从气态底物发酵生产2-苯乙醇的制作方法
从气态底物发酵生产2
‑
苯乙醇
1.相关申请的交叉引用
2.本技术要求于2020年3月18日提交的美国临时专利申请第62/991,428号的权益,所述美国临时专利申请的全部内容通过引用并入本文。
3.政府权利
4.本发明是根据美国能源部授予的合作协议de
‑
ee0007728在政府支持下进行的。
技术领域
5.本技术涉及用于通过对包括二氧化碳(co2)、一氧化碳(co)和/或氢气(h2)的底物进行微生物发酵来生产2
‑
苯乙醇的方法以及用于在此类方法中使用的经基因修饰的微生物。
背景技术:
6.缓解迫切的气候变化需要大幅减少温室气体(ghg)的排放,所述ghg如通过煤和石油等化石燃料的燃烧生成的那些。虽然化学品和运输燃料的可持续来源目前不足以显著取代对化石碳的依赖性,但是气体发酵最近已成为用于将如co2、co和/或h2等气体生物固定成可持续燃料和化学品的替代性平台。具体地,气体发酵技术可以利用包含气化的有机物(例如,市政固体废物或农业废物)或工业废气(例如,来自钢厂或炼油厂)在内的广泛范围的原料来产生乙醇、喷气燃料和各种其它产品。仅靠气体发酵就可以取代30%的原油使用量并使全球co2排放量减少10%,但是,与任何破坏性技术一样,在充分发挥这一潜力之前,必须克服许多技术挑战。
[0007]2‑
苯乙醇(2
‑
pe)是商用化学品,以其“似玫瑰的”芳香在食品、香精和香料行业中具有6010mt/yr的市场并且在化工行业中作为酯中间体并且理论上讲在苯乙烯生产中具有900mt/yr的市场。当前,2
‑
pe是从天然来源取得的以应用于食品、香精和香料行业中,或者其通过化学手段合成以用于其它应用。因此,需要减轻对天然和石化过程的依赖性的改进的2
‑
pe生产方法。
技术实现要素:
[0008]
在一方面,本公开提供了一种能够产生2
‑
苯乙醇的微生物,其中所述微生物包括将苯丙酮酸盐转化为苯乙醛的异源酶和将苯乙醛转化为2
‑
苯乙醇的异源酶。
[0009]
在一些实施例中,所述将苯丙酮酸盐转化为苯乙醛的异源酶为脱羧酶,并且所述将苯乙醛转化为2
‑
苯乙醇的异源酶为苯乙醛还原酶。具体地,所述脱羧酶可以为苯丙酮酸盐特异性脱羧酶。
[0010]
在一些实施例中,所述微生物为c1固定型微生物、伍德
‑
隆达尔微生物(wood
‑
ljungdahl microorganism)和/或细菌。在一些实施例中,所述微生物是以下属的成员:醋酸杆菌属(acetobacterium)、碱杆菌属(alkalibaculum)、布劳特氏菌属(blautia)、丁酸杆菌属(butyribacterium)、梭菌属(clostridium)、真杆菌属(eubacterium)、穆尔氏菌属
(moorella)、产醋杆菌属(oxobacter)、鼠孢菌属(sporomusa)或嗜热厌氧杆菌属(thermoanaerobacter)。
[0011]
在一些实施例中,所述微生物天然地能够产生苯丙酮酸盐。在一些实施例中,所述微生物包括将co、co2和/或h2转化为乙酰辅酶a的伍德
‑
隆达尔途径。
[0012]
在一些实施例中,所述微生物包括以下中的一种或多种:将乙酰辅酶a转化为丙酮酸盐的天然酶;将丙酮酸盐转化为磷酸烯醇丙酮酸盐的天然酶;将磷酸烯醇丙酮酸盐和赤藓糖
‑4‑
磷酸盐转化为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐的天然酶;将2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐转化为3
‑
脱氢奎尼酸盐的天然酶;将3
‑
脱氢奎尼酸盐转化为3
‑
脱氢莽草酸盐的天然酶;将3
‑
脱氢莽草酸盐转化为莽草酸盐的天然酶;将莽草酸盐转化为莽草酸3
‑
磷酸盐的天然酶;将莽草酸3
‑
磷酸盐转化为5
‑
o
‑
(1
‑
羧基乙烯基)
‑
莽草酸3
‑
磷酸盐的天然酶;将5
‑
o
‑
(1
‑
羧基乙烯基)
‑
莽草酸3
‑
磷酸盐转化为分支酸盐的天然酶;或将分支酸盐转化为苯丙酮酸盐的天然酶。
[0013]
在一些实施例中,所述将乙酰辅酶a转化为丙酮酸盐的天然酶为丙酮酸盐:铁氧还蛋白氧化还原酶(pfor;e.c.1.2.7.1);所述将丙酮酸盐转化为磷酸烯醇丙酮酸盐的天然酶为丙酮酸磷酸双激酶(ppdk;e.c.2.7.9.1);所述将磷酸烯醇丙酮酸盐和赤藓糖
‑4‑
磷酸盐转化为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐的天然酶为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸合酶(dahps或dahp合酶;e.c.2.5.1.54);所述将2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐转化为3
‑
脱氢奎尼酸盐的天然酶为3
‑
脱氢奎尼酸合酶(dhqs;e.c.4.2.3.4);所述将3
‑
脱氢奎尼酸盐转化为3
‑
脱氢莽草酸盐的天然酶为3
‑
脱氢奎尼酸脱水酶(dhq;e.c.4.2.1.10);所述将3
‑
脱氢莽草酸盐转化为莽草酸盐的天然酶为莽草酸脱氢酶(sdh;e.c.1.1.1.25);所述将莽草酸盐转化为莽草酸3
‑
磷酸盐的天然酶为莽草酸激酶(sk;e.c.2.7.1.71);所述将莽草酸3
‑
磷酸盐转化为5
‑
o
‑
(1
‑
羧基乙烯基)
‑
莽草酸3
‑
磷酸盐的天然酶为5
‑
烯醇丙酮酸莽草酸3
‑
磷酸合酶(epsps或epsp合酶;e.c.2.5.1.19);所述将5
‑
o
‑
(1
‑
羧基乙烯基)
‑
莽草酸3
‑
磷酸盐转化为分支酸盐的天然酶为分支酸合酶(cs;e.c.4.2.3.5);或所述将分支酸盐转化为苯丙酮酸盐的天然酶为双官能分支酸变位酶(e.c.5.4.99.5)/预苯酸脱水酶(e.c.4.2.1.51)。
[0014]
在一些实施例中,所述微生物进一步包括以下中的一种或多种:将磷酸烯醇丙酮酸盐和赤藓糖
‑4‑
磷酸盐转化为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐的异源酶;将3
‑
脱氢莽草酸盐转化为莽草酸盐的异源酶;或将分支酸盐转化为苯丙酮酸盐的异源酶。
[0015]
在一些实施例中,所述将磷酸烯醇丙酮酸盐和赤藓糖
‑4‑
磷酸盐转化为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐的异源酶为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸合酶;所述将3
‑
脱氢莽草酸盐转化为莽草酸盐的异源酶为莽草酸脱氢酶;或所述将分支酸盐转化为苯丙酮酸盐的异源酶为双官能分支酸变位酶/预苯酸脱水酶。
[0016]
在一些实施例中,所述微生物包括:
[0017]
(1)ecaroe、kivd、lepar、phea2和arog;
[0018]
(2)ecaroe、abppdc、ecpar、phea1和arog;
[0019]
(3)cgaroe、abppdc、lepar、phea2和arog;
[0020]
(4)ecaroe、abppdc、lepar、phea2和arog;
[0021]
(5)cgaroe、kivd、rrpar、phea2和arog;
[0022]
(6)ecaroe、abppdc、lepar、phea2和arog;
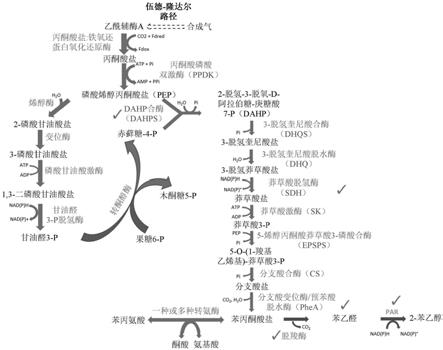
[0023]
(7)phea1、arog、abppdc和ecpar;
[0024]
(8)egaroe、abppdc、lepar、phea1和arog;
[0025]
(9)ecaroe、abppdc、ecpar、phea2和arog;
[0026]
(10)cgaroe、aro10、ecpar、phea2和arog;
[0027]
(11)cgaroe、abppdc、lepar、phea1和arog;
[0028]
(12)cgaroe、aro10、lepar、phea2和arog;
[0029]
(13)ecaroe、abppdc、lepar、phea1和arog;
[0030]
(14)ecaroe、aro10、ecpar、phea2和arog;
[0031]
(15)ecaroe、aro10、lepar、phea2和arog;
[0032]
(16)cgaroe、abppdc、lepar、phea1和arog;
[0033]
(17)cgaroe、aro10、rspar、phea2和arog;
[0034]
(18)cgaroe、abppdc、blpar、phea2和arog;
[0035]
(19)ecaroe、kivd、blpar、phea2和arog;
[0036]
(20)cgaroe、aro10、blpar、phea2和arog;
[0037]
(21)ecaroe、abppdc、blpar、phea2和arog;
[0038]
(22)ecaroe、abppdc、adh6、phea1和arog;
[0039]
(23)cgaroe、abppdc、adh6、phea2和arog;
[0040]
(24)cgaroe、abppdc、ecpar、phea2和arog;
[0041]
(25)ecaroe、aro10、rspar、phea2和arog;
[0042]
(26)ecaroe、aro10、adh6、phea2和arog;或
[0043]
(27)ecaroe、abppdc、ecpar、phea2和arog。
[0044]
在一些实施例中,所述微生物包括组合菌株2c03、1c10、2c54、2c09、2c47、2c55、2c53、h57、1c47、2c10、2a18、1c54、2c01、1c31、2b17、2c52、1c09、h18、h58、2c27、2d22、2d07、2d24、2d03、1c29、2b26、2c38、2a27、2c12或2b27的异源基因和启动子。
[0045]
在一些实施例中,所述微生物发酵包括co、co2和/或h2的气态底物以产生2
‑
苯乙醇。在一些实施例中,所述气态底物包括合成气或工业废气。
[0046]
在一些实施例中,所述微生物不产生任何其它c3+醇。在一些实施例中,所述微生物不产生任何其它c3、c4、c5、c6、c7、c8、c9或c10醇。
[0047]
在另一方面,本公开提供了一种生产2
‑
苯乙醇的方法,所述方法包括在气态底物存在的情况下培养所述微生物。在一些实施例中,所述气态底物包括c1碳源,所述c1碳源包括co、co2和/或h2。在一些实施例中,所述气态底物包括合成气或工业废气。
[0048]
根据对某些实施例的以下更详细的描述和权利要求,本公开的具体实施例将变得显而易见。
附图说明
[0049]
图1示出了导致由产乙醇梭菌(clostridium autoethanogenum)从气体发酵生物合成2
‑
苯乙醇的代谢途径。需要异源表达的酶促步骤是脱羧酶步骤和苯乙醛还原酶(par)步骤。虚线箭头描绘了产乙醇梭菌中的多酶促核心代谢途径。碳和氢通过伍德
‑
隆达尔途径
同化为乙酰辅酶a,所述乙酰辅酶a然后通过丙酮酸盐:铁氧还蛋白氧化还原酶转化为丙酮酸盐。芳香族氨基酸的生物合成起始于磷酸烯醇丙酮酸盐和赤藓糖
‑4‑
磷酸盐,所述磷酸烯醇丙酮酸盐和赤鲜糖
‑4‑
磷酸盐是由丙酮酸盐通过基础代谢途径,如糖异生和磷酸戊糖途径生物合成的。2
‑
苯丙酮酸盐可以通过苯丙氨酸与酮酸之间的转氨酶反应从莽草酸盐途径得到。在本文所公开的实例中异源表达或过表达的酶用勾号标记标记出。nad(p)h包括nadph和nadh。
[0050]
图2a
‑
2b示出了在肖特瓶(schott bottle)中存在不同量的2
‑
pe的情况下产乙醇梭菌的自养生长。图2a:生长曲线;图2b:通过gc
‑
fid测量的2
‑
pe浓度。unc=未经攻击;n=2;误差条=s.d.
