一种高抗氧化活性富硒肽及其应用
1.本发明属于抗氧化技术领域,具体涉及一种高抗氧化活性的富硒肽及其应用。
背景技术:
2.硒(se)是维持人和动物健康所必需的微量营养素之一,具有重要的生理功能,包括抗氧化,抗癌,抗疲劳和免疫调节等。据估计,全世界有10亿人因硒摄入不足而诱发系列疾病。研究发现,天然有机硒比无机硒相比具有更多优势,比如低毒性、生物利用率高和多种生理活性。因此,近年来对天然富硒产品的研究越来越多,其中包括富硒大米、富硒绿茶、富硒双孢菇、富硒酵母等。
3.辣木属辣木科,是原产于热带或南亚热带干旱或半干旱地区的多年生木本植物。大量研究表明,辣木叶、种子和根提取物对人体有许多益处,如抗糖尿病、保护肝脏,抗尿石和改善肝纤维化的作用。辣木籽蛋白的含量高达37.48%,其中val、leu、ile、his等多种氨基酸的含量接近或远高于联合国粮食及农业组织 /世界卫生组织标准模式谱,可作为优质蛋白质资源加以开发。辣木籽蛋白水解物具有多种生物活性,包括抗高血压、抗糖尿病,抗炎和减少疼痛相关的腹部绞痛的功效。然而,目前关于辣木籽蛋白肽的研究多以混合物形式开展,对其多肽组成、氨基酸序列及活性机制研究有待深入。目前已有专利一种具有肝细胞氧化损伤保护作用的天然寡肽公开了一种从辣木籽中分离得到的具有细胞损伤保护作用的多肽,但其并非富硒多肽。研发一种富硒辣木籽活性肽将极具研究意义。
技术实现要素:
4.本发明的目的在于克服现有技术的缺陷或不足,提供一种高抗氧化活性的富硒肽。本发明从富硒辣木籽蛋白粉中分离提纯得到一种高抗氧化活性的富硒肽,该富硒肽具有高硒含量,且具有高抗氧化活性和细胞损伤保护作用,可阻断 keap1-nrf2的相互作用。
5.本发明的另一目的在于提供上述富硒肽在制备抗氧化剂中的应用。
6.本发明的另一目的在于提供上述富硒肽制备细胞氧化损伤保护剂中的应用。
7.本发明的另一目的在于提供上述富硒肽在制备阻断keap1-nrf2的相互作用的阻断剂中的应用。
8.为实现上述发明目的,本发明采用如下技术方案:
9.一种高抗氧化活性的富硒肽,所述富硒肽具有如下氨基酸序列:flml(如 seqidno:1,phe-leu-met-leu)或lammvl(如seqidno:2, leu-ala-met-met-val-leu),且所述flml中的甲硫氨酸m中的硫元素被硒取代,记为flseml;所述lammvl中的第一个甲硫氨酸m中的硫元素被硒取代,记为lasemmvl。
10.本发明的发明人从富硒辣木籽中分离出两种具有如下序列的富硒肽:flml 和lammvl,且特定位点的甲硫氨酸m中的硫元素被硒取代。该富硒肽不仅具有较好的硒含量,且具有显著的的抗氧化活性,对细胞损伤具有较好的保护作用。
11.优选地,所述富硒肽来源于富硒辣木籽。
12.更为优选地,所述富硒肽来源于富硒辣木籽蛋白粉。
13.优选地,所述富硒肽的分子量为570.2321da或724.3096da。
14.具体地,当富硒肽为flseml时,其分子量为570.2321da。当富硒肽为 lasemmvl时,其分子量为724.3096da。
15.本发明还请求保护上述富硒肽在制备抗氧化剂中的应用。
16.本发明提供的富硒肽具有高抗氧化活性,其可作为天然抗氧化剂添加至食品、化妆品、保健品或药品。
17.优选地,所述富硒肽制备食品、化妆品、保健品或药品中的应用。
18.更为优选地,所述富硒肽在食品、化妆品、保健品或药品中的添加量为 0.291~0.662μg/ml。
19.上述富硒肽在制备细胞氧化损伤保护剂中的应用也在本发明的保护范围内。
20.本发明的富硒肽可有效降低胞内活性氧积累,显著提高被损伤细胞中的超氧化物歧化酶(sod)和过氧化氢酶(cat)活性。
