包括酶标二抗、显色剂和终止剂。
[0057]
更优选地,显色剂为显色液a与显色液b。
[0058]
更优选地,终止剂为10%h2so4(v/v)。
[0059]
更优选地,酶标二抗为anti-vhh-hrp二抗。
[0060]
与现有技术相比,本发明具有以下有益效果:
[0061]
本发明制备得到的纳米抗体可通过基因工程重组表达的方式进行大量制备。基因工程 重组表达的方式是将编码该纳米抗体的基因克隆至表达载体,以蛋白表达的形式进行该纳 米抗体的大量制备。所述纳米抗体经过原核生物表达后,以蛋白的形式进行免疫学检测分 析。
[0062]
本发明制备得到的纳米抗体可以应用于腐霉利残留的实际样品检测。该纳米抗体具有 灵敏度高、特异性强、耐高温、耐酸碱、易保存的优点,并且具有生产成本低、生产周期 短等优势,可作为检测腐霉利的新型材料。本发明制备纳米抗体的方法具有普遍适用性, 可用于其他小分子物质纳米抗体的筛选和制备,具有较高的应用价值。
附图说明
[0063]
图1为腐霉利半抗原、载体蛋白与完全抗原的紫外扫描光谱图。
[0064]
图2为腐霉利阳性克隆的elisa鉴定示意图。
[0065]
图3为腐霉利纳米抗体nbfm5的sds-page图。
[0066]
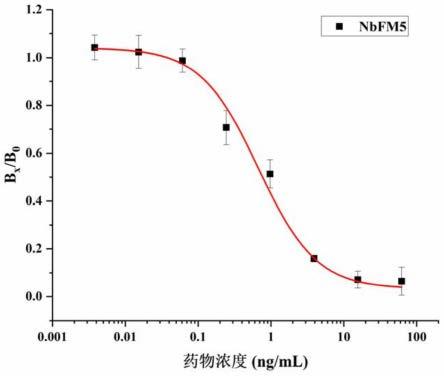
图4为基于纳米抗体nbfm5的elisa抗原抗体工作浓度的优化。
[0067]
图5为基于纳米抗体nbfm5建立的间接竞争elisa标准曲线。
[0068]
图6为纳米抗体nbfm5在不同浓度的甲醇/乙腈/丙酮下的活性示意图。
[0069]
图7为纳米抗体nbfm5在70℃条件下孵育不同时间后抗体的活性示意图。
[0070]
图8为纳米抗体nbfm5在不同ph的pbs中的活性示意图。
[0071]
图9为纳米抗体nbfm5在不同温度条件下储存不同时间的活性示意图。
具体实施方式
[0072]
下面结合说明书附图及具体实施例对本发明作出进一步地详细阐述,所述实施例只用 于解释本发明,并非用于限定本发明的范围。下述实施例中所使用的试验方法如无特殊说 明,均为常规方法;所使用的材料、试剂等,如无特殊说明,为可从商业途径得到的试剂 和材料。
[0073]
实施例1免疫抗原prm-lf和检测抗原prm-bsa的制备
[0074]
腐霉利半抗原的设计与合成由实验室前期工作完成,其结构式如(ⅰ)所示(该化合 物已经公开于,cn201910194037-一种腐霉利半抗原及其合成方法和应用):
[0075][0076]
一、实验方法
[0077]
1、完全抗原prm-lf的制备
[0078]
结构式如(ⅰ)所示的腐霉利半抗原与乳铁蛋白(lactoferrin,lf)通过活泼酯法偶 联形成免疫抗原,其结构式如(ⅱ)所示,
[0079][0080]
具体操作方法如下:称取2.876mg edc与1.725mg nhs,加入0.1ml dmf溶解固 体。将上述混合物溶液加入3.518mg腐霉利半抗原,室温避光搅拌或震荡反应4h活化半 抗原。称取20mg lf,加入2ml碳酸盐缓冲液(ph 9.4)配制成10mg/ml蛋白溶液, 冰浴冷却。搅拌冰浴中逐滴滴加活化后的腐霉利半抗原,并用ph试纸确定溶液在碱性范 围内,室温搅拌过夜。用pbs透析3次,分装后冻存于-20℃。用紫外分光光度法扫描偶 联原料(半抗原和载体
蛋白lf)和偶联产物,利用扫描得到的紫外吸收光谱判断偶联是 否成功。
[0081]
2、完全抗原prm-bsa的制备
[0082]
结构式如(ⅰ)所示的腐霉利半抗原与牛血清蛋白(bovine serμm albμmin,bsa)通 过活泼酯法偶联形成人工检测抗原,其结构式如(ⅲ)所示,
[0083][0084]
具体操作方法如下:称取6.535mg edc与3.92mg nhs,加入0.15ml dmf溶解固 体。将上述混合物溶液加入7.995mg腐霉利半抗原,室温避光搅拌或震荡反应4h活化半 抗原。称取50mg bsa加入5ml碳酸盐缓冲液(ph 9.4)配制成10mg/ml蛋白溶液, 冰浴冷却。搅拌冰浴中逐滴滴加活化后的半抗原,并用ph试纸确定溶液在碱性范围内, 室温搅拌过夜。用pbs透析3次,分装后在-20℃冻存。用pbs透析3次,分装后在-20℃ 冻存。用紫外分光光度法扫描偶联原料(半抗原和载体蛋白bsa)和偶联产物,利用扫描 得到的紫外吸收光谱判断偶联是否成功。
[0085]
二、实验结果
[0086]
腐霉利半抗原、载体蛋白(lf、bsa)和偶联产物的紫外扫描谱图见图1。腐霉利半 抗原的特征峰位于210nm以及280nm处,lf紫外吸收峰位于210nm以及280nm处,bsa 紫外吸收峰位于228nm以及280nm处。载体蛋白lf与腐霉利半抗原偶联后的完全抗原 prm-lfⅱ的紫外吸收峰为229nm、280nm,载体蛋白bsa与腐霉利半抗原偶联后的完 全抗原prm-bsa
ꢀⅲ