[0051]
图3a示出了生长曲线,并且图3b示出了在以下三个脱羧酶菌株之间在肖特瓶中使用150kpa合成气共混物(50%co、10%h2、30%co2和10%n2)自养生长期间的终点醇产生滴度:abppdc、aro10和对照kivd_la。n=3;误差条=s.d.
[0052]
图4a
‑
4c示出了用于2
‑
pe生产的产乙醇梭菌中的苯乙醛还原酶(par)的过表达。图4a:物种起源、氨基酸长度、辅因子特异性和所采用par的参考文献。图4b:使用氨基酸序列的par变体的系统发生树。比例尺对应于每个氨基酸0.2变化。图4c:在200kpa合成气共混物(50%co、10%h2、30%co2和10%n2)存在的情况下在温育7天之后具有不同par变体的菌株在12孔板中的归一化2
‑
pe产量;n=3
‑
5;误差条=s.d.
[0053]
图5示出了使用金门方法(golden gate method)的2
‑
pe途径基因的组合装配。
[0054]
图6示出了用于实例4中所描述的两个2
‑
pe组合文库的基因和启动子变体。
[0055]
图7a
‑
c示出了使用200kpa合成气共混物(50%co、10%h2、30%co2和10%n2)在12孔板中的162个2
‑
pe组合菌株的表征。图7a:相对于对照菌株sfa212的2
‑
pe滴度;图7b:通过生物质浓度归一化的2
‑
pe滴度;图7c:生物质浓度;温育持续时间=8
‑
10天;n=2;误差条=s.d.
[0056]
图8a
‑
e示出了根据以下对组合菌株中的终点2
‑
pe滴度的比较:质粒的数量(图8a);双官能分支酸变位酶/预苯酸脱水酶变体(图8b);脱羧酶变体(图8c);莽草酸脱氢酶变体(图8d);以及苯乙醛还原酶变体(图8e)。温育持续时间=8
‑
10天;误差条=s.d.
[0057]
图9a
‑
b示出了12个组合菌株在使用合成气共混物(40%co、20%h2、20%co2和20%n2)的连续cstr发酵中的表征。相对于参考菌株sfa212,在菌株1c29中观察到2
‑
pe滴度增加2.9倍。与参考菌株相比,10个菌株实现了更高的2
‑
pe产率。
[0058]
图10a
‑
b示出了组合菌株1c29在具有氮限制的两个连续cstr试验中的2
‑
pe产生。
[0059]
图11a
‑
c示出了使用菌株sfa212测试经预处理的衍生自玉米秸秆的合成气(23.3%h2、38.7%co、19.6%co2和18.4%n2)在连续cstr发酵中的作用的结果。图11a:通过hplc分析的代谢物曲线;图11b:通过gc
‑
tcd分析的气体曲线(负值=摄取);图11c:通过分析的gc
‑
fid的2
‑
pe曲线。在第6.9天与第10.9天之间(总共4天)使用了实际合成气。
具体实施方式
[0060]
本文公开了用于由合成气从头生物合成2
‑
苯乙醇(2
‑
pe)的方法和组合物,所述方法和组合物具有减轻对用于制造2
‑
pe的天然和石化过程的依赖性的潜力。
[0061]
定义
[0062]
除非另有定义,否则如贯穿本说明书使用的以下术语定义如下:
[0063]
术语“发酵”应解释为在底物中产生化学变化的代谢过程。例如,发酵过程接收一种或多种底物并通过利用一种或多种微生物来产生一种或多种产物。术语“发酵”、“气体发酵”等应解释为接收一种或多种底物如通过气化产生的合成气并通过利用一种或多种c1固定型微生物来产生一种或多种产物的过程。优选地,发酵过程包含使用一种或多种生物反应器。发酵过程可以描述为“分批”或“连续”。“分批发酵”用于描述这样的发酵过程:其中生物反应器填充有原材料例如碳源以及微生物,其中产物在生物反应器中保留到直到发酵完成。在“分批”过程中,在发酵完成之后,提取产物并在下一“分批”开始之前清洁生物反应器。“连续发酵”用于描述这样的发酵过程:其中发酵过程持续较长时间段,并且产物和/或代谢物在发酵期间提取。优选地,发酵过程是连续的。
[0064]
当用于指微生物时,术语“非天然存在的”旨在意指微生物具有在所参考物种的天然存在的菌株(包含所参考物种的野生型菌株)中未发现的至少一种基因修饰。非天然存在的微生物通常在实验室或研究机构中开发。
[0065]
术语“基因修饰”、“基因改变”或“基因工程”广义上指通过人工对微生物的基因组或核酸的操作。同样,术语“经基因修饰的”、“经基因改变的”或“经基因工程化的”是指含有这种基因修饰、基因改变或基因工程的微生物。这些术语可以用于区分实验室生成的微生物与天然存在的微生物。基因修饰的方法包含例如异源基因表达、基因或启动子插入或缺失、核酸突变、经改变的基因表达或失活、酶工程、定向进化、基于知识的设计、随机诱变方法、基因改组和密码子优化。
[0066]
如梭菌(clostridia)等微生物的代谢工程可以极大地扩展其产生除天然代谢物如乙醇以外的许多重要燃料和化学分子的能力。近年来,已经开发了用于针对梭菌进行基因组工程的若干种不同方法,包含基于内含子的方法(clostron)(kuehne,《菌株工程:方法和方案(strain eng:methods and protocols)》,389
‑
407,2011)、等位基因交换法(ace)(heap,《核酸研究(nucl acids res)》,40:e59,2012;ng,《公共科学图书馆:综合(plos one)》,8:e56051,2013)、三重杂交(liew,《微生物前沿(frontiers microbiol)》,7:694,2016)、通过i
‑
scei介导的方法(zhang,《微生物学方法杂志(journal microbiol methods),108:49
‑
60,2015)、mazf(al
‑
hinai,《应用及环境微生物学(appl environ microbiol),78:8112
‑
8121,2012)或其它方法(argyros,《应用及环境微生物学,77:8288
‑
8294,2011)、cre
‑
lox(ueki,《mbio》,5:e01636
‑
01614,2014)以及crispr/cas9(nagaraju,《生物燃料生物技术(biotechnol biofuels)》,9:219,2016)。然而,由于缓慢而费力的循环时间以及这些基因技术跨物种的可转移性的限制,以迭代方式引入多于几个基因改变仍然极具挑战性。此外,尚未充分了解梭菌中的c1代谢,以至于无法可靠地预测将使趋于产物合成的c1摄取、转化和碳/能量/氧化还原流最大化的修饰。因此,在梭菌中引入靶途径仍然是繁琐且耗时的过程。
[0067]“重组”指示核酸、蛋白质或微生物是基因修饰、基因工程或基因重组的产物。一般而言,术语“重组”是指含有源自多个来源如两种或更多种不同的微生物菌株或物种的基因材料或由其编码的核酸、蛋白质或微生物。
[0068]“野生型”是指区别于突变形式或变体形式的在其存在于自然中时的有机体、菌株、基因或特性的典型形式。
[0069]“内源”或“天然”是指在衍生出本公开的微生物的野生型或亲代微生物中存在或表达的核酸或蛋白质。例如,内源基因是天然存在于衍生出本公开的微生物的野生型或亲代微生物中的基因。在一个实施例中,内源基因的表达可以由外源调控元件如外源启动子控制。
[0070]“外源”是指起源于本公开的微生物之外的核酸或蛋白质。例如,可以人工或重组产生外源基因或酶并将其引入本公开的微生物中或在本公开的微生物中表达。也可以从异源微生物中分离外源基因或酶并将其引入本公开的微生物中或在本公开的微生物中表达。外源核酸可以被改编以整合到本公开的微生物的基因组中或在本公开的微生物中例如在质粒中保持染色体外状态。
[0071]“异源”是指在衍生出本公开的微生物的野生型或亲代微生物中不存在的核酸或蛋白质。例如,异源基因或酶可以衍生自不同的菌株或物种并被引入本公开的微生物中或在本公开的微生物中表达。异源基因或酶可以以其存在于不同菌株或物种中的形式被引入本公开的微生物或在本公开的微生物中表达。替代性地,可以以某种方式修饰异源基因或酶,例如,通过对其进行密码子优化以用于在本公开的微生物中表达或通过对其进行工程化以改变功能如以逆转酶活性方向或改变底物特异性。
[0072]
术语“多核苷酸”、“核苷酸”、“核苷酸序列”、“核酸”和“寡核苷酸”可互换使用。所述术语指任何长度的核苷酸(脱氧核糖核苷酸或核糖核苷酸)或其类似物的聚合形式。多核苷酸可以具有任何三维结构并且可以进行任何已知或未知的功能。以下是多核苷酸的非限制性实例:基因或基因片段的编码或非编码区、由连锁分析定义的基因座(loci)(基因座(locus))、外显子、内含子、信使rna(mrna)、转移rna、核糖体rna、短干扰rna(sirna)、短发夹rna(shrna)、微rna(mirna)、核酶、cdna、重组多核苷酸、分支多核苷酸、质粒、载体、任何序列的分离的dna、任何序列的分离的rna、核酸探针和引物。多核苷酸可以包括一个或多个经修饰的核苷酸,如甲基化核苷酸或核苷酸类似物。如果存在,则对核苷酸结构的修饰可以在组装聚合物之前或之后进行。核苷酸序列可以被非核苷酸组分中断。多核苷酸可以在聚合后如通过与标记组分缀合来进一步修饰。
[0073]
如本文所使用的,“表达”是指多核苷酸从dna模板转录的过程(如转录为mrna或其它rna转录物)和/或经转录mrna随后被翻译成肽、多肽或蛋白质的过程。转录物和编码的多肽可以统称为“基因产物”。
[0074]
术语“多肽”、“肽”和“蛋白质”在本文中可互换使用以指任何长度的氨基酸的聚合物。聚合物可以是直链或支链的,其可以包括经修饰氨基酸,并且其可以被非氨基酸中断。术语还涵盖已被修饰的氨基酸聚合物;例如,二硫键形成、糖基化、脂化、乙酰化、磷酸化或任何其它操作,如与标记组分缀合。如本文所使用的,术语“氨基酸”包含天然和/或非天然或合成的氨基酸,包含甘氨酸和d或l光学异构体两者以及氨基酸类似物和肽模拟物。
[0075]“酶活性”或简称为“活性”广义上是指酶促活性,包含但不限于酶的活性、酶的量或酶催化反应的可用性。因此,“增加”酶活性包含增加酶的活性、增加酶的量或增加酶催化反应的可用性。类似地,“降低”酶活性包含降低酶的活性、降低酶的量或降低酶催化反应的可用性。
[0076]“密码子优化”是指用于在特定菌株或物种中对核酸进行优化的或改进的翻译的核酸如基因突变。密码子优化可以产生更快的翻译速率或更高的翻译准确性。
[0077]“过表达”是指与衍生出本公开的微生物的野生型或亲代微生物相比本公开的微生物中的核酸或蛋白质的表达增加。过表达可以通过本领域已知的任何手段实现,包含修饰基因拷贝数、基因转录速率、基因翻译速率或酶降解速率。
[0078]