21.上述富硒肽在制备阻断keap1-nrf2的相互作用的阻断剂中的应用也在本发明的保护范围内。
22.本发明的富硒肽可以阻断keap1-nrf2的相互作用,使得其可作为阻断 keap1-nrf2的相互作用的阻断剂。
23.与现有技术相比,本发明具有如下有益效果:
24.本发明从富硒辣木籽蛋白粉中分离提纯得到一种高抗氧化活性的富硒肽,该富硒肽具有高硒含量,且具有高抗氧化活性和细胞损伤保护作用,可阻断 keap1-nrf2的相互作用。
附图说明
25.图1为f1-3组分对dpph和abts自由基清除率;
26.图2为flseml和lasemmvl的二级质谱图;
27.图3为flseml和lasemmvl对hepg2细胞毒性作用实验结果;
28.图4为flseml和lasemmvl的细胞抗氧化活性实验结果;
29.图5为aaph诱导hepg2细胞氧化应激损伤实验结果;
30.图6为flseml和lasemmvl对细胞损伤的保护作用实验结果;
31.图7为flseml和lasemmvl对ros的抑制效果;
32.图8为flseml和lasemmvl对aaph诱导损伤的hepg2细胞中sod (图8a)和cat(图8b)抗氧化酶活性的影响试验结果;
33.图9为flseml和lasemmvl与人源性keap1蛋白的结合模式。
具体实施方式
34.下面结合实施例进一步阐述本发明。这些实施例仅用于说明本发明而不用于限制本发明的范围。下例实施例中未注明具体条件的实验方法,通常按照本领域常规条件或按照制造厂商建议的条件;所使用的原料、试剂等,如无特殊说明,均为可从常规市场等商业途径得到的原料和试剂。本领域的技术人员在本发明的基础上所做的任何非实质性的变化
及替换均属于本发明所要求保护的范围。
35.实施例中提及的f1-3组分通过如下过程制备得到:
36.1.富硒辣木籽蛋白的酶解:将1.5g富硒辣木籽蛋白粉(商购得到)溶于30 ml水中(s/l:5%),调ph至1。然后加入1%(e/s:0.32%)的胃蛋白酶。水解1.64h,在90℃加热10min后冷却至室温,4000r/min离心20min,收集上清液,4℃保存备用。
37.2.富硒辣木籽蛋白酶解物的分离:
38.(1)使用装有截留分子量为10kda超滤膜的amicon ultra-15离心超滤管对富硒辣木籽蛋白酶解液(步骤1得到的上清液)进行分离,离心机转速为4000 rpm,离心时间20min。收集分子量小于10kda的组分,-20℃保存备用。
39.(2)将超滤分离获得的组分(《10kda)过0.45μm滤膜后,采用c18反相色谱柱(20mm
×
450mm,10μm)进行制备液相色谱分离。色谱条件为:流动相a:双蒸水+0.1%(体积分数,下同)三氟乙酸(trifluoroacetic acid,tfa);流动相b:乙腈+0.1%tfa;梯度洗脱:0~40min,6%~25%流动相b;40~70min, 25%~70%流动相b;70~75min,70%~90%流动相b;75~85min,90%~90%流动相b;85~90min,90%~6%;进样量4ml;流速10ml/min;检测波长280nm。收集各洗脱峰,得到7个组分,按收集的时间顺序依次命名为f1~f7。
40.3.f1组分的分离纯化:对f1组分进行第二次分离纯化,色谱条件:shimadzuprc-ods(k)钢柱(30mm
×