的紫外吸收峰为242nm、278nm。由此可见,两种人工抗原的紫外吸 收峰相对与半抗原和载体蛋白基本都发生了明显的位移且吸光度也发生了变化,表明腐霉 利半抗原与载体蛋白lf、bsa成功实现偶联。
[0087]
实施例2双峰驼免疫抗体文库的构建
[0088]
1、双峰驼免疫方案
[0089]
用健康的双峰驼进行动物免疫,以实施例1制备的结构式如(ⅱ)所示的完全抗原 prm-lf作为免疫抗原,在双峰驼背颈部进行皮下注射,每次免疫剂量为0.5mg免疫抗原。 首次免疫用0.5ml完全弗氏佐剂与免疫抗原混合乳化后用于免疫,后续的加强免疫用0.5 ml不完全弗氏佐剂与抗原乳化后免疫,每次免疫间隔2周,共进行4次加强免疫。在第 四、第五次免疫后,采集50~100ml外周血,用于构建纳米抗体文库。
[0090]
2、双峰驼淋巴细胞的分离
[0091]
采集双峰驼外周血需尽快进行淋巴细胞分离,具体操作方法如下:将双峰驼外周
血与 无菌生理盐水在无rna酶的干净容器中等体积混合稀释。稀释后的外周血用商品化淋巴 细胞分离液进行离心,不同密度的血细胞离心后分布淋巴细胞分离液不同深度,其中淋巴 细胞在液面下约1/3深度处,形成白色细胞层。在室温条件下,850g,离心25min。用无 rna酶的胶头滴管或移液器小心收集到灭酶的干净容器中,用无菌生理盐水洗涤细胞表 面多余的淋巴细胞分离液,离心收集细胞沉淀。收集的淋巴细胞中加入裂解液trnsol, 用无rna酶的移液器或胶头滴管反复吹打至淋巴细胞完全裂解于trnsol中,于-80℃保 存备用。
[0092]
3、总rna的提取
[0093]
采用商品化rna提取试剂盒或其他有效的rna提取方法,从上述保存于trnsol裂 解液中的淋巴细胞中提取总rna,根据rna提取试剂盒说明书进行操作即可。提取总 rna后取少许样品进行核酸电泳并在超微量分光光度计(nanodrop)中测定rna浓度。
[0094]
4、cdna的合成
[0095]
采用商品化反转录试剂盒将上一步所得的rna样品中的mrna反转录成cdna,根 据说明书进行操作即可。反转录完成后将产物先混合均匀,再分装于不同的无菌离心管, 保存在-80℃环境。
[0096]
5、纳米抗体目的基因vhh的扩增
[0097]
采用巢式pcr进行两步法扩增得到纳米抗体目的基因vhh,所用到的引物序列如表 1所示:
[0098]
表1双峰驼重链抗体基因引物序列
[0099][0100][0101]
第一轮pcr用cdna作为其pcr模板,具体反应参数如表2所示:
[0102]
表2巢式pcr第一步反应体系及反应条件
[0103][0104]
第一步pcr产物在核酸电泳后会出现1000bp以及750bp两种产物条带,对750bp 条
带进行切胶回收,并测定浓度。
[0105]
第二轮pcr用第一轮pcr回收产物作为模板进行第二轮pcr扩增,具体反应参数如 表3所示:
[0106]
表3巢式pcr第二步反应体系及反应条件
[0107][0108]
6、基因文库构建
[0109]
(1)目标基因vhh及载体pcomb3xss的酶切
[0110]
采用sfi i酶对vhh目标基因及pcomb3xss载体进行酶切反应。酶切条件:50℃恒温 反应16h。
[0111]
pcomb3xss载体酶切产物通过琼脂糖凝胶回收分子量为3500bp的条带;vhh基因酶 切产物通过dna回收试剂盒直接清洁回收。
[0112]
(2)酶切产物的连接
[0113]
将载体pcomb3xss和vhh片段混匀(摩尔比1:3),16℃反应16h后用dna回收 试剂盒清洁回收。
[0114]
(3)电击转化
[0115]
取5μl连接产物加入50μl电转感受态e.coil tg1中,轻轻混匀后转移到0.1cm的 电转杯中电击转化(电压为1.8kv),电击后立即向电转杯加入950μl预热到37℃的 soc培养基,37℃250rpm摇菌1h复苏细胞。
[0116]
取100μl复苏菌液进行梯度稀释,将各浓度梯度稀释菌液各取100μl涂布于直径90 mm的lb-amp培养皿作为计数板,37℃培养过夜。剩余未稀释的复苏菌液全部涂布于直 径120mm的lb-amp培养皿,每1ml菌液涂布于2~3个培养皿作为扩增板,37℃扩 增培养过夜。
[0117]
统计计数培养皿上的菌落数,计算复苏菌液中的细菌总数,进行多次电击转化,使转 化菌落总数累计达到107cfu以上,此数目即为纳米抗体基因文库的库容量。
[0118]
将扩增板中的转基因大肠杆菌菌落用细胞刮刀刮下,混合均匀后加入终浓度为25%的 甘油,取50μl菌液进行梯度稀释测定细胞数,其余菌液分装后于-80℃冻存,即为腐霉 利纳米抗体基因库。
[0119]
7、噬菌体救援
[0120]
根据上述转基因大肠杆菌细胞数测定结果接种10倍库容量以上的细胞于200mllb-amp中,37℃250rpm培养至对数期;加入1ml滴度为10
12