术语“变体”包含其序列不同于参考核酸和蛋白质的序列如现有技术中公开的或本文例举的参考核酸和蛋白质的序列的核酸和蛋白质。本公开的方法和组合物可以使用进行与参考核酸或蛋白质基本上相同的功能的变体核酸或蛋白质实践。例如,变体蛋白质可以进行与参考蛋白质基本上相同的功能或催化与参考蛋白质基本上相同的反应。变体基因可以编码与参考基因相同或基本上相同的蛋白质。变体启动子可以具有与参考启动子基本上相同的能力来促进一种或多种基因的表达。
[0079]
此类核酸或蛋白质在本文中可以被称为“功能等效变体”或“功能同系物”。举例来说,核酸的功能等效变体可以包含等位基因变体、基因片段、突变基因、多态性等。来自其它微生物的同源基因也是功能等效变体的实例。这些同源基因包含如丙酮丁醇梭菌(clostridium acetobutylicum)、拜氏梭菌(clostridium beijerinckii)或扬氏梭菌(clostridium ljungdahlii)等物种中的同源基因,其详细信息可在如genbank或ncbi等网站上公开获得。功能等效变体还包含其序列由于特定微生物的密码子优化而变化的核酸。核酸的功能等效变体将与参考的核酸优选地具有至少大约70%、大约80%、大约85%、大约90%、大约95%、大约98%或更高的核酸序列同一性(同源性百分比)。蛋白质的功能等效变体(即,功能同系物)将与参考的蛋白质优选地具有至少大约70%、大约80%、大约85%、大约90%、大约95%、大约98%或更高的氨基酸同一性(同源性百分比)。可以使用本领域已知的任何方法评估变体核酸或蛋白质的功能等效性。
[0080]
核酸或氨基酸序列同一性要求两个比对序列之间具有相同的氨基酸序列。因此,与参考序列共享80%的核酸或氨基酸同一性的候选序列要求在比对后,候选序列中80%的核酸或氨基酸与参考序列中对应的核酸或氨基酸相同。根据本发明的同一性是借助于计算机分析以及其中建议的默认参数确定的,所述计算机分析如但不限于clustalw计算机比对程序(higgins等人,《核酸研究》22:4673
‑
4680)。使用例如clustalw软件与其默认设置,将查询的成熟(生物活性)部分与参考多肽进行比对。对完全保守残基的数目进行计数并除以参考多肽的长度。
[0081]
可以使用本领域已知的任何方法将核酸递送到本公开的微生物。例如,核酸可以作为裸核酸递送或者可以用一种或多种试剂如脂质体配制。适当时,核酸可以是dna、rna、cdna或其组合。在某些实施例中可以使用限制性抑制剂。另外的载体可以包含质粒、病毒、噬菌体、粘粒和人工染色体。在优选实施例中,使用质粒将核酸递送到本公开的微生物。举例来说,转化(包含转导或转染)可以通过电穿孔、超声波处理、聚乙二醇介导的转化、化学或天然感受态、原生质体转化、前噬菌体诱导或缀合来实现。在具有活性限制酶系统的某些实施例中,可能会有必要在将核酸引入到微生物中之前使核酸甲基化。
[0082]
此外,可以将核酸设计成包括调控元件如启动子以增加或以其它方式控制特定核酸的表达。启动子可以是组成型启动子或诱导型启动子。理想情况下,启动子是伍德
‑
隆达尔途径启动子、铁氧还蛋白启动子、丙酮酸盐:铁氧还蛋白氧化还原酶启动子、rnf复合操纵子启动子、atp合酶操纵子启动子或磷酸转乙酰酶/乙酸激酶操纵子启动子。
[0083]“微生物”是微观有机体,具体地是细菌、古菌、病毒或真菌。本公开的微生物通常
是细菌。如本文所使用的,对“微生物”的叙述应被视为涵盖“细菌”。
[0084]“亲代微生物”是用于生成本公开的微生物的微生物。亲代微生物可以是天然存在的微生物(即,野生型微生物)或先前已经被修饰的微生物(即,突变或重组微生物)。本公开的微生物可以被修饰成表达或过表达未在亲代微生物中表达或过表达的一种或多种酶。类似地,本公开的微生物可以被修饰成含有亲代微生物所不含的一个或多个基因。本公开的微生物还可以被修饰成不表达或表达在亲代微生物中表达的较低量的一种或多种酶。在一个实施例中,亲代微生物是产乙醇梭菌、扬氏梭菌或拉氏梭菌(clostridium ragsdalei)。在优选实施例中,亲代微生物是产乙醇梭菌lz1561,其根据《布达佩斯条约(budapest treaty)》的条款于2010年6月7日由位于德国布伦瑞克省d
‑
38124 inhoffenstraβe 7b(inhoffenstraβe 7b,d
‑
38124braunschweig,germany)的德国微生物菌种保藏中心(deutsche sammlung von mikroorganismen und zellkulturen gmbh,dsmz)保藏在一致登录号dsm23693下。此菌株在国际专利申请第pct/nz2011/000144号中进行了描述,所述国际专利申请以wo 2012/015317公开。
[0085]
术语“衍生自”指示核酸、蛋白质或微生物从不同的(例如,亲代或野生型)核酸、蛋白质或微生物修饰或改编,以产生新的核酸、蛋白质或微生物。此类修饰或改编通常包含核酸或基因的插入、缺失、突变或取代。通常,本公开的微生物衍生自亲代微生物。在一个实施例中,本公开的微生物衍生自产乙醇梭菌、扬氏梭菌或拉氏梭菌。在优选实施例中,本公开的微生物衍生自产乙醇梭菌lz1561,其保藏在dsmz登录号dsm23693下。
[0086]
本公开的微生物可以基于功能特性进一步分类。例如,本公开的微生物可以是或可以衍生自c1固定型微生物、厌氧菌、产乙酸菌、产乙醇菌(ethanologen)、一氧化碳营养菌(carboxydotroph)和/或甲烷氧化菌。表1提供了微生物的代表性列表并标识了其功能特性。
[0087][0088]1伍氏醋酸杆菌可以从果糖中产生乙醇,但不能从气体中产生乙醇。
[0089]2尚未研究马氏梭菌是否可以依靠co生长。
[0090]3已报告,热醋穆尔氏菌中的一种菌株穆尔氏菌huc22
‑
1从气体中产生乙醇。
[0091]4尚未研究卵形鼠孢菌是否可以依靠co生长。
[0092]5尚未研究森林土壤醋酸鼠孢菌是否可以依靠co生长。
[0093]6尚未研究球形鼠孢菌是否可以依靠co生长。
[0094]“伍德
‑
隆达尔”是指碳固定的伍德
‑
隆达尔途径,如例如由ragsdale,《生物化学与生物物理学报(biochim biophys acta)》,1784:1873
‑
1898,2008所描述的。“伍德
‑
隆达尔微生物”可预见地指代含有伍德
‑
隆达尔途径的微生物。一般来说,本公开的微生物含有天然伍德
‑
隆达尔途径。在本文中,伍德
‑
隆达尔途径可以是天然的未经修饰的伍德
‑
隆达尔途径,或者所述伍德
‑
隆达尔途径可以是有一定程度的基因修饰(例如,过表达、异源表达、敲除等)的伍德
‑
隆达尔途径,只要所述伍德
‑
隆达尔途径仍然起作用以将co、co2和/或h2转化为乙酰辅酶a即可。
[0095]“c1”是指单碳分子,例如co、co2、ch4或ch3oh。“c1氧化物”是指也包括至少一个氧原子的单碳分子,例如co、co2或ch3oh。“c1碳源”是指用作本公开的微生物的部分或唯一碳源的单碳分子。例如,c1碳源可以包括co、co2、ch4、ch3oh或ch2o2中的一种或多种。优选地,c1碳源包括co和co2中的一种或两种。“c1固定型微生物”是能够从c1碳源产生一种或多种产物的微生物。本公开的微生物通常是c1固定型细菌。在优选实施例中,本公开的微生物衍生自表1中标识的c1固定型微生物。
[0096]“厌氧菌”是不需要氧气即可生长的微生物。如果氧气超过某一阈值存在,则厌氧菌可能会产生不良反应或甚至死亡。然而,一些厌氧菌能够耐受低水平的氧气(例如,0.000001
‑
5%的氧气)。本公开的微生物通常是厌氧菌。在一些实施例中,本公开的微生物衍生自表1中标识的厌氧菌。
[0097]“产乙酸菌”是使用伍德
‑
隆达尔途径作为其进行能量保存和合成乙酰辅酶a和乙酰辅酶a衍生的产物如乙酸盐的主要机制的专性厌氧细菌(ragsdale,《生物化学与生物物理学报》,1784:1873
‑
1898,2008)。具体地,产乙酸菌使用伍德
‑
隆达尔途径作为(1)用于从co2还原合成乙酰辅酶a的机制、(2)末端电子接受能量保存过程、(3)用于在细胞碳的合成中固定(同化)co2的机制(drake,产乙酸菌原核生物(acetogenic prokaryotes),见于《原核生物(the prokaryotes)》,第3版,第354页,纽约,纽约州,2006)。所有天然存在的产乙酸菌都是c1固定型、厌氧型、自养型和非甲烷氧化型。本公开的微生物通常是产乙酸菌。在优选实施例中,本公开的微生物衍生自表1中标识的产乙酸菌。
[0098]“产乙醇菌”是产生或能够产生乙醇的微生物。本公开的微生物通常是产乙醇菌。在优选实施例中,本公开的微生物衍生自表1中标识的产乙醇菌。
[0099]“自养菌”是能够在不存在有机碳的情况下生长的微生物。相反,自养菌使用无机碳源,如co和/或co2。本公开的微生物通常是自养菌。在优选实施例中,本公开的微生物衍生自表1中标识的自养菌。
[0100]“一氧化碳营养菌”是能够利用co作为碳和能量的唯一来源的微生物。本公开的微生物通常是一氧化碳营养菌。在优选实施例中,本公开的微生物衍生自表1中标识的一氧化碳营养菌。
[0101]“甲烷氧化菌”是能够利用甲烷作为碳和能量的唯一来源的微生物。在某些实施例中,本公开的微生物是甲烷氧化菌或衍生自甲烷氧化菌。在其它实施例中,本公开的微生物不是甲烷氧化菌或不衍生自甲烷氧化菌。
[0102]
更广泛地说,本公开的微生物可以衍生自表1中标识的任何属或种。例如,微生物可以是选自由以下组成的组的属的成员:醋酸杆菌属、碱杆菌属、布劳特氏菌属、丁酸杆菌属、梭菌属、真杆菌属、穆尔氏菌属、产醋杆菌属、鼠孢菌属和嗜热厌氧杆菌属。具体地,微生
物可以衍生自选自由以下组成的组的亲代细菌:伍氏醋酸杆菌、巴氏碱杆菌、长生布劳特氏菌、食甲基丁酸杆菌、醋酸梭菌、产乙醇梭菌、食一氧化碳梭菌、科斯卡塔梭菌、德氏梭菌、蚁酸醋酸梭菌、扬氏梭菌、马氏梭菌、拉氏梭菌、粪味梭菌、粘液真杆菌、热自养穆尔氏菌、热醋穆尔氏菌、普氏产醋杆菌、卵形鼠孢菌、森林土壤醋酸鼠孢菌、球形鼠孢菌和凯伍嗜热厌氧菌。
[0103]
已知这些类型的微生物在生命的热力学极限内生存(schuchmann,《自然综述:微生物学(nat rev microbiol)》,12:809
‑
821,2014),因此,在本发明之前,对于这些类型的微生物是否将能够合成c8芳香族醇一无所知,更别提以有意义的数量合成,尤其是考虑到2
‑
pe对这些微生物具有毒性作用的潜力。
[0104]
在优选实施例中,本公开的微生物衍生自包括物种产乙醇梭菌、扬氏梭菌和拉氏梭菌的梭菌属簇。这些物种最初由以下报告和表征:abrini,《微生物学文献集(arch microbiol)》,161:345