250mm,15μm);流动相a:双蒸水+0.1%三氟乙酸;b:甲醇+0.1%三氟乙酸;梯度洗脱:0~10min,5%~10%流动相b;10~40min, 10%~20%流动相b;40~45min,20%~90%流动相b;45~55min,90%~90%流动相b;55~65min,90%~5%;上样量4ml洗脱流速10ml/min,在280nm的紫外波长下监测洗脱峰。收集得到5个活性组分,按收集的时间顺序依次命名为 f1-1~f1-5。f1-3组分进行后续处理。
41.实施例1富硒肽的分离纯化
42.从富硒辣木籽蛋白酶解物中分离得到的f1-3组分,对该组分的dpph和 abts自由基清除率及硒含量进行测定。测试过程及结果如下:
43.(1)dpph自由基清除率
44.测试过程:
45.将100μl样品溶液加入到100μl 0.15mm dpph溶液中。然后将混合物在室温下黑暗中放置30分钟。使用酶标仪在517nm处测量96孔板的吸光度。dpph 自由基清除率的计算公式为:
46.dpph自由基清除率(%)=[1
ꢀ‑
(a
样品-a
空白
)/a
对照
]
×
100 (1)
[0047]a样品
:100μl样品溶液加入100μl dpph溶液的吸光度;
[0048]a空白
:100μl样品溶液加入100μl乙醇的吸光度;
[0049]a对照
:100μl dpph溶液加入100μl蒸馏水的吸光度。
[0050]
测试结果:如图1,在0.5mg/ml时,f1-3组分的dpph自由基清除率为 38.10%。
[0051]
(2)abts自由基清除率
[0052]
测试过程:
[0053]
将7mm abts和2.45mm过硫酸钾溶液充分混合,在室温下黑暗中反应12 小时。然后用蒸馏水稀释混合物至吸光度在734nm处为0.70
±
0.02,得到abts 工作液。将100μl abts溶液与100μl样品溶液混合,振荡1min,在37℃恒温下反应10min后,在734nm进行检测。abts
自由基清除活性的计算公式为:
[0054]
abts自由基清除率(%)=[1-(a
样品-a
空白
)/a
对照
]
×
100(2)
[0055]a样品
:100μl样品溶液加入100μl abts溶液的吸光度;
[0056]a空白
:100μl样品溶液加入100μl蒸馏水的吸光度;
[0057]a对照
:100μl abts溶液加入100μl蒸馏水的吸光度。
[0058]
测试结果:如图1,在0.5mg/ml时,f1-3组分的abts自由基清除率为 34.29%。
[0059]
(3)硒含量
[0060]
根据国家标准gb 5009.93-2017《食品中硒的测定》,采用氢化物原子荧光光谱法(hg-afs)测定总硒含量。称取适量样品置于锥形瓶中,用10ml浓硝酸和高氯酸混合物(v/v,9:1)在150℃下消化2.5h,冷却后,在消化后的样品中加入5ml 6mol/l盐酸以还原se
6+-se
4+
。冷却后,用超纯水将消化后的溶液稀释至25ml,然后将10ml的溶液转移到反应容器中,其中加入2ml为6mol/l 盐酸和1ml 10%(w/w)铁氰化钾,混匀待测,同时做空白对照。
[0061]
除f1-3组分作为样品外,还以辣木籽蛋白酶解物作为样品,对比硒含量的变化。
[0062]
仪器条件为:负高压280v,硒灯电流80ma,原子化器高度8mm,载气流量为300ml/min;屏蔽气流量为800ml/min,读数时间12s,延迟时间3s,重复测定3次,测定样品硒含量,测试结果如表1。
[0063]
表1富硒辣木籽蛋白酶解物和f1-3组分的硒含量
[0064][0065]
从表1可知,f1-3组分中的硒含量为13.772mg/kg,是富硒辣木籽蛋白酶解物中硒含量(2.253mg/kg)的6倍多。这一结果表明,经分离提纯后,更高硒含量的肽段被富集,这也间接说明硒在辣木籽中多肽的抗氧化活性上发挥了较大作用。