cfu/ml以上的辅助噬菌 体m13k07,37℃静置30min后,250rpm培养1h,加入卡那霉素(70μg/ml)37℃,250 rpm培养过
夜。12000rpm 4℃15min离心,取上清,加入1/4体积的peg/nacl溶液(100 g peg 8000与73.05g氯化钠加水定容至500ml),冰浴2~3h。12000rpm 4℃15min 离心,弃上清,沉淀用1ml tbs重悬,转移到2ml离心管,12000rpm 4℃5min离心, 过0.22μm聚醚砜滤膜。取10μl噬菌体测定滴度,其余加入终浓度50%甘油,-80℃保存, 即为腐霉利纳米抗体噬菌体库。
[0121]
实施例3纳米抗体的亲和淘选及其鉴定
[0122]
一、实验方法
[0123]
1、纳米抗体的亲和淘选
[0124]
(1)检测抗原固定化
[0125]
用包被液(0.375g na2co3与0.7325g nahco3加水定容至250ml)将实施例1制备 的结构式如(ⅲ)所示的完全抗原prm-bsa稀释至1μg/ml,加入酶标板微孔中,每孔 100μl,4℃静置过夜。第二天用稀释20倍后的洗液pbst(0.01m pbs,0.05%tween-20) 洗板两次后,每孔加入150μl 1%bsa-pbs(w/v)溶液37℃静置2h。倒出孔内液体, 在吸水纸上拍干并在37℃下烘1h后存于4℃备用。
[0126]
(2)阳性噬菌体筛选
[0127]
将上述腐霉利纳米抗体噬菌体文库中加入bsa,使bsa终浓度为1%(w/v),将噬 菌体加入3个有固定化抗原的微孔中,每孔加入100μl,37℃孵育1h。弃去孔中游离的 噬菌体,用pbst洗涤微孔10次,再用pbs洗涤微孔5次。
[0128]
在孔中加入浓度为5μg/ml的腐霉利标准品,每孔100μl,37
°
条件下震荡孵育4h进 行竞争反应,将微孔中的液体收集到无菌离心管中。此时的噬菌体称为“竞争output”。为 防止多样性的损失,在竞争洗脱结束的孔中加入胰蛋白酶trypsin(10mg/ml)洗脱30 min,将微孔中的液体收集到无菌离心管中,称为“酶洗output”,第一轮筛选完成。
[0129]
在筛选前后都要进行噬菌体滴度的测定,具体操作方法如下:
[0130]
取10μl两种output噬菌体分别进行10倍梯度稀释,选取预期滴度以及前后相邻的 两个稀释度(一般input稀释倍数为107、108、109,output稀释倍数为102、103、104), 取10μl稀释液侵染100μl对数期的e.coil tg1,37℃静置30min,涂布lb-agar-amp 平板,培养过夜后进行菌落计数并计算噬菌体滴度,计算公式如下:
[0131][0132]
(3)噬菌体扩增
[0133]
经过筛选的噬菌体扩增后用于下一轮筛选,噬菌体扩增的方法如下:
[0134]
将一半的竞争洗脱output噬菌体加入4ml对数期的e.coil tg1,静置30min。加入 lb培养基至总体积为10ml并加入amp至工作浓度(10μg/ml),37℃250rpm摇菌 30min,加入50μl滴度为10
12
cfu/ml的m13ko7辅助噬菌体,静置30min,37℃、250rpm 摇菌1h,加入lb至总体积为100ml,加入amp及kana,37℃250rpm摇菌过夜。
[0135]
将菌液离心,上清转移到新的离心瓶中,加入1/4体积的peg与nacl的混合溶液(100g peg 8000与73.05g氯化钠加水定容至500ml),混匀后冰浴2h,12000rpm离心20min, 将上清弃去并倒扣在吸水纸上尽量除去残余水分,用1ml含1%bsa(w/v)的tbs将白 色沉淀重悬,过0.22μm聚醚砜滤膜后于-20℃保存,作为下一轮筛选“input”,重复步骤(2) 的筛选方案。
[0136]
以上筛选步骤一共进行4轮,每轮检测抗原浓度固定为1ug/ml不变,第2、3、4轮 竞争反应所用药物浓度分别降到2000ng/ml,400ng/ml,40ng/ml。
[0137]
2、阳性克隆的鉴定
[0138]
采用间接酶联免疫吸附法进行阳性噬菌体克隆的鉴定。具体方法为:
[0139]
(1)抗原固定化
[0140]
检测抗原用包被液(0.375g na2co3与0.7325g nahco3加水定容至250ml)稀释至1 μg/ml,4℃静置过夜。第二天用稀释20倍后的洗液pbst(0.01m pbs,0.05%tween-20) 洗涤两次后,用pbs现配2%的脱脂奶粉(w/v),每孔加入150μl,37℃封闭2h,弃去 封闭液,拍干并37℃烘60min,存放于4℃待用。
[0141]
(2)纳米抗体小量表达
[0142]
第三、四轮淘筛的酶洗以及竞争洗脱的output滴度测定平板上随机挑选96个单菌落, 接种到每孔装有0.5ml lb-amp的96孔板中,同时接种一个未被噬菌体侵染的e.coil tg1 单菌落作为阴性对照,37℃培养过夜,作为菌液“母板”。