‑
351,1994(产乙醇梭菌);tanner,《国际系统细菌学杂志(int j system bacteriol)》,43:232
‑
236,1993(扬氏梭菌);以及huhnke,wo 2008/028055(拉氏梭菌)。
[0105]
这三个物种有许多相似之处。具体地,这些物种都是梭菌属的c1固定型、厌氧型、产乙酸型、产乙醇型和一氧化碳营养型成员。这些物种具有类似的基因型和表型以及类似的能量保存和发酵代谢模式。此外,这些物种聚集于16s rrna dna超过99%相同的梭菌rrna同源性组i中、具有约22
‑
30mol%的dna g+c含量、是革兰氏阳性的、具有类似的形态和大小(对数生长期细胞介于0.5
‑
0.7
×3‑
5μm之间)、是嗜温性的(在30
‑
37℃下生长最佳)、具有约4
‑
7.5的类似ph范围(最佳ph为约5.5
‑
6)、缺乏细胞色素并且通过rnf复合体保存能量。此外,在这些物种中已示出了羧酸到其对应的醇的还原(perez,《生物技术与生物工程(biotechnol bioeng)》,110:1066
‑
1077,2012)。重要的是,这些物种还都示出在含co气体上的强自养性生长、产生乙醇和乙酸盐(或乙酸)作为主要发酵产物并且在某些条件下产生少量的2,3
‑
丁二醇和乳酸。
[0106]
然而,这三种物种也有许多不同之处。这些物种分离自不同的来源:来自兔子肠道的产乙醇梭菌、来自鸡圈废物的扬氏梭菌以及来自淡水沉积物的拉氏梭菌。这些物种在各种糖(例如,鼠李糖、阿拉伯糖)、酸(例如,葡糖酸盐、柠檬酸盐)、氨基酸(例如,精氨酸、组氨酸)和其它底物(例如,甜菜碱、丁醇)的利用方面有所不同。此外,这些物种在针对某些维生素(例如,硫胺素、生物素)的营养缺陷型方面有所不同。虽然已经发现这些基因和蛋白质的一般结构和数量在所有物种中都是相同的,但是这些物种在伍德
‑
隆达尔途径基因和蛋白质的核酸和氨基酸序列方面存在差异(《生物技术最新观点(curr opin biotechnol)》,22:320
‑
325,2011)。
[0107]
因此,总的来说,产乙醇梭菌、扬氏梭菌或拉氏梭菌的特性中的许多特性并不对所述物种具有特异性,而是梭菌属的c1固定型、厌氧型、产乙酸型、产乙醇型和一氧化碳营养型成员的这一簇的一般特性。然而,由于这些物种实际上是不同的,因此这些物种中的一种物种的基因修饰或操作在这些物种的另一种物种中可能不具有相同的作用。例如,可以观察到生长、性能或产物产生的不同。
[0108]“底物”是指本公开的微生物的碳源和/或能量来源。底物通常呈气态并且包括c1碳源,例如co、co2和/或ch4。优选地,底物包括co或co+co2的c1碳源。底物可以进一步包括其
它非碳组分,如h2、n2或电子。
[0109]
底物通常包括至少一定量的co,如约1、2、5、10、20、30、40、50、60、70、80、90或100mol%的co。底物可以包括一定范围的co,如约20
‑
80mol%、30
‑
70mol%或40
‑
60mol%的co。优选地,底物包括约40
‑
70mol%的co(例如,钢厂或高炉气)、约20
‑
30mol%的co(例如,碱性氧气炉气)或约15
‑
45mol%的co(例如,合成气)。在一些实施例中,底物可以包括相对较低量的co,如约1
‑
10mol%或1
‑
20mol%的co。本公开的微生物通常将底物中的co的至少一部分转化为产物。在一些实施例中,底物不包括或基本上不包括(<1mol%)co。
[0110]
底物可以包括一定量的h2。例如,底物可以包括约1、2、5、10、15、20或30mol%的h2。在一些实施例中,底物可以包括相对较高量的h2,如约60、70、80或90mol%的h2。在另外的实施例中,底物不包括或基本上不包括(<1mol%)h2。
[0111]
底物可以包括一定量的co2。例如,底物可以包括约1
‑
80mol%或1
‑
30mol%的co2。在一些实施例中,底物可以包括小于约20、15、10或5mol%的co2。在另一个实施例中,底物不包括或基本上不包括(<1mol%)co2。
[0112]
尽管底物通常是气态的,但底物也可以以替代形式提供。举例来说,可以使用微泡分散发生器将底物溶解在用含co气体饱和的液体中。通过另外的实例,底物可以吸附到固体载体上。
[0113]
底物和/或c1碳源可以是作为工业过程的副产物或从其它某个来源获得的废气,例如来自汽车废气、电解、热解、焙干或气化。例如,可以通过热解、焙干或气化来回收废料,以生成底物和/或c1碳源。在某些实施例中,工业过程选自由以下组成的组:含铁金属产品制造如钢厂制造、非含铁产品制造、石油精炼、煤炭气化、电力生产、碳黑生产、纸和纸浆制造、黑液气化、氨生产、甲醇生产以及焦炭制造。在这些实施例中,可以使用任何便利的方法从工业过程中捕获底物和/或c1碳源,然后将获底物和/或c1碳源排放到大气中。
[0114]
底物和/或c1碳源可以是合成气,如通过煤炭或精炼残余物的气化、生物质或木质纤维素材料的气化或天然气的重整而获得的合成气。在另一个实施例中,合成气可以从城市固体废物或工业固体废物的气化中获得。
[0115]
底物的组成可以对反应的效率和/或成本有显著影响。例如,氧气(o2)的存在可以降低厌氧发酵过程的效率。根据底物的组成,可能需要处理、擦洗或过滤底物以去除任何不期望的杂质如毒素、不期望的组分或灰尘颗粒和/或增加期望组分的浓度。
[0116]
在某些实施例中,发酵在不存在碳水化合物底物如糖、淀粉、木质素、纤维素或半纤维素的情况下进行。
[0117]
本公开的微生物可以用气态底物培养以产生一种或多种产物。例如,除了2
‑
苯乙醇之外,本公开的微生物还可以产生或还可以被工程化以产生乙醇(wo 2007/117157)、乙酸盐(wo 2007/117157)、1
‑
丁醇(wo 2008/115080、wo 2012/053905和wo 2017/066498)、丁酸盐(wo 2008/115080)、2,3
‑
丁二醇(wo 2009/151342和wo 2016/094334)、乳酸盐(wo 2011/112103)、丁烯(wo 2012/024522)、丁二烯(wo 2012/024522)、甲基乙基酮(2
‑
丁酮)(wo 2012/024522和wo 2013/185123)、乙烯(wo 2012/026833)、丙酮(wo 2012/115527)、异丙醇(wo 2012/115527)、脂质(wo 2013/036147)、3
‑
羟基丙酸盐(3
‑
hp)(wo 2013/180581)、包含异戊二烯在内的萜烯(wo 2013/180584)、脂肪酸(wo 2013/191567)、2
‑
丁醇(wo 2013/185123)、1,2
‑
丙二醇(wo 2014/036152)、1
‑
丙醇(wo 2017/066498)、1
‑
己醇(wo 2017/
066498)、1
‑
辛醇(wo 2017/066498)、分支酸盐衍生的产物(wo 2016/191625)、3
‑
羟基丁酸盐(wo 2017/066498)、1,3
‑
丁二醇(wo 2017/066498)、2
‑
羟基异丁酸盐或2
‑
羟基异丁酸(wo 2017/066498)、异丁烯(wo 2017/066498)、己二酸(wo 2017/066498)、1,3
‑
己二醇(wo 2017/066498)、3
‑
甲基
‑2‑
丁醇(wo 2017/066498)、2
‑
丁烯
‑1‑
醇(wo 2017/066498)、异戊酸盐(wo 2017/066498)、异戊醇(wo 2017/066498)和/或单乙二醇(wo 2019/126400)。在某些实施例中,微生物生物质本身可以被视为产物。这些产物可以被进一步转化,以产生柴油、喷气燃料和/或汽油中的至少一种组分。在某些实施例中,可以将2
‑
苯乙醇用作香精、精油、香料和肥皂中的成分。另外,可以进一步处理微生物生物质以产生单细胞蛋白(scp)。
[0118]“天然产物”是由未经基因修饰的微生物产生的产物。例如,乙醇、乙酸盐和2,3
‑
丁二醇是产乙醇梭菌、扬氏梭菌和拉氏梭菌的天然产物。“非天然产物”是由经基因修饰的微生物产生的产物,而不是由衍生出经基因修饰的微生物的未经基因修饰的微生物产生的产物。
[0119]“选择率”是指目标产物的产量与微生物所产生的全部发酵产物的产量的比率。本公开的微生物可以被工程化以便以特定选择率或最小选择率产生产物。在一个实施例中,目标产物占由本公开的微生物产生的全部发酵产物的至少约5%、10%、15%、20%、30%、50%或75%。在一个实施例中,目标产物占由本公开微的生物产生的全部发酵产物的至少10%,使得本公开的微生物针对目标产物的选择率为至少10%。在另一个实施例中,目标产物占由本公开的微生物产生的全部发酵产物的至少30%,使得本公开的微生物针对目标产物的选择率为至少30%。
[0120]“提高效率”、“提高的效率”等包含但不限于提高生长速率、产物产生速率或体积、消耗每体积底物的产物体积或产物选择率。可以相对于衍生出本公开的微生物的亲代微生物的性能来测量效率。
[0121]
培养通常在生物反应器中进行。术语“生物反应器”包含由一个或多个容器、塔或管路布置组成的培养/发酵装置,如连续搅拌槽反应器(cstr)、固定化细胞反应器(icr)、滴流床反应器(tbr)、气泡柱、气升式发酵器、静态混合器或适合气
‑
液接触的其它容器或其它装置。在一些实施例中,生物反应器可以包括第一生长反应器和第二培养/发酵反应器。可以向这些反应器中的一个或两个提供底物。如本文所使用的,术语“培养”和“发酵”可互换使用。这些术语涵盖培养/发酵过程的生长阶段和产物生物合成阶段两者。
[0122]
培养通常在含有足以允许微生物生长的营养素、维生素和/或矿物质的水性培养基中维持。优选地,水性培养基是厌氧微生物生长培养基,如基本厌氧微生物生长培养基。合适的培养基是本领域众所周知的。
[0123]
培养/发酵应当期望地在用于产生目标产物的适当条件下进行。培养/发酵通常在厌氧条件下进行。要考虑的反应条件包含压力(或分压)、温度、气体流速、液体流速、培养基ph、培养基氧化还原电势、搅拌速率(如果使用连续搅拌槽反应器的话)、接种物水平、用于确保处于液相的气体不会变成限制的最大气体底物浓度以及用于避免产物抑制的最大产物浓度。具体地,可以控制底物的引入速率来以确保处于液相的气体的浓度不会变成限制,因为在气体限制的条件下,产物可能会因培养而被消耗。
[0124]