[0066]
在此基础上,利用lc-ms/ms对f1-3组分中肽段的氨基酸序列进行鉴定,具体过程如下:
[0067]
取多肽样品(即分离得到的f1-3组分)溶于washing buffer(0.1%fa,2% acn),经c18除盐柱进行脱盐处理后,利用在线纳喷离子源的lc-ms/ms分析样品的肽段组成和氨基酸序列。整套系统为串联easy-nanolc 1200的q exactive
tm plus质谱仪(thermo fisher scientific,ma,usa)。色谱条件:色谱柱(acclaim pepmap c18,75μm
×
25cm);柱流量控制在300nl/min;柱温为 40℃;进样量5μl;电喷雾电压1.9kv;流动相a(0.1%甲酸水溶液)、流动相 b(含0.1%甲酸的乙腈溶液,洗脱梯度为:0~60min,5%~38%b。质谱仪在数据依赖采集模式下运行,自动在ms和ms/ms采集间切换。质谱参数设置如下: (1)一级质谱扫描范围(m/z):100-1500;分辨率:70000;自动增益控制目标: 3e6;最大进样时间:100ms;包括电荷状态:1-3;(2)二级质谱自动增益控制目标:8e3;最大进样时间:50ms;碰撞能量:28;动态排除时间为6s。使用 de novo软件(peaks studio 8.5)对ms/ms数据对照ncbi数据库进行序列搜索,选择甲硫氨酸氧化,乙酰化,脱酰胺化,谷氨酸及谷氨酰胺环化,氨基甲基化和硒
取代硫进行可变修饰。前体耐受量为10ppm,产物离子耐受量为0.05da。在辣木(moringa oleifera)数据库中搜索获得的质谱数据,选择平均置信度(alc) 大于50%的肽序列。
[0068]
共鉴定得到103个富硒肽,由4-10个氨基酸残基组成,分子量约为0.5-1.2 kda。
[0069]
疏水性氨基酸的存在对自由基的清除起着重要作用,可以增加肽在脂质介质中的溶解度,支持疏水自由基的可及性。脂质抑制活性也通过肽和自由基的相互作用增强。故随后通过多肽疏水性氨基酸占比分析,筛选得到疏水性占比很高的 2个富硒肽:flseml(570.2321da,sem代表该甲硫氨酸m中的硫元素被硒 se取代,下同)和lasemmvl(724.3096da),其二级质谱图见图2(m代表 sem),分子量鉴定结果如表2。
[0070]
表2 f1-3组分中富硒肽的鉴定
[0071][0072]
实施例3富硒肽的抗氧化活性测定
[0073]
对实施例2得到的两种富硒肽的抗氧化活性进行测定。
[0074]
(1)富硒肽对hepg2细胞毒性作用(mtt)检测
[0075]
hepg2细胞培养:10%fbs混合dmem培养基,将细胞置于5%co2细胞培养箱内进行培养。待hepg2细胞贴壁生长,融合成致密单层且细胞覆盖率达 80%以上时,用0.25%胰蛋白酶溶液消化传代或进行下一步试验。
[0076]
富硒肽对hepg2细胞的毒性检测:将1
×
105cells/ml的hepg2细胞接种于 96孔板中(每组设置6个复孔),设置对照组和试验组,对照组和试验组加入100 μl细胞悬液,培养24h细胞贴壁,对照组加入100μl的培养液,试验组加入 100μl样品溶液,在37℃,5%co2培养箱中孵育24h后,弃去旧培养液,加入100μl mtt(0.5mg/ml)孵育4h。最后,每孔加入100μl dmso溶解形成的甲瓒盐,酶标仪检测490nm处吸光值。细胞存活率计算公式:
[0077]
hepg2细胞存活率(%)=a