[0143]
从母板中每孔取出10μl菌液接种到另一块每孔装有1ml lb-amp的96孔深孔板中, 接种的孔编号要与母版对应,37℃,180rpm培养3h,每孔加入iptg使其最终工作浓度 为1mm,37℃180rpm培养过夜,母板4℃保存备用。
[0144]
(3)酶联免疫鉴定阳性克隆
[0145]
深孔板4000rpm离心20min,取两块有固定化抗原的酶标板,板1作为检测抗体的 效价板,每孔加入50μl pbs;板2作为抑制板,加入含有50ng/ml腐霉利标准药物的 pbs,并从离心后的96孔板中吸取上清,在对应编号的酶标孔中每孔加入50μl。37℃孵 育30min,用稀释20倍后的洗液pbst(0.01m pbs,0.05%tween-20)洗五次,拍干孔 内液体。用pbst将anti-vhh-hrp二抗(genscript,a01861-200)稀释5000倍,每孔加 入100μl,37℃孵育30min。用稀释20倍后的洗液pbst(0.01m pbs,0.05%tween-20) 洗五次,拍干孔内液体,每孔加入100μl预先用等体积显色液a与显色液b混合的tmb 显色液(solarbio,pr1210),37℃显色10min;加入50μl终止液10%h2so4(v/v)终 止反应;用酶标仪测定450nm处的吸收值。
[0146]
根据以下公式可计算出每个阳性克隆的抑制率。选取在板1中od值大于阴性对照孔 3倍,且具有明显抑制(抑制率>20%)的克隆,记录其对应孔的编号,并将母板中对应 孔的菌液转移到无菌离心管中,加入甘油冻存备用。
[0147][0148]
二、实验结果
[0149]
将经过间接竞争elisa鉴定获得的纳米抗体的菌株送到测序公司进行基因测序,根 据dna测序结果及密码子表可获得纳米抗体的氨基酸序列。结果显示获得了7株具有不 同序列的腐霉利纳米抗体,分别命名为nbfm1~nbfm7。这七株阳性克隆的鉴定结果如图 2所示,nbfm1~nbfm7的抑制率分别是78.91%、74.68%、88.61%、44.01%、93.57%、 38.02%和36.19%,其中nbfm5在间接竞争elisa检测中表现最为突出,抑制率最高。
[0150]
nbfm5具有seq id no.1所示的氨基酸序列,该纳米抗体包括4个fr框架区和3 个cdr互补决定区,其排列顺序为fr1、cdr1、fr2、cdr2、fr3、cdr3、fr4,其 中框架区(fr1-fr4)
依次如seq id no.2、seq id no.3、seq id no.4、se1%q id no.5 所示;互补决定区(cdr1-cdr3)依次如seq id no.6、seq id no.7、和seq id no.8 所示。
[0151]
同时,得到编码所述纳nbfm5的基因的核苷酸序列如seq id no.9所示。
[0152]
具体地:
[0153]
所述纳米抗体的vhh的氨基酸序列如seq id no.1所示:
[0154]
evqlldsgggsvqaggslrlscevftskiatgymgwfrqapgkereavaithtgsg npwyadsvkgrftisqdnakltvylqmnnlrpedtatytcaareagswelrrlspewy tywgqgtlvtvss;
[0155]
所述纳米抗体的框架区fr1的氨基酸如seq id no.2所示:
[0156]
evqlldsgggsvqaggslrlscevf;
[0157]
所述纳米抗体的框架区fr2的氨基酸如seq id no.3所示:
[0158]
mgwfrqapgkereavai;
[0159]
所述纳米抗体的框架区fr3的氨基酸如seq id no.4所示:
[0160]
wyadsvkgrftisqdnakltvylqmnnlrpedtatytc;
[0161]
所述纳米抗体的框架区fr4的氨基酸如seq id no.5所示:
[0162]
wgqgtlvtvss;
[0163]
所述纳米抗体的互补决定区cdr1的氨基酸如seq id no.6所示:
[0164]
tskiatgy;
[0165]
所述纳米抗体的互补决定区cdr2的氨基酸如seq id no.7所示:
[0166]
thtgsgnp;
[0167]
所述纳米抗体的互补决定区cdr3的氨基酸如seq id no.8所示:
[0168]
aareagswelrrlspewyty;
[0169]
所述编码特异性纳米抗体的基因的核苷酸序列如seq id no.9所示:
[0170]