在某些实施例中,发酵在没有光的情况下或在存在不足以满足光合微生物的能量需求的光量的情况下进行。在某些实施例中,本公开的微生物是非光合微生物。
[0125]
在一些实施例中,可以使用本领域中已知的任何方法或方法的组合从发酵液中分离或纯化2
‑
苯乙醇,所述方法包含例如分级蒸馏、蒸发、渗透蒸发、气体剥离、相分离和提取发酵(包含例如液体
‑
液体提取)。在某些实施例中,通过从生物反应器中连续去除发酵液的一部分、从发酵液中分离微生物细胞(便利地通过过滤)并从发酵液中回收2
‑
苯乙醇来从发酵液中回收2
‑
苯乙醇。醇和/或丙酮通常可以例如通过蒸馏回收。分离的微生物细胞优选地回收到生物反应器中。去除2
‑
苯乙醇之后残留的无细胞渗透物也优选地返回到生物反应器中。可以向无细胞渗透物添加另外的营养素来补给培养基,随后使其返回到生物反应器中。
[0126]
实施例
[0127]
在一个实施例中,本公开提供了一种能够产生2
‑
苯乙醇的微生物,其中所述微生物包括将苯丙酮酸盐转化为苯乙醛的异源酶和将苯乙醛转化为2
‑
苯乙醇的异源酶。
[0128]
在一些实施例中,所述将苯丙酮酸盐转化为苯乙醛的异源酶为脱羧酶,并且所述将苯乙醛转化为2
‑
苯乙醇的异源酶为苯乙醛还原酶。具体地,所述脱羧酶可以为苯丙酮酸盐特异性脱羧酶。在一些实施例中,编码异源脱羧酶的核酸包括aro10(来自酿酒酵母(saccharomyces cerevisiae))或abppdc(来自巴西固氮螺菌(azospirillum brasilense))。在一些实施例中,编码异源脱羧酶的核酸包括:(i)seq id no:1、2或4;(ii)与编码脱羧酶的seq id no:1、2或4具有至少90%同一性的核酸,所述核酸在功能上与由seq id no:1、2或4编码的所述脱羧酶同源;或(iii)编码与由seq id no:1、2或4编码的所述脱羧酶具有至少90%同一性的功能同源多肽的核酸。
[0129]
在一些实施例中,编码异源脱羧酶的核酸包括aro10、abppdc或者seq id no:1、2或4或者与其共享例如至少70%或至少75%或至少80%或至少85%或至少86%或至少87%或至少88%或至少89%或至少90%或至少91%或至少92%或至少93%或至少94%或至少95%或至少96%或至少97%或至少98%或至少99%或100%核酸序列同一性的功能同系物。在一些实施例中,编码异源脱羧酶的核酸编码与由aro10、abppdc或者seq id no:1、2或4编码的脱羧酶有例如至少70%或至少75%或至少80%或至少85%或至少86%或至少87%或至少88%或至少89%或至少90%或至少91%或至少92%或至少93%或至少94%或至少95%或至少96%或至少97%或至少98%或至少99%或100%多肽序列同一性的功能同源多肽。
[0130]
在一些实施例中,编码异源par酶的核酸包括blpar、ecpar、lepar、rrpar、rspar或adh6(来源列于图4a中)。在一些实施例中,编码异源par酶的核酸包括:(i)seq id no:3、5、6、7、8或9;(ii)与编码par酶的seq id no:3、5、6、7、8或9具有至少90%同一性的核酸,所述核酸在功能上与由seq id no:3、5、6、7、8或9编码的所述par酶同源;或(iii)编码与由seq id no:3、5、6、7、8或9编码的所述par酶具有至少90%同一性的功能同源多肽的核酸。
[0131]
在一些实施例中,编码异源par酶的核酸包括blpar、ecpar、lepar、rrpar、rspar、adh6或者seq id no:3、5、6、7、8或9或者与其共享例如至少70%或至少75%或至少80%或至少85%或至少86%或至少87%或至少88%或至少89%或至少90%或至少91%或至少92%或至少93%或至少94%或至少95%或至少96%或至少97%或至少98%或至少99%或100%核酸序列同一性的功能同系物。在一些实施例中,编码异源par酶的核酸编码与由blpar、ecpar、lepar、rrpar、rspar、adh6或者seq id no:3、5、6、7、8或9编码的par酶有例如至少70%或至少75%或至少80%或至少85%或至少86%或至少87%或至少88%或至少
89%或至少90%或至少91%或至少92%或至少93%或至少94%或至少95%或至少96%或至少97%或至少98%或至少99%或100%多肽序列同一性的功能同源多肽。
[0132]
在一些实施例中,根据本公开所述的微生物进一步包括以下中的一种或多种:将磷酸烯醇丙酮酸盐和赤藓糖
‑4‑
磷酸盐转化为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐的异源酶;将3
‑
脱氢莽草酸盐转化为莽草酸盐的异源酶;或将分支酸盐转化为苯丙酮酸盐的异源酶。
[0133]
在一些实施例中,所述将磷酸烯醇丙酮酸盐和赤藓糖
‑4‑
磷酸盐转化为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐的异源酶为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸合酶;所述将3
‑
脱氢莽草酸盐转化为莽草酸盐的异源酶为莽草酸脱氢酶;或所述将分支酸盐转化为苯丙酮酸盐的异源酶为双官能分支酸变位酶/预苯酸脱水酶。
[0134]
在一些实施例中,编码异源莽草酸脱氢酶的核酸包括cgaroe(来自谷氨酸棒状杆菌(corynebacterium glutamicum))或ecaroe(来自大肠杆菌(escherichia coli))。在一些实施例中,编码异源莽草酸脱氢酶的核酸包括:(i)seq id no:11或12;(ii)与编码莽草酸脱氢酶的seq id no:11或12具有至少90%同一性的核酸,所述核酸在功能上与由seq id no:11或12编码的所述莽草酸脱氢酶同源;或(iii)编码与由seq id no:11或12编码的所述莽草酸脱氢酶具有至少90%同一性的功能同源多肽的核酸。
[0135]
在一些实施例中,编码异源莽草酸脱氢酶的核酸包括cgaroe、ecaroe或者seq id no:11或12或者与其共享例如至少70%或至少75%或至少80%或至少85%或至少86%或至少87%或至少88%或至少89%或至少90%或至少91%或至少92%或至少93%或至少94%或至少95%或至少96%或至少97%或至少98%或至少99%或100%核酸序列同一性的功能同系物。在一些实施例中,编码异源莽草酸脱氢酶的核酸编码与由cgaroe、ecaroe或者seq id no:11或12编码的莽草酸脱氢酶有例如至少70%或至少75%或至少80%或至少85%或至少86%或至少87%或至少88%或至少89%或至少90%或至少91%或至少92%或至少93%或至少94%或至少95%或至少96%或至少97%或至少98%或至少99%或100%多肽序列同一性的功能同源多肽。
[0136]
在一些实施例中,编码异源dahp合酶的核酸包括arog。在一些实施例中,编码异源dahp合酶的核酸包括:(i)seq id no:10;(ii)与编码dahp合酶的seq id no:10具有至少90%同一性的核酸,所述核酸在功能上与由seq id no:10编码的所述dahp合酶同源;或(iii)编码与由seq id no:10编码的所述dahp合酶具有至少90%同一性的功能同源多肽的核酸。
[0137]
在一些实施例中,编码异源dahp合酶的核酸包括arog或seq id no:10或与其共享例如至少70%或至少75%或至少80%或至少85%或至少86%或至少87%或至少88%或至少89%或至少90%或至少91%或至少92%或至少93%或至少94%或至少95%或至少96%或至少97%或至少98%或至少99%或100%核酸序列同一性的功能同系物。在一些实施例中,编码异源dahp合酶的核酸编码与由arog或seq id no:10编码的dahp合酶共享例如至少70%或至少75%或至少80%或至少85%或至少86%或至少87%或至少88%或至少89%或至少90%或至少91%或至少92%或至少93%或至少94%或至少95%或至少96%或至少97%或至少98%或至少99%或100%多肽序列同一性的功能同源多肽。
[0138]
在一些实施例中,编码异源双官能分支酸变位酶/预苯酸脱水酶的核酸包括phea1
或phea2。在一些实施例中,编码异源双官能分支酸变位酶/预苯酸脱水酶的核酸包括:(i)seq id no:13或14;(ii)与编码双官能分支酸变位酶/预苯酸脱水酶的seq id no:13或14具有至少90%同一性的核酸,所述核酸在功能上与由seq id no:13或14编码的所述双官能分支酸变位酶/预苯酸脱水酶同源;或(iii)编码与由seq id no:13或14编码的所述双官能分支酸变位酶/预苯酸脱水酶具有至少90%同一性的功能同源多肽的核酸。
[0139]
在一些实施例中,编码异源双官能分支酸变位酶/预苯酸脱水酶的核酸包括phea1、phea2或者seq id no:13或14或者与其共享例如至少70%或至少75%或至少80%或至少85%或至少86%或至少87%或至少88%或至少89%或至少90%或至少91%或至少92%或至少93%或至少94%或至少95%或至少96%或至少97%或至少98%或至少99%或100%核酸序列同一性的功能同系物。在一些实施例中,编码异源双官能分支酸变位酶/预苯酸脱水酶的核酸编码与由phea1、phea2或者seq id no:13或14编码的双官能分支酸变位酶/预苯酸脱水酶共享例如至少70%或至少75%或至少80%或至少85%或至少86%或至少87%或至少88%或至少89%或至少90%或至少91%或至少92%或至少93%或至少94%或至少95%或至少96%或至少97%或至少98%或至少99%或100%多肽序列同一性的功能同源多肽。
[0140]
在一些实施例中,所述微生物包括选自以下选项中的一组异源酶:
[0141]
(1)ecaroe、kivd、lepar、phea2和arog;
[0142]
(2)ecaroe、abppdc、ecpar、phea1和arog;
[0143]
(3)cgaroe、abppdc、lepar、phea2和arog;
[0144]
(4)ecaroe、abppdc、lepar、phea2和arog;
[0145]
(5)cgaroe、kivd、rrpar、phea2和arog;
[0146]
(6)ecaroe、abppdc、lepar、phea2和arog;
[0147]
(7)phea1、arog、abppdc和ecpar;