试验组
/a
对照组
×
100
ꢀꢀꢀꢀ
(3)
[0078]a试验组
为试验组的吸光值,a
对照组
为空白组的吸光值。
[0079]
具体地,样品溶液为0.0001、0.0005、0.005、0.05、0.1、0.25和0.5mg/ml。
[0080]
测试结果如图3,在浓度0.1mg/ml范围内时,flseml和lasemmvl对 hepg2细胞均无细胞毒性作用。当浓度为0.1mg/ml时,flseml和lasemmvl 的细胞活力分别为93.79%和91.34%。因此可以选择该浓度范围的富硒肽进行后续的细胞抗氧化实验。
[0081]
(2)细胞内抗氧化活性(caa)的测定
[0082]
caa的测定方法如下:hepg2细胞以6
×
104cells/孔的密度接种在黑色96 孔板上。培养24h后,除去培养基,用100μl pbs洗涤一遍。加入100μl富硒肽溶液(0.0005-0.025mg/ml)或溶剂对照(含有终浓度25μm dcfh-da),放入培养箱培养1h后,吸出溶液并用pbs缓冲溶液洗涤3次,加入100μl 600μm aaph溶液,并在空白组使用hbss缓冲液代替aaph溶液。最后,在37℃下每隔5min读取一次荧光,发射波长为528nm,激发波长为485nm,总测量时间1h。通过对每个时间点荧光曲线下的面积与时间曲线积分,计算富硒肽的caa值,公式如下:
[0083]
caa(%)=100
×
[1-(auc
sample-auc
nc
)/(auc
pc-auc
nc
)]
ꢀꢀꢀ
(4)
[0084]
其中,auc
sample
为样品的曲线下面积;auc
nc
为阴性对照(单独使用 dcfh-da处理的
细胞)的曲线下面积;auc
pc
为阳性对照(用dcfh-da和 aaph处理的细胞)的曲线下面积。
[0085]
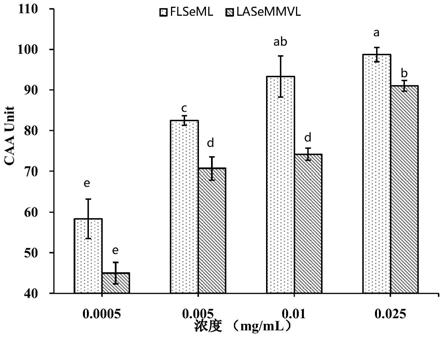
如图4,为富硒肽的细胞抗氧化活性实验结果。从图4可知, flseml和lasemmvl的caa值均随着浓度的增加而上升,并呈良好的剂量-效应关系。从ec
50
值可知,flseml和lasemmvl均在hepg2细胞中表现出了优异的抗氧化活性,flseml和lasemmvl的ec
50
值分别为0.291μg/ml和 0.662μg/ml,显著低于从桑叶蛋白中鉴定的三种抗氧化肽svl(240μg/ml)、 eavq(310μg/ml)和rdy(240μg/ml)。由此可以看出flseml和lasemmvl 具有极低的ec
50
值(如表3),在极低浓度下即展现极强的细胞抗氧化活性,这可能与活性肽中硒元素的协同抗氧化作用具有直接关系。
[0086]
表3富硒肽细胞抗氧化活性的ec
50
值
[0087][0088]
(3)aaph诱导hepg2细胞氧化应激损伤的模型建立
[0089]
aaph被用作过氧自由基的诱导剂,与细胞膜的多不饱和脂质相互作用,通过细胞膜渗透到细胞内部,导致活性氧的产生,进而对细胞产生氧化损伤作用。因此,aaph通常用于诱导氧化应激,用于构建细胞氧化损伤模型试验。
[0090]
aaph损伤模型的构建方法如下:将密度为1
×
105个/ml的hepg2细胞接种于96孔板,每孔100μl,在培养箱孵育24h后,弃去上清液并向每孔加入100 μl aaph溶液(终浓度为0.1-25mm)和100μl dmem培养基(对照组)。孵育24h后,用上述mtt法测定细胞存率。
[0091]