gaggtgcagctgctggattctgggggaggctcggtacaggctggagggtctct gagactctcgtgtgaagtttttacaagcaagatagcaaccggctacatgggctggtt ccggcaggctccgggtaaggagcgcgaggccgtcgcaattactcataccggtagtgg aaacccatggtatgccgactccgtgaagggccgcttcaccatctcccaagacaacgc caagctcacggtgtatctgcaaatgaacaacctgagacctgaggacactgccacgt acacctgtgcggcccgggaggcagggtcttgggagctcaggcggttatctccagaa tggtatacctactggggccaggggaccctggtcaccgtctcctca。
[0171]
实施例4纳米抗体nbfm5的大量制备
[0172]
用试剂盒提取携带nbfm5基因(核苷酸序列如seq id no.9所示)的表达载体的菌 株质粒,将其转化到感受态e.coil bl21(de3)中并涂布于lb-agar-amp平板培养得到 单菌落。挑一单菌落接种于10ml lb-amp中,37℃,250rpm培养过夜。将过夜培养物 按1%(v/v)比例接种于100ml lb-amp中,37℃,250rpm培养至对数期,加入iptg 至工作浓度1mm,37℃,250rpm培养过夜。
[0173]
菌液离心并弃去上清,用蔗糖渗透压冻融法提取抗体周质腔蛋白,具体做法为:加入 3.75ml tes溶液重悬沉淀,于-80℃冻结2h以上,37℃水浴融化后加入15ml 5倍稀释的 tes,250rpm震荡2h充分提取蛋白,离心留上清,将上清通过镍-琼脂糖凝胶亲和层析 纯化,即得图3所示的纳米抗体nbfm5(氨基酸序列如seq id no.1所示)。
[0174]
实施例5纳米抗体nbfm5工作浓度及灵敏度的测定
[0175]
一、实验方法
[0176]
1、抗原固定化
[0177]
用包被液(0.375g na2co3与0.7325g nahco3加水定容至250ml)将实施例1制备 的结构式如(ⅲ)所示的完全抗原prm-bsa稀释至2000、1000、500、250ng/ml等一 系列浓度,分别加入酶标板微孔中,每孔100μl,4℃静置过夜。第二天用稀释20倍后的 洗液pbst(0.01m pbs,0.05%tween-20)洗板两次后,每孔加入150μl 1%bsa-pbs(w/v) 溶液37℃静置2h。倒出孔内液体,在吸水纸上拍干并在37℃下烘1h后存于4℃备用。
[0178]
2、确定抗体工作浓度
[0179]
用pbs将纯化后的氨基酸序列如seq id no.1所示的纳米抗体nbfm5稀释分别稀释 成一系列不同浓度的工作溶液。在不同包被浓度的酶标板中加入每孔50μl pbs,再加入 50μl梯度稀释后的抗体,每个抗体浓度共进行3孔重复试验,同时用3个孔进行空白对 照(100μl pbs)。37℃孵育30min,用稀释20倍后的洗液pbst(0.01m pbs, 0.05%tween-20)洗涤微孔5次,在吸水纸上拍干后每孔加入100μl用pbst稀释至5000 倍的anti-vhh-hrp二抗(genscript,a01861-200)。37℃孵育30min,用稀释20倍后的 洗液pbst(0.01m pbs,0.05%tween-20)洗涤微孔5次,在吸水纸上拍干后每孔加入100μl 预先用等体积显色液a与显色液b混合的tmb显色液(solarbio,pr1210),37℃避光 孵育10min,每孔加入50μl终止液10%h2so4(v/v),在酶标仪上读450nm od值。
[0180]
od450nm在1~1.5之间的抗体浓度即为纳米抗体工作浓度,在上述实验条件下,氨 基酸序列如seq id no.1所示的纳米抗体nbfm5的在包被浓度为2000、1000、500、250 ng/ml时的工作浓度分别为43.33、43.33、43.33、54.17ng/ml。
[0181]
3、间接竞争elisa绘制标准曲线
[0182]
将腐霉利标准品用pbs稀释成一系列不同浓度的腐霉利溶液,在不同包被浓度的酶 标板中每孔加入50μl不同浓度的腐霉利,每个浓度进行3孔重复试验,并准备3孔药物 空白组(50μl pbs)。用pbs将氨基酸序列如seq id no.1所示的纳米抗体nbfm5分别 稀释至工作浓度43.33、43.33、43.33、54.17ng/ml,在相应不同包被浓度的孔中加入50μl 稀释后的抗体。37℃孵育30min,用稀释20倍后的洗液pbst(0.01m pbs,0.05%tween-20) 洗涤微孔5次,在吸水纸上拍干后每孔加入100μl用pbst稀释5000倍的anti-vhh-hrp 二抗(genscript,a01861-200)。37℃孵育30min,用稀释20倍后的洗液pbst(0.01m pbs, 0.05%tween-20)洗涤微孔5次,在吸水纸上拍干后每孔加入100μl预先用等体积显色液 a与显色液b混合的tmb显色液(solarbio,pr1210),37℃避光孵育10min,每孔加入 50μl终止液10%h2so4(v/v),在酶标仪上读450nm od值。