[0148]
(8)egaroe、abppdc、lepar、phea1和arog;
[0149]
(9)ecaroe、abppdc、ecpar、phea2和arog;
[0150]
(10)cgaroe、aro10、ecpar、phea2和arog;
[0151]
(11)cgaroe、abppdc、lepar、phea1和arog;
[0152]
(12)cgaroe、aro10、lepar、phea2和arog;
[0153]
(13)ecaroe、abppdc、lepar、phea1和arog;
[0154]
(14)ecaroe、aro10、ecpar、phea2和arog;
[0155]
(15)ecaroe、aro10、lepar、phea2和arog;
[0156]
(16)cgaroe、abppdc、lepar、phea1和arog;
[0157]
(17)cgaroe、aro10、rspar、phea2和arog;
[0158]
(18)cgaroe、abppdc、blpar、phea2和arog;
[0159]
(19)ecaroe、kivd、blpar、phea2和arog;
[0160]
(20)cgaroe、aro10、blpar、phea2和arog;
[0161]
(21)ecaroe、abppdc、blpar、phea2和arog;
[0162]
(22)ecaroe、abppdc、adh6、phea1和arog;
[0163]
(23)cgaroe、abppdc、adh6、phea2和arog;
[0164]
(24)cgaroe、abppdc、ecpar、phea2和arog;
[0165]
(25)ecaroe、aro10、rspar、phea2和arog;
[0166]
(26)ecaroe、aro10、adh6、phea2和arog;或
[0167]
(27)ecaroe、abppdc、ecpar、phea2和arog。
[0168]
在一些实施例中,所述微生物包括选自以下选项中的一组异源核酸:(1)ecaroe、kivd、lepar、phea2和arog;
[0169]
(2)ecaroe、abppdc、ecpar、phea1和arog;
[0170]
(3)cgaroe、abppdc、lepar、phea2和arog;
[0171]
(4)ecaroe、abppdc、lepar、phea2和arog;
[0172]
(5)cgaroe、kivd、rrpar、phea2和arog;
[0173]
(6)ecaroe、abppdc、lepar、phea2和arog;
[0174]
(7)phea1、arog、abppdc和ecpar;
[0175]
(8)egaroe、abppdc、lepar、phea1和arog;
[0176]
(9)ecaroe、abppdc、ecpar、phea2和arog;
[0177]
(10)cgaroe、aro10、ecpar、phea2和arog;
[0178]
(11)cgaroe、abppdc、lepar、phea1和arog;
[0179]
(12)cgaroe、aro10、lepar、phea2和arog;
[0180]
(13)ecaroe、abppdc、lepar、phea1和arog;
[0181]
(14)ecaroe、aro10、ecpar、phea2和arog;
[0182]
(15)ecaroe、aro10、lepar、phea2和arog;
[0183]
(16)cgaroe、abppdc、lepar、phea1和arog;
[0184]
(17)cgaroe、aro10、rspar、phea2和arog;
[0185]
(18)cgaroe、abppdc、blpar、phea2和arog;
[0186]
(19)ecaroe、kivd、blpar、phea2和arog;
[0187]
(20)cgaroe、aro10、blpar、phea2和arog;
[0188]
(21)ecaroe、abppdc、blpar、phea2和arog;
[0189]
(22)ecaroe、abppdc、adh6、phea1和arog;
[0190]
(23)cgaroe、abppdc、adh6、phea2和arog;
[0191]
(24)cgaroe、abppdc、ecpar、phea2和arog;
[0192]
(25)ecaroe、aro10、rspar、phea2和arog;
[0193]
(26)ecaroe、aro10、adh6、phea2和arog;或
[0194]
(27)ecaroe、abppdc、ecpar、phea2和arog。
[0195]
在一些实施例中,所述微生物包括组合菌株2c03、1c10、2c54、2c09、2c47、2c55、2c53、h57、1c47、2c10、2a18、1c54、2c01、1c31、2b17、2c52、1c09、h18、h58、2c27、2d22、2d07、2d24、2d03、1c29、2b26、2c38、2a27、2c12或2b27的异源基因和启动子,如表3中所描述的。
[0196]
在另一个实施例中,本公开提供了质粒,所述质粒可以用于转化细菌以产生本公开的2
‑
苯乙醇性细菌。在一些实施例中,所述质粒包括(a)编码异源脱羧酶的核酸;以及(b)编码异源苯乙醛还原酶(par)的核酸。在一些实施例中,所述质粒包括(a)编码异源脱羧酶的核酸;(b)编码异源苯乙醛还原酶(par)的核酸;(c)编码异源莽草酸脱氢酶的核酸;(d)编
码异源dahp合酶的核酸;以及(e)编码异源双官能分支酸变位酶/预苯酸脱水酶的核酸。在一些实施例中,本公开提供了一种双质粒系统,第一质粒包括(a)编码异源脱羧酶的核酸;(b)编码异源苯乙醛还原酶(par)的核酸;和(c)编码异源莽草酸脱氢酶的核酸;并且第二质粒包括(d)编码异源dahp合酶的核酸;以及(e)编码异源双官能分支酸变位酶/预苯酸脱水酶的核酸。当一个或多个质粒转化到细菌中时,期望的是,细菌表达所述酶。替代性地,如果将诱导型启动子用于表达,则所述酶仅当被诱导时才可以被表达。
[0197]
一个实施例是一种能够产生2
‑
苯乙醇的微生物,其中所述微生物包括:(a)将苯丙酮酸盐转化为苯乙醛的异源酶;以及(b)将苯乙醛转化为2
‑
苯乙醇的异源酶。
[0198]
根据实施例所述的微生物,其中:(a)所述将苯丙酮酸盐转化为苯乙醛的异源酶为脱羧酶;并且(b)所述将苯乙醛转化为2
‑
苯乙醇的异源酶为苯乙醛还原酶。
[0199]
根据实施例所述的微生物,其中所述脱羧酶为苯丙酮酸盐特异性脱羧酶。
[0200]
根据实施例所述的微生物,其中所述微生物为c1固定型微生物。
[0201]
根据实施例所述的微生物,其中所述微生物为伍德
‑
隆达尔微生物。
[0202]
根据实施例所述的微生物,其中所述微生物为细菌。
[0203]
根据实施例所述的微生物,其中所述微生物是选自由醋酸杆菌属、碱杆菌属、布劳特氏菌属、丁酸杆菌属、梭菌属、真杆菌属、穆尔氏菌属、产醋杆菌属、鼠孢菌属和嗜热厌氧杆菌属组成的组的属的成员。
[0204]
根据实施例所述的微生物,其中所述微生物天然地能够产生苯丙酮酸盐。
[0205]
根据实施例所述的微生物,其中所述微生物包括将co、co2和/或h2转化为乙酰辅酶a的伍德
‑
隆达尔途径。
[0206]
根据实施例所述的微生物,其中所述微生物包括以下中的一种或多种:(c)将乙酰辅酶a转化为丙酮酸盐的天然酶;(d)将丙酮酸盐转化为磷酸烯醇丙酮酸盐的天然酶;(e)将磷酸烯醇丙酮酸盐和赤藓糖
‑4‑
磷酸盐转化为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐的天然酶;(f)将2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐转化为3
‑
脱氢奎尼酸盐的天然酶;(g)将3
‑
脱氢奎尼酸盐转化为3
‑
脱氢莽草酸盐的天然酶;(h)将3
‑
脱氢莽草酸盐转化为莽草酸盐的天然酶;(i)将莽草酸盐转化为莽草酸3
‑
磷酸盐的天然酶;(j)将莽草酸3
‑
磷酸盐转化为5
‑
o
‑
(1
‑
羧基乙烯基)
‑
莽草酸3
‑
磷酸盐的天然酶;(k)将5
‑
o
‑
(1
‑
羧基乙烯基)
‑
莽草酸3
‑
磷酸盐转化为分支酸盐的天然酶;或(l)将分支酸盐转化为苯丙酮酸盐的天然酶。
[0207]
根据实施例所述的微生物,其中:(c)所述将乙酰辅酶a转化为丙酮酸盐的天然酶为丙酮酸盐:铁氧还蛋白氧化还原酶;(d)所述将丙酮酸盐转化为磷酸烯醇丙酮酸盐的天然酶为丙酮酸磷酸双激酶;(e)所述将磷酸烯醇丙酮酸盐和赤藓糖
‑4‑
磷酸盐转化为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐的天然酶为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸合酶;(f)所述将2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐转化为3
‑
脱氢奎尼酸盐的天然酶为3
‑
脱氢奎尼酸合酶;(g)所述将3
‑
脱氢奎尼酸盐转化为3
‑
脱氢莽草酸盐的天然酶为3
‑
脱氢奎尼酸脱水酶;(h)所述将3
‑
脱氢莽草酸盐转化为莽草酸盐的天然酶为莽草酸脱氢酶;(i)所述将莽草酸盐转化为莽草酸3
‑
磷酸盐的天然酶为莽草酸激酶;(j)所述将莽草酸3
‑
磷酸盐转化为5
‑
o
‑
(1
‑
羧基乙烯基)
‑
莽草酸3
‑
磷酸盐的天然酶为5
‑
烯醇丙酮酸莽草酸3
‑
磷酸合酶;(k)所述将5
‑
o
‑
(1
‑
羧基乙烯基)
‑
莽草酸3
‑
磷酸盐转化为分支酸盐的天然酶为
分支酸合酶;或(l)所述将分支酸盐转化为苯丙酮酸盐的天然酶为双官能分支酸变位酶/预苯酸脱水酶。
[0208]
根据实施例所述的微生物,其中所述微生物进一步包括以下中的一种或多种:(e)将磷酸烯醇丙酮酸盐和赤藓糖
‑4‑
磷酸盐转化为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐的异源酶;(h)将3
‑
脱氢莽草酸盐转化为莽草酸盐的异源酶;或(l)将分支酸盐转化为苯丙酮酸盐的异源酶。
[0209]
根据实施例所述的微生物,其中:(e)所述将磷酸烯醇丙酮酸盐和赤藓糖
‑4‑
磷酸盐转化为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸盐的异源酶为2
‑
脱氢
‑3‑
脱氧
‑
d
‑
阿拉伯糖
‑
庚糖酸7
‑