如图5所示,当aaph浓度》0.5mm时,细胞内氧化应激显著增加(p《0.05),当aaph浓度为2.5mm时,细胞存活率从100%降为78.18%,随着浓度增加细胞存活率也呈显著下降趋势。考虑到氧化应激损伤程度太弱或者太强都不利于后续的损伤保护实验,故选取aaph的浓度2.5mm作为损伤实验浓度用于构建 hepg2细胞氧化损伤模型。
[0092]
(4)富硒肽对aaph诱导损伤的hepg2细胞的保护作用
[0093]
每孔100μl hepg2细胞(1
×
105个/ml)接种于96孔板,在培养箱中孵育 24h后,弃去上清液,并向保护组的每孔中加入100μl富硒肽溶液(浓度为0.001、 0.005、0.025mg/ml)。将100μl细胞培养液加入到损伤组和对照组中,继续培养24h后,向保护组和损伤组中加入100μl 2.5mm的aaph溶液。对照组每孔加入100μl细胞培养液。培养24h后按照mtt法测定细胞存活率。
[0094]
富硒肽对细胞损伤的保护作用见图6。从图6可以看出,与空白对照组相比,经aaph(浓度:2.5mm)处理后损伤组的细胞存活率显著降低至78.72% (p《0.05),表明细胞损伤模型造模成功且稳定可靠。与单纯的损伤组相比,经 2个富硒肽处理的实验组的细胞存活率均有不同程度的改善。随富硒肽浓度增加细胞存活率出现显著提升,当浓度为0.025mg/ml时,经富硒肽flseml和 lasemmvl处理的hepg2细胞存活率分别从78.72%显著提升至90.71%和 93.92%。
[0095]
(5)富硒肽对aaph诱导损伤的hepg2细胞内ros含量的影响
[0096]
ros是需氧细胞在代谢过程中产生的一系列活性氧簇,中、高浓度的ros 可通过细胞氧化应激诱导细胞凋亡甚至死亡。dcfh-da本身无荧光,可自由穿透细胞膜,细胞内ros可将dcfh-da氧化成带荧光的dcf,基于此原理可以定性定量的检测细胞内ros的动态变化。
[0097]
按照(4)富硒肽对aaph诱导损伤的hepg2细胞的保护作用中的方法将细胞分组,培养结束后,弃各孔培养液。按照如下方法对细胞内活性氧ros的研究方法检测:将稀释于dmem中的dcfh-da(10μm)溶液加入各孔中,孵育30min,并用pbs洗涤3次。用多功能酶标仪(激发波长485nm,发射波长 528nm)检测荧光强度。
[0098]
flseml和lasemmvl对ros的抑制效果见图7。与对照组相比,损伤组细胞内的荧光强度显著增强了1.35倍,表明aaph诱导细胞氧化损伤后细胞内产大量ros。然而,经不同浓度富硒肽处理组的ros水平显著低于损伤组 (p《0.05),且随浓度增大效果更明显。在浓度0.025mg/ml时,flseml和 lasemmvl处理组的ros含量分别比损伤组显著下降了44.45%和37.95%,这说明flseml和lasemmvl可以有效抑制细胞内ros的产生和堆积,进而起到保护细胞被氧化损伤的作用。
[0099]
(6)富硒肽对aaph诱导损伤的hepg2细胞内抗氧化酶活性的影响
[0100]
细胞内的部分酶,比如常见的sod和cat等,在机体生理活动中起到抗氧化的防御作用,它们可以消除生物体中多余活性氧。因此,细胞内抗氧化酶活力的变化成为测定抗氧化活性的标志物。
[0101]
按照(4)富硒肽对aaph诱导损伤的hepg2细胞的保护作用中的方法对细胞进行分组,在6孔板中接种培养hepg2细胞(0.5
×