[0183]
将药物空白组od450nm值平均值记为b0,将不同药物浓度下的od450nm平均值记 为b
x
,用excel计算不同药物浓度下的b
x
/b0比值以及每组平行数据的标准差。以药物浓 度为横坐标,b
x
/b0比值为纵坐标,在origin软件中绘制散点图并进行logistic函数拟合建 立间接竞争标准曲线。此外,将得到的不同包被浓度的标曲参数ic50、b0、b0/ic50进行 比较选定最佳抗原抗体工作浓度。
[0184]
二、实验结果
[0185]
通过棋盘滴定法优化氨基酸序列如seq id no.1所示的纳米抗体nbfm5的间接竞争 elisa方法的抗原抗体工作浓度。结果如图4所示,随着检测抗原浓度的下降,ic50值 逐
渐下降,b0/ic50值先升后降,在包被浓度为500ng/ml时,ic50值相对比较低且b0/ic50 最大,因此选定包被浓度为500ng/ml,抗体浓度为43.33ng/ml作为此elisa方法的最佳 抗原和抗体浓度。
[0186]
根据以上选择的抗原抗体浓度,基于氨基酸序列如seq id no.1所示的纳米抗体 nbfm5建立的间接竞争elisa标准曲线图如图5所示,可以看出,标准曲线呈s型,线 性相关性较好,检测范围为0.19~2.38ng/ml,ic50为0.67ng/ml,最低检测限(lod)为 0.13ng/ml,检测灵敏度较高。
[0187]
实施例6一种检测腐霉利的方法
[0188]
1、预包被板:用包被液(0.375g na2co3与0.7325g nahco3加水定容至250ml)将 实施例1制备的结构式如(ⅲ)所示的完全抗原prm-bsa稀释至500ng/ml,加入酶标 板微孔中,每孔100μl,4℃静置过夜。第二天用稀释20倍后的洗液pbst(0.01m pbs, 0.05%tween-20)洗板两次后,每孔加入150μl 1%bsa-pbs(w/v)溶液37℃静置2h。 倒出孔内液体,在吸水纸上拍干并在37℃下烘1h后存于4℃备用。
[0189]
2、标记标准品空白孔b0、标准品和样品孔,进行3孔重复试验。在b0中加入50μl pbs,在各标准品孔中加入50μl不同浓度的标准品溶液,在各样品孔中加入50μl经过稀 释的样品溶液。用pbs将氨基酸序列如seq id no.1所示的纳米抗体nbfm5稀释至工作 浓度43.33ng/ml,在以上所有孔中加入50μl稀释后的抗体溶液,37℃孵育30min。
[0190]
3、用稀释20倍后的洗液pbst(0.01m pbs,0.05%tween-20)洗涤微孔5次,在吸 水纸上拍干。
[0191]
4、每孔加入100μl用pbst稀释5000倍的anti-vhh-hrp二抗(genscript, a01861-200),37℃孵育30min,用稀释20倍后的洗液pbst(0.01m pbs,0.05%tween-20) 洗涤微孔5次,在吸水纸上拍干。
[0192]
5、每孔加入100μl预先用等体积显色液a与显色液b混合的tmb显色液(solarbio, pr1210),37℃避光孵育10min。
[0193]
6、每孔加入50μl终止液10%h2so4(v/v),在酶标仪上读450nm od值。
[0194]
三、结果判读
[0195]
将标准品空白od450nm值平均值记为b0,将不同药物浓度下的od450 nm以及样品 待测孔的平均值记为b
x
,计算不同药物浓度或样品孔的b
x
/b0比值以及每组平行数据的标 准差。以标准品浓度的对数值为横坐标,b
x
/b0比值为纵坐标,绘制标准曲线图。根据样 品孔平均吸光度值,可从曲线上得到对应点的横坐标,即为腐霉利浓度的对数值,求得反 对数即为测定液中腐霉利浓度。由于样品经过了预先稀释,因此根据标准曲线所得出的样 品浓度要再乘以其稀释倍数。
[0196]
实施例7一种检测腐霉利的试剂盒
[0197]
一、试剂盒的组成
[0198]
1、预包被bsa偶联腐霉利抗原的可拆酶标板:用包被液(0.375g na2co3与0.7325g nahco3加水定容至250ml)将实施例1制备的结构式如(ⅲ)所示的完全抗原prm-bsa 稀释至500ng/ml,加入酶标板微孔中,每孔100μl,4℃静置过夜。第二天用稀释20倍 后的洗液pbst(0.01m pbs,0.05%tween-20)洗板两次后,每孔加入150μl 1%bsa-pbs (w/v)溶液37℃静置1h。倒出孔内液体,在吸水纸上拍干并在37℃下烘1h。
[0199]
2、酶标试剂:anti-vhh-hrp二抗,使用时需要稀释5000倍。
[0200]
3、标准品:不同浓度的腐霉利标准品溶液。
[0201]
4、抗体与样品的稀释液:pbs。
[0202]
5、标记抗体的稀释液:pbst。
[0203]