磷酸合酶;(h)所述将3
‑
脱氢莽草酸盐转化为莽草酸盐的异源酶为莽草酸脱氢酶;或(l)所述将分支酸盐转化为苯丙酮酸盐的异源酶为双官能分支酸变位酶/预苯酸脱水酶。
[0210]
根据所述实施例中任一项所述的微生物,其中所述微生物包括:
[0211]
(1)ecaroe、kivd、lepar、phea2和arog;
[0212]
(2)ecaroe、abppdc、ecpar、phea1和arog;
[0213]
(3)cgaroe、abppdc、lepar、phea2和arog;
[0214]
(4)ecaroe、abppdc、lepar、phea2和arog;
[0215]
(5)cgaroe、kivd、rrpar、phea2和arog;
[0216]
(6)ecaroe、abppdc、lepar、phea2和arog;
[0217]
(7)phea1、arog、abppdc和ecpar;
[0218]
(8)egaroe、abppdc、lepar、phea1和arog;
[0219]
(9)ecaroe、abppdc、ecpar、phea2和arog;
[0220]
(10)cgaroe、aro10、ecpar、phea2和arog;
[0221]
(11)cgaroe、abppdc、lepar、phea1和arog;
[0222]
(12)cgaroe、aro10、lepar、phea2和arog;
[0223]
(13)ecaroe、abppdc、lepar、phea1和arog;
[0224]
(14)ecaroe、aro10、ecpar、phea2和arog;
[0225]
(15)ecaroe、aro10、lepar、phea2和arog;
[0226]
(16)cgaroe、abppdc、lepar、phea1和arog;
[0227]
(17)cgaroe、aro10、rspar、phea2和arog;
[0228]
(18)cgaroe、abppdc、blpar、phea2和arog;
[0229]
(19)ecaroe、kivd、blpar、phea2和arog;
[0230]
(20)cgaroe、aro10、blpar、phea2和arog;
[0231]
(21)ecaroe、abppdc、blpar、phea2和arog;
[0232]
(22)ecaroe、abppdc、adh6、phea1和arog;
[0233]
(23)cgaroe、abppdc、adh6、phea2和arog;
[0234]
(24)cgaroe、abppdc、ecpar、phea2和arog;
[0235]
(25)ecaroe、aro10、rspar、phea2和arog;
[0236]
(26)ecaroe、aro10、adh6、phea2和arog;或
[0237]
(27)ecaroe、abppdc、ecpar、phea2和arog。
[0238]
根据所述实施例中任一项所述的微生物,其中所述微生物包括组合菌株2c03、1c10、2c54、2c09、2c47、2c55、2c53、h57、1c47、2c10、2a18、1c54、2c01、1c31、2b17、2c52、1c09、h18、h58、2c27、2d22、2d07、2d24、2d03、1c29、2b26、2c38、2a27、2c12或2b27的异源基因和启动子。
[0239]
根据所述实施例中任一项所述的微生物,其中所述微生物发酵包括co、co2和/或h2的气态底物以产生2
‑
苯乙醇。
[0240]
根据实施例所述的微生物,其中所述气态底物包括合成气或工业废气。
[0241]
根据实施例所述的微生物,其中所述微生物不产生任何其它c3+醇。
[0242]
根据实施例所述的微生物,其中所述微生物不产生任何其它c3、c4、c5、c6、c7、c8、c9或c10醇。
[0243]
一个实施例是一种产生2
‑
苯乙醇的方法,所述方法包括在气态底物存在的情况下培养根据所述实施例中任一项所述的微生物。
[0244]
根据实施例所述的方法,其中所述气态底物包括c1碳源,所述c1碳源包括co、co2和/或h2。
[0245]
根据所述实施例中任一项所述的方法,其中所述气态底物包括合成气或工业废气。
[0246]
实例
[0247]
以下实例进一步说明了本公开的方法和组合物,但不应被解释为以任何方式限制本公开的范围。
[0248]
在此工作中,在c1固定型细菌中进行了脱羧酶和苯乙醛还原酶(par)的靶向代谢工程化,以提高2
‑
pe的选择率和产量。为了进一步提高通向2
‑
pe生物合成的通量,以组合的方式另外包含了来自莽草酸盐途径的三个异源基因。图1中突出显示了2
‑
pe生物合成途径和在以下实例中异源性表达的基因。
[0249]
实例1.肖特瓶中的产乙醇梭菌的2
‑
pe耐受性
[0250]
已经报告2
‑
pe在26
‑
43mm(3.2
‑
5.3g/l)水平下对革兰氏阳性微生物如金黄色葡萄球菌(staphylococcus aureus)和粪肠球菌(enterococcus faecium)具有生长抑制作用(corre等人,《微生物学研究(res microbiol.)》。1990;141(4):483
‑
97)。为了研究2
‑
pe对产乙醇梭菌的自养生长的毒性,在肖特瓶中使用合成气共混物(50%co、10%h2、30%co2和10%n2)进行2
‑
pe攻击实验(在0g/l、0.2g/l、0.6g/l、2g/l和5g/l下)。与未经攻击的培养相比,2g/l的2
‑
pe导致生长滞后期增加并且生物质浓度较低(图2a
‑
b)。在5g/l攻击水平下,未检测到生长(图2a
‑
b)。0.2g/l和0.6g/l的2
‑
pe攻击对产乙醇梭菌的自养生长几乎没有影响。
[0251]
实例2.针对提高的2
‑
pe选择率和产量对脱羧酶变体的测试
[0252]
为了进行2
‑
pe菌株优化,将具有苯丙酮酸盐特异性脱羧酶(来自酿酒酵母的aro10(kneen等人,《欧洲生物化学联合会杂志(febs j.)》2011;278:1842
‑
53)(seq id no:4)和来自巴西固氮螺菌的abppdc(spaepen等人,《细菌学杂志(j bacteriol.)》2007;189:7626lp
‑
7633)(seq id no:2))的两个质粒转化到产乙醇梭菌中,从而分别得到菌株sfa212和sfa213。这两个脱羧酶菌株与表达脱羧酶kivd_la(来自乳酸乳球菌(lactococcus lactis))(seq id no:1)的对照菌株(sfa200)一起在肖特瓶中的合成气混合物(50%co、
10%h2、30%co2和10%n2)下进行自养生长(图3a
‑
b)。这三个脱羧酶菌株显示出类似的生长曲线,但醇曲线不同(图3a
‑
b)。带有火焰电离检测的终点气相色谱法(gc
‑
fid)结果表明,aro10菌株产生的2
‑
pe比对照kivd_la菌株少57%,同时产生了5.6mg/l的2
‑
甲基
‑1‑
丙醇和4.0mg/l的3
‑
甲基
‑1‑
丁醇。相比之下,abppdc菌株产生的2
‑
pe量与对照菌株类似,但均未产生支链醇。在一些实施例中,通过abppdc菌株对2
‑
pe的选择率的增加可以简化下游产物分离并改善菌株稳定性,因为支链醇可以对细胞毒性具有累加效应。
[0253]
对于此实例和本文所公开的其它实例,如下针对直链醇和支链醇进行gc
‑
fid。使用配备有自动进样器、火焰电离检测(fid)和与zb
‑
1柱(30m
×
0.32mm
×
0.3μm)串联连接的phenomenex zb
‑
waxplus柱(30m
×
0.32mm
×
1μm)的agilent 7890b gc通过气相色谱法分析来测量醇浓度。通过将1.400ml样品转移到干净的2
‑
ml微量离心管中、然后加入20μl内标溶液(含乙酸苯酯的乙醇)来制备样品。向此样品中加入400μl氯仿并记录质量。将样品水平地震荡60秒、以14,000x g离心5分钟,然后将200μl底层转移到含有小容量插入物的玻璃小瓶中。使用10:1的分流比并且在250℃的入口温度下进行1
‑
ul进样分析。在起始柱箱温度为70℃(不保持)、初始以3℃/分钟升温到120℃(不保持)并且最终以7℃/分钟升温到230℃(保持14分钟)的情况下实现分离。以氦气为载气,将柱流速设置为35厘米/秒。将fid设置为280℃,其中空气为400毫升/分钟、氢气为40毫升/分钟并且氦气(补充气体)为15毫升/分钟。使用线性拟合利用内标校准来计算结果。
[0254]
实例3.针对提高的2
‑
pe产量对苯乙醛还原酶(par)变体的测试
[0255]
将催化2
‑
pe途径(图1)的最后步骤的苯乙醛还原酶(par)的五种基因变体(lepar(seq id no:9)、ecpar(seq id no:6)、blpar(seq id no:5)、rrpar(seq id no:7)和rspar(seq id no:8);图4a
‑
b)用脱羧酶abppdc(seq id no:2)和莽草酸脱氢酶ecaroe(seq id no:11)单独克隆到表达载体中。在转化到产乙醇梭菌中后,在具有200kpa合成气混合物(50%co、10%h2、30%co2和10%n2)的12孔板中对所得重组菌株的2
‑
pe产量进行测试。在第1天加入诱导剂,并在第7天收集样品以进行gc
‑
fid分析。新par菌株中的三个菌株(blpar、ecpar和rspar)产生了比adh6(seq id no:3)对照菌株更多的2
‑
pe(通过生物质归一化)(图4c)。
[0256]
实例4.对2
‑
pe途径的组合分析
[0257]
为了确定2
‑
pe的生物合成的最佳通量和基因变体,首先在大肠杆菌中使用金门(gg)方法对2
‑
pe途径基因和启动子进行了组合组装(图5),然后转化到利用c1的微生物中。为了便于组装的筛选,首先将侧接金门位点gg1和gg6的ccdb毒素
‑
抗毒素反向选择性标记物克隆到梭菌
‑
大肠杆菌穿梭载体pmtl8225(heap,《微生物学方法杂志(j microbiol methods)》,78:79
‑
85,2009)中。这些穿梭载体具有预先克隆的梭菌启动子(在图5中示出为p1)和终止子(在图5中示出为t3)。将编码莽草酸脱氢酶(由aroe编码)、dahp合酶(由arog编码)、双官能分支酸变位酶/预苯酸脱水酶(phea)、脱羧酶和苯乙醛(par)的基因与两个供体载体(pdonor 2和pdonor 4,提供了终止子和启动子)一起加入到穿梭载体中以用于gg组装。启动子序列、金门位点和组装工作流程在《合成生物学(synthetic biology)》(2020)第5(1)卷:ysaa019中进行了描述。具有ermb抗生素选择性标记物的所得组合质粒(表3中统称为质粒1)具有用于以绝缘方式表达aroe(或phea+arog)、脱羧酶和par中的每一个的启动子和终止子。