106cells/孔),每孔2ml。将富硒肽溶液(最终浓度为0.001、0.005、0.025mg/ml)加入保护组。最后用 aaph诱导损伤组和保护组。细胞用0.25%胰酶消化,培养基终止,用pbs洗涤细胞,1000r/min离心10min收集细胞,-20℃保存备用。每孔收集的细胞加入 0.5ml pbs溶解沉淀制成细胞悬浮液,用超声波粉碎仪破碎细胞,采用超氧化物歧化酶(sod)测定试剂盒和过氧化氢酶(cat)测定试剂盒测定细胞sod 和cat活性。采用bca试剂盒测定细胞裂解物中的蛋白质浓度,使sod和cat 的水平正常化。测定结果以每毫克蛋白质的酶活性单位给出。
[0102]
2个富硒肽对aaph诱导损伤的hepg2细胞中sod和cat抗氧化酶活性的影响试验结果分别见图8a和图8b。aaph处理hepg2细胞后,损伤组的sod 和cat的活性与对照组相比明显被抑制(p《0.05),而经富硒肽的预处理后,它们的活性随着肽浓度的增加而增加,尤其在高剂量组(0.025mg/ml)预处理下, 2个肽均能够将超氧化物歧化酶提高到与正常细胞相当的水平;flseml和 lasemmvl处理组细胞的cat活力与对照组无明显差异(p》0.05)。结果表明富硒肽对细胞氧化损伤的保护作用与肽能提高细胞内抗氧化酶的活力有内在关系。
[0103]
实施例3基于分子对接技术研究富硒肽的抗氧化机制
[0104]
分子对接是一种基于模拟分子相互作用的计算机方法,可用于预测目标活性物与受体蛋白之间的作用位点和作用方式。单磷酸腺苷激活的蛋白激酶(ampk) 介导的keap1-nrf2信号通路是体内细胞对各种外源性和内源性因素引起的氧化应激情况下机体进行自我防御以及保护的重要通路。抗氧化应激反应能够被组织内的nrf2蛋白激活,从而发挥清除体内自由基的作用。在正常情况下,keap1 分子能够与nrf2分子结合,促进nrf2被降解,使得抗氧化应激反应被抑制。抗氧化剂分子通过结合keap1的kelch结构域,占据keap1-nrf2通路间的结合位点,抑制其相互作用并有效激活抗氧化通路。自带的配体16mer nrf2通过与 keap1蛋白kelch结构域结合的关键残基有arg380、arg415和arg483的三个 arg残基,ser363、ser508、ser555和ser602的四个ser残基,以及tyr334、asn382 和gln530。此外,asn387、his436、tyr525、tyr572和phe577的残基在keap1-nrf2 蛋白-蛋白相互作用中发挥
关键作用。
[0105]
在本实施例中,为了探索富硒肽与人源性keap1受体蛋白作为潜在抑制剂的可能结合模式,采用autodock vina程序进行分子对接,预测活性肽的作用位点和抗氧化机制。keap1蛋白的晶体结构(编码:4l7b)从蛋白质数据库 (http://www.rcsb.org/pdb/home/home.do)下载。用pymol软件去除受体分子中含有的水分子原始配体,保存为pdb格式文件。随后用autodock tools 1.5.6软件进行加氢,保存为pdbqt格式文件备用。配体小分子结构由marvinsketch软件绘制,并构建和优化配体的三维结构,使用autodock tools打开,并自动添加 gasteiger电荷,设置旋转键,保存为pdbqt文件。采用autodock tools进行蛋白质-配体对接的分析,对接结果以结合能值表示,值越低代表小分子与蛋白结合得越紧,其抑制活性越强。此外,使用discovery studio 4.5软件对与结合位点残基相互作用的所有模式进行了可视化。
[0106]
2个富硒肽与人源性keap1蛋白的结合模式见图9。从图9a(1)和9b(1) 可以看出,2个富硒肽成功对接到keap1口袋中,可见2个肽的结合位置均为同一个疏水空腔,并占据nrf2的部分结合位点,从而直接阻断keap1和nrf2之间的蛋白质-蛋白质相互作用。2个富硒肽与受体蛋白keap1作用位点及相互作用力见图9a(2)至9b(2)以及表4。