6、显色液a与显色液b。
[0204]
7、终止液:10%(v/v)h2so4。
[0205]
8、浓缩洗涤液(20
×
):pbst(0.01m pbs,0.05%tween-20),用于洗板。
[0206]
9、检测抗体:氨基酸序列如seq id no.1所示的纳米抗体nbfm5。
[0207]
二、试剂盒的使用
[0208]
预先进行编号,标记标准品空白孔b0、标准品和样品孔,进行3孔重复试验。在b0中加入50μl pbs,在各标准品孔中加入50μl不同浓度的标准品溶液,在各样品孔中加入 50μl经过稀释的样品溶液。用pbs将氨基酸序列如seq id no.1所示的纳米抗体nbfm5 稀释至工作浓度43.33ng/ml,在以上所有孔中加入50μl稀释后的抗体溶液,37℃孵育 30min。用稀释20倍后的洗液pbst(0.01m pbs,0.05%tween-20)洗涤微孔5次在吸水 纸上拍干后每孔加入100μl酶标记二抗。37℃孵育30min,再用稀释20倍后的洗液pbst (0.01m pbs,0.05%tween-20)洗涤微孔5次,在吸水纸上拍干后每孔加入100μl预先 用等体积显色液a与显色液b混合的tmb显色液(solarbio,pr1210),37℃避光孵育10min,每孔加入50μl终止液,5min内在酶标仪上读450nm的od值。
[0209]
三、结果判读
[0210]
将标准品空白od450nm值平均值记为b0,将不同药物浓度下的od450 nm以及样品 待测孔的平均值记为b
x
,用excel计算不同药物浓度或样品孔的b
x
/b0比值以及每组平行 数据的标准差。以标准品浓度的对数值为横坐标,b
x
/b0比值为纵坐标,绘制标准曲线图。 根据样品孔平均吸光度值,可从曲线上得到对应点的横坐标,即为腐霉利浓度的对数值, 求得反对数即为测定液中腐霉利浓度。由于样品经过了预先稀释,因此根据标准曲线所得 出的样品浓度要再乘以其稀释倍数。
[0211]
实施例8纳米抗体nbfm5的特异性测定
[0212]
一、实验方法
[0213]
以实施例1制备的结构式如(ⅲ)所示的完全抗原prm-bsa为检测抗原,纳米抗体 nbfm5为检测抗体,配制其他9种腐霉利的类似物(乙烯菌核利、乙烯利、菌核净、异 菌脲、灭菌丹、滴滴涕、乙滴涕、六六六)以及腐霉利代谢物(3,5-二氯苯胺)标准品 溶液,使用实施例7的试剂盒,按照实施例5中的间接竞争elisa方法绘制每一种腐霉 利类似物及代谢物的标准曲线,得出各自的ic50值。用以下公式计算各药物与纳米抗体 nbfm5的交叉反应率:
[0214][0215]
二、实验结果
[0216]
结果显示,纳米抗体nbfm5与腐霉利代谢物3,5-二氯苯胺以及其类似物乙烯菌核 利、乙烯利、菌核净、异菌脲、灭菌丹、滴滴涕、乙滴涕、六六六的交叉反应率均低于 0.1%(ic50大于700ng/ml);说明纳米抗体nbfm5能够特异性地识别腐霉利,本发明 所建立的检
测方法对于腐霉利检测的特异性高。
[0217]
实施例9纳米抗体nbfm5在不同浓度有机溶剂下中的活性测定
[0218]
一、实验方法
[0219]
用pbs分别配制不同体积比(0%、20%、40%、60%、80%、100%)的甲醇、乙腈、 丙酮溶液,把腐霉利标准品统一稀释至实例5中建立的标准曲线抑制率约为80%的药物浓 度3ng/ml。用pbs把氨基酸序列如seq id no.1所示的纳米抗体nbfm5稀释至工作浓 度43.33ng/ml。效价孔每孔先加入50μl不同体积比有机溶剂配制的标准品稀释液,再 加入50μl稀释后的氨基酸序列如seq id no.1所示的纳米抗体溶液。抑制孔每孔先加入 50μl已稀释至3ng/ml的腐霉利标准品溶液,再加入50μl稀释后的氨基酸序列如seq idno.1所示的纳米抗体溶液。
[0220]
使用实施例7中的检测试剂盒,分别测定在不同有机溶剂条件下,抗体与检测抗原以 及腐霉利标准药物(3ng/ml)的结合能力,评价氨基酸序列如seq id no.1所示的纳米抗 体nbfm5对不同有机溶剂的耐受能力。
[0221]
二、实验结果
[0222]
以上抗体是使用pbs作为稀释液的,因此抗体接触到的实际有机浓度分别为0%、 10%、20%、30%、40%、50%。抑制率可由实例3的抑制率公式计算可得。测定结果如图 6所示,氨基酸序列如seq id no.1所示的纳米抗体nbfm5在10%以下的低浓度甲醇溶 液中与检测抗原以及小分子的结合活性受影响较小,而乙腈以及丙酮溶液浓度达到10% 时,抗体与检测抗原的结合活性只能保留30%、40%。另外,nbfm5与检测抗原以及小 分子药物的结合活性都随着有机溶剂浓度的提升急剧下降。当有机溶剂混合液中甲醇、乙 腈、丙酮分别比例达到40%、20%、20%时抗体完全失活。
[0223]