[0258]
针对此工作,构建并测试了两个2
‑
pe组合文库:单质粒文库和双质粒文库(表3)。表2中示出了组合分析中所包含的基因变体(2x aroe(seq id no:11、12)、3x脱羧酶(seq id no:1、2、4)、6x par(seq id no:3、5、6、7、8、9)、2x双官能分支酸变位酶/预苯酸脱水酶(phea)(seq id no:13、14)和1x dahp合酶(arog)(seq id no:10))。图12示出了用于实例4中所描述的两个2
‑
pe组合文库的基因和启动子变体。
[0259]
表2.用于组合分析的2
‑
pe途径基因变体。
[0260][0261][0262]
在双质粒文库的情况下,使用强度不同的三个启动子(p
fer
‑
laco
‑
us
、p
wl
和p
pfor
)来表
达质粒1中的aroe、脱羧酶和par中的每一个。具有驱动phea1+arog或phea2+arog的表达的仅一个启动子(p
wl
)的质粒2(catp抗生素选择性标记物)被单独转化到产乙醇梭菌中。所得的转化体随后成为转移组合质粒1的宿主,从而产生了携带两个表达载体的重组菌株,所述两个表达载体赋予了相容的革兰氏阳性复制子不同的抗生素抗性。双质粒组合文库的总排列为1944(3
×2×3×3×3×6×1×2×
1)。
[0263]
在单质粒文库的情况下,质粒1中的aroe被(phea1/phea2)+arog替换。这产生了仅1个质粒(省略了aroe)和排列972(3
×2×3×3×3×
6)。在此过程期间,核糖体结合位点与arog的起始密码子之间的间隔子距离从5个核苷酸扩展到8个核苷酸,以增强arog的翻译。
[0264]
为了确定组合质粒在大肠杆菌中的组装效率并且为了研究启动子和基因变体组合,从转化的大肠杆菌菌株neb10
‑
β中提取400个质粒并对其进行测序。对测序结果的分析表明组装率为51.3%,其中每个启动子和基因变体具有良好的多样性。将这些经过序列验证的组合质粒转化到产乙醇梭菌中之后,使总共162个组合菌株(42个单质粒菌株和120个双质粒菌株)在12孔板中进行自养生长。
[0265]
在37℃下在具有2ml基本培养基和200kpa合成气混合物(50%co、10%h2、30%co2和10%n2)的12孔板中利用技术复制品进行8
‑
10天的生长实验(图7a
‑
c)。然后取得肉汤样品以用于生物质测量和gc
‑
fid分析以确定2
‑
pe滴度。选择载有在诱导型启动子p
ipl12
下表达adh6与abppdc的质粒(与质粒1具有相同的主链)的菌株sfa212的三个生物学克隆作为对照菌株,因为此菌株在12孔板、肖特瓶和连续搅拌槽反应器(cstr)中一致地产生较高的2
‑
pe量。将每12孔板一个培养基空白对照孔作为阴性对照纳入,并且gc
‑
fid分析示出<1.7mg/l 2
‑
pe,这表明有很少到没有交叉孔污染。
[0266]
温育8
‑
10天后,2
‑
pe组合菌株的生物质浓度达到0.6
‑
1.0gdcw/l(图7c)。在这162个组合菌株中,有80个菌株(占所筛选文库的49%)产生的2
‑
pe比对照菌株多>1.5倍(图7a)。48个菌株(所筛选文库的30%)产生的2
‑
pe不到对照菌株所产生的2
‑
pe的一半(图7a)。
[0267]
对2
‑
pe组合文库进行详细分析,以识别基因变体和质粒数对2
‑
pe产量的影响(图8a
‑
e)。在单质粒文库与双质粒文库、脱羧酶与莽草酸脱氢酶之间未观察到显著的2
‑
pe滴度差异(图8a、8c和8d)。具有phea2的菌株示出比具有phea1菌株略高的2
‑
pe滴度(图8b)。par变体(例如,blpar)中的一些产生的2
‑
pe比adh6变体产生更多(图8e)。在每个基因变体或质粒数中观察到的较大的2
‑
pe滴度变化可以归因于其它途径基因变体和启动子组合的集体效应,所述集体效应稀释了组合设计中的单一参数的影响。
[0268]
通过相对于对照菌株sfa212对2
‑
pe滴度进行排名,得出了前30种产2
‑
pe组合菌株与其基因型的列表(表3)。尽管所筛选组合文库的74%是具有任一aroe的双质粒菌株,但前30个产2
‑
pe菌株中的90%携带了两个质粒。所有前10个产2
‑
pe菌株均由脱羧酶变体kivd_la或abppdc组成,而具有脱羧酶aro10的菌株在前11个与前30个2
‑
pe生产者之间出现8次。具有par变体lepar和ecpar的菌株占前30个2
‑
pe生产者的70%,这相对于其在所筛选文库中的总体呈现度57%而言不成比例地更高。具有par变体adh6的菌株在前30个2
‑
pe生产者中显著呈现过低:在前30个中为仅6.7%相较于在总体所筛选文库中为13.6%。
[0269]
表3.12孔板生长测定中的前30个产2
‑
pe组合菌株的基因型。
[0270][0271]
实例5.来自cstr中的合成气发酵的2
‑
pe产量
[0272]
在连续发酵模式下,在cstr中表征总共12个组合菌株,以比较其相较于参考菌株sfa212的2
‑
pe产量。使用来自肖特瓶的活跃生长的(早期指数的)培养物作为用于在大气压下使用合成气共混物(40%co、20%h2、20%co2和20%n2)的2
‑
l cstr的接种物。一旦生物质浓度达到约0.5gdcw/l,就维持1/天的培养基稀释速率和0.5/天的细菌稀释速率。
[0273]
在这些连续cstr条件下,参考菌株sfa212的2
‑
pe滴度达到70mg/l。五个组合菌株(1c29、2c54、2c03、2c53和2c31)产生的2
‑
pe比参考菌株多(图9a)。具体地,菌株1c29产生200mg/l的2
‑
pe,比参考菌株高2.9倍。基于其2
‑
pe产率,12个组合菌株中有10个相对于参考菌株显示出优越的2
‑
pe性能(图9b)。
[0274]
组合菌株1c29在两次重复的cstr试验中的性能在图10a
‑
b中示出。为了提高2
‑
pe产量,对于重复1(图10a)在第7天并且对于重复2(图10b)在第9天将nh4oh形式的氮供应物限制在15mm。结果,2
‑
pe产量在两个试验中均增加并且分别在350mg/l和300mg/l处达到峰值。试验1的2
‑
pe生产率在16毫克/升/小时时瞬间达到峰值。
[0275]
衍生自废料如生物质、林业残留物和市政固体废物的气化的合成气通常含有毒性污染物。例如,甲烷、乙烷、乙烯和乙炔都在检测到的污染物之列,并且已知所述甲烷、乙烷、乙烯和乙炔在ppm水平上对微生物有毒性。为了评估工程化的产乙醇梭菌中的从头2
‑
pe生物合成的稳健性,使用衍生自玉米秸秆的合成气对菌株sfa212进行连续cstr试验。这种“实际”合成气使用气体处理系统进行预先处理,从而得到23.3%h2、38.7%co、19.6%co2和18.4%n2的处理后气体组合物。由于实际合成气的可用性有限,因此最初使用合成共混气
(具有类似的气体组成)来建立稳态发酵培养,之后在第6.9天将气体供应物转变为实际合成气。
[0276]
实际合成气发酵的结果在图11a
‑
c中示出,其中在第6.9天与第10.9天之间(总共4天)使用了实际合成气。为了实现更长的连续cstr试验,使用气体组成略有不同的若干个实际合成气瓶,这导致了观察到的气体摄取波动(图11b)。转变为实际合成气后,2
‑
pe产量继续增加并在第10天达到稳定状态。第8.0天与第13.7天之间的平均2
‑
pe滴度和生产率分别为208毫克/升/小时和11.95毫克/升/小时。
[0277]
序列
[0278]
表4.实例中描述并且表2中汇总的核酸的序列。
[0279]
[0280]
[0281]
[0282]
[0283]
[0284]
[0285]
[0286]
[0287][0288][0289]
本文引用的所有参考文献(包含出版物、专利申请和专利)均通过引用特此并入,
其程度如同每篇参考文献被单独并且具体地指出通过引用并入并且在本文中被整体阐述。本说明书中对任何现有技术的提及不是并且不应被视为承认现有技术形成了任何国家中所涉及领域中公知常识的一部分。
[0290]
除非本文中另有指示或明显与上下文相矛盾,否则在描述本公开的方法和组合物的上下文中(特别是在以下权利要求的上下文中)使用的术语“一个/种(a/an)”和“所述(the)”以及类似的指代词应解释为涵盖单数和复数两者。除非另外指出,否则术语“包括”、“具有”、“包含”和“含有”应解释为开放式术语(即,意味着“包含但不限于”)。术语“基本上由
……
组成”将组合物、过程或方法的范围限制在特定的材料或步骤,或者限制在那些对组合物、过程或方法的基本和新颖特性没有实质性影响的材料或步骤。替代方案(例如,“或”)的使用应理解为意指替代方案中的一个、两个或其任何组合。如本文所使用的,术语“约”是指所指定范围、值或结构的
±
20%,除非另有指示。
[0291]
除非本文中另有指示,否则本文中对值的范围的叙述仅旨在用作单独地指出落入所述范围内的每个单独的值的速记方法并且每个单独的值并入本说明书中,如同所述值在本文中被单独地叙述一样。例如,除非另有指示,否则任何浓度范围、百分比范围、比率范围、整数范围、大小范围或厚度范围应被理解为包含所叙述范围内的任何整数的值并且在适当的情况下包含其分数(如整数的十分之一和百分之一)。
[0292]
除非本文中另有指示或明显与上下文相矛盾,否则本文所描述的所有方法均可以以任何合适的顺序进行。除非另外声明,否则对本文所提供的任何和所有实例或示例性语言(例如,“如”)的使用仅旨在更好地阐明本公开的方法和组合物并且不对所公开的方法和组合物的范围构成限制。说明书中的任何语言均不应被解释为将任何未要求保护的要素指示为是实践所公开方法和组合物所必需的。
[0293]
本文描述了本公开的优选实施例。在阅读前文描述后,对本领域的普通技术人员而言,那些优选实施例的变型可以变得显而易见。发明人预期技术人员在适当时采用此类变型,并且发明人打算使所公开的方法和组合物以与本文具体描述的方式不同的其它方式来进行实践。因此,在适用法律允许的情况下,本公开包含所附权利要求中叙述的主题的所有修改和等同物。此外,除非另外在本文中指示或另外明确地与上下文相矛盾,否则所公开的方法和组合物涵盖上文描述的要素在其所有可能的变型中的任何组合。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1