[0107]
flseml对keap1的疏水位置(-8.9kcal/mol)具有最高亲和力,其主要与 ser555和asn414形成两个氢键,与tyr334,tyr525,tyr572,arg415和phe577 形成疏水相互作用。其中包括此肽的一个苯环与tyr334的苯环之间存在1个π-π t型相互作用,与phe577形成π-π堆积效应。在这些结合残基中,ser555,tyr334, tyr525,tyr572,arg415和phe577是keap1-nrf2相互作用的关键残基,表明小肽可能干扰keap1-nrf2相互作用。
[0108]
lasemmvl主要与arg415形成4个氢键,与arg380、和asn382形成2 个氢键,还与asn414、ala556和gly509形成3个氢键,与tyr572形成π-硫键。
[0109]
另外这两个肽均与附近的氨基酸残基形成各种非共价键。所有这些相互作用促使肽紧密结合在keap1的结合位点。
[0110]
这些结果表明flseml和lasemmvl均可以插入keap1的口袋并介导nrf2 的激活,均为keap1的抑制剂。因此,flseml和lasemmvl能够与keap1 蛋白的关键氨基酸残基以及附近的氨基酸发生相互作用,阻碍keap1-nrf2的相互作用,维持组织高nrf2水平,进而激活抗氧化应激反应,从而增强机体清除自由基的能力。另外,flseml和lasemmvl中硒原子连接甲基的c原子可与 arg415、typ572和phe577形成疏水相互作用。硒可能影响或直接构成肽的活性中心,可以催化或直接参与自由基的清除,增强肽的抗氧化能力。研究表明, se-met是蛋氨酸中的硫被硒取代所形成的一种有机硒,作为植物中硒的主要存在形式之一,可以抑制生长迟缓,对氧化应激和细胞凋亡的影响很大。富硒辣木籽中有机硒主要以硒代蛋氨酸形式存在,因此推测硒代蛋氨酸和肽可能对富硒肽的抗氧化活性有协同作用。
[0111]
表4富硒肽与keap1(4l7b)的分子对接结果
[0112][0113]
从上述各实施例可知,本发明首次从富硒辣木籽蛋白酶解物中纯化鉴定出2 个新型富硒肽(flseml和lasemmvl)并对其活性机制进行了研究。细胞实验表明,flseml和lasemmvl显示了优秀的细胞抗氧化活性,而且对aaph 诱导的hepg2细胞氧化损伤具有明显的保护作用。更重要的是,flseml和lasemmvl均可通过降低ros保护aaph诱导的hepg2细胞免受氧化应激,增强内源性抗氧化酶(sod和cat)的防御能力系统。为了进一步了解富硒肽的抗氧化活性机制,通过分子对接模拟预测肽与keap1之间作用位点和作用方式,结果表明flseml和lasemmvl通过结合keap1口袋中的nrf2位点,与 tyr334,tyr525,tyr572,arg380,arg415、phe577、asn382和ser555关键氨基酸结合的氢键、π-π相互作用和疏水相互作用可能有助于肽与keap1的结合能力,可以通过参与keap1-nrf2信号通路来促进体内抗氧化活性。这些新型富硒多肽具有作为天然抗氧化剂在功能食品中的应用前景。
[0114]
最后应当指出的是,以上实施例仅是本发明的具有代表性的例子。显然,本发明的技术方案并不限于上述实施例,还可有许多变形。本领域的普通技术人员能从本发明内容直接导出或联想到所有变形,均应认为是本发明的权利要求的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1