说明氨基酸序列如seq id no.1所示的纳米抗体nbfm5对这三种在标准方法前处理 当中常用的有机溶剂(甲醇、乙腈、丙酮)耐受性较差,应该在抗体工作溶液中尽量除去 有机溶剂,避免高浓度有机溶剂对抗体的灵敏度产生影响。
[0224]
实施例10纳米抗体nbfm5在不同温度下的活性测定
[0225]
一、实验方法
[0226]
用pbs氨基酸序列如seq id no.1所示的纳米抗体nbfm5稀释至工作浓度(43.33 ng/ml)后分别在高温70℃条件下孵育不同的时间(0、10、20、30、40、50、60min), 再用pbs把腐霉利标准品稀释至实例5中建立的标准曲线抑制率约为80%的药物浓度3 ng/ml。使用实施例7的试剂盒,分别测定抗体在高温处理不同时间后与检测抗原以及腐 霉利标准药物(3ng/ml)的结合活性。
[0227]
二、实验结果
[0228]
抑制率可由实例3实验方法中的抑制率公式计算可得。测定结果如图7所示,氨基酸 序列如seq id no.1所示的纳米抗体nbfm5与检测抗原结合活性以及小分子待测物的结 合活性随孵育时间的延长呈下降趋势,其中纳米抗体与小分子待测物的结合活性下降趋势 较为平缓。即使在70℃条件下加热1个小时后,氨基酸序列如seq id no.1所示的纳米 抗体nbfm5仍具有50%的抗原结合活性与66%的抑制率。说明氨基酸序列如seq id no.1 所示的纳米抗体nbfm5可在高温变性后重新折叠恢复大部分的抗原结合能力,这个特性 可使其在较为高温的检测环境中有较大的应用优势。
[0229]
实施例11纳米抗体nbfm5在不同ph条件下的活性测定
[0230]
一、实验方法
[0231]
用不同ph值(2.4、3.4、5.4、6.4、7.4、8.4、9.4、10.4)的0.01m pbs作为稀释液, 分别将氨基酸序列如seq id no.1所示的纳米抗体nbfm5的溶剂稀释至工作浓度(43.33 ng/ml),将腐霉利标准品稀释至实例5中建立的标准曲线抑制率约为80%的药物浓度3 ng/ml。使用实施例7的试剂盒,分别测定抗体与检测抗原以及腐霉利标准药物(3ng/ml) 的结合能力,评价氨基酸序列如seq id no.1所示的纳米抗体nbfm5不同ph条件下中 与检测抗原以及腐霉利标准药物的结合活性。
[0232]
二、实验结果
[0233]
抑制率可由实例3的抑制率公式计算可得。测定结果如图8所示,在ph 5.4~9.4范 围内,氨基酸序列如seq id no.1所示的纳米抗体nbfm5与检测抗原以及小分子待测物 的结合活性基本稳定不变,在这个范围外,随着ph偏离中性程度越来越大,抗体与检测 抗原以及小分子待测物的结合活性逐渐下降。当ph下降至到2.4时,抗体基本失活,而 当ph达到10.4时,抗体抑制率仅降30%左右。总体趋势而言,nbfm5可以在较宽的ph 范围内保持较好的抗原结合活性,但是更适合在碱性环境下工作。
[0234]
实施例12纳米抗体nbfm5在不同温度条件下的储存稳定性测定
[0235]
一、实验方法
[0236]
在氨基酸序列如seq id no.1所示的纳米抗体nbfm5溶液中分别加入0.03%防腐剂 proclin 300(v/v)以及0.01%蛋白酶抑制剂(v/v),分装20μl每份,分别放置到日常使用中最 常见的四个温度(-20、4、25、37℃)中避光储存。使用实施例7的试剂盒,分别测定氨 基酸序列如seq id no.1所示的纳米抗体nbfm5在不同温度条件下储存不同时间(0、7、 14、30、60day)后与检测抗原以及腐霉利标准药物(3ng/ml)的结合能力。
[0237]
二、实验结果
[0238]
一般商品化抗体保存都需要加防腐剂或者蛋白酶抑制剂,本实施例证明了添加防腐剂 后的抗体储存稳定性,确定其货架期,以便在实际应用中更具有参考价值。抑制率可由实 例3实验方法中的抑制率公式计算可得。结果如图9所示,氨基酸序列如seq id no.1 所示的纳米抗体nbfm5在添加了防腐剂以及蛋白酶抑制剂后,在不同温度中储存一个月, 其与检测抗原以及腐霉利标准药物的结合活性基本不变,甚至储存时间达到两个月时,只 有在37℃条件下储存的纳米抗体nbfm5抗原结合活性降至41%,其他温度条件下储存的 抗体的结合活性基本仍为100%。说明氨基酸序列如seq id no.1所示的纳米抗体nbfm5 具有高度的储存稳定性,进行一般的防腐试剂添加后可以在室温条件下长时间运输、储存 和使用。
[0239]
最后所应当说明的是,以上实施例仅用以说明本发明的技术方案而非对本发明保护范 围的限制,对于本领域的普通技术人员来说,在上述说明及思路的基础上还可以做出其它 不同形式的变化或变动,这里无需也无法对所有的实施方式予以穷举。凡在本发明的精神 和原则之内所作的任何修改、等同替换和改进等,均应包含在本发明权利要求的保护范围 之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1