IL10受体结合分子和使用方法与流程
背景技术:
1、细胞因子和生长因子配体通常通过细胞表面受体亚基的多聚化发出信号。在一些情况下,细胞因子充当多特异性(例如,双特异性或三特异性)配体,其促进此类受体亚基的缔合,使它们的胞内结构域接近,从而可以发生胞内信号转导。细胞因子的性质决定了哪些受体亚基缔合以形成细胞因子受体复合物。因此,细胞因子将单个受体亚基桥接为受体复合物,从而导致胞内信号转导。
2、细胞因子受体亚基的胞内结构域具有富含脯氨酸的jak结合结构域,其通常位于细胞膜内表面附近的细胞因子受体亚基的胞内结构域的盒1/盒(box1/box)区域。胞内jak激酶与jak结合结构域关联。当胞内结构域受体亚基接近时,通常是通过受体的同源配体与受体亚基的胞外结构域的结合,jak相互磷酸化。已在哺乳动物细胞中鉴定出四种janus激酶:jak1、jak2、jak3和tyk2。ihle等.(1995)nature 377(6550):591-4,1995;o'shea和plenge(2012)immunity 36(4):542-50。jak的磷酸化诱导jak的构象变化,提供进一步磷酸化其他胞内蛋白质的能力,其启动导致多种胞内因子激活的级联反应,所述胞内因子转导与受体相关的胞内信号,从而导致胞内反应(例如基因转录),通常称为下游信号转导。在许多情况下,被jak磷酸化的蛋白质是信号转导和转录激活因子(stat)蛋白家族的成员。迄今为止,已鉴定出哺乳动物stat家族的七个成员:stat1、stat2、stat3、stat4、stat5a、stat5b和stat6。delgoffe等,(2011)curr opin immunol.23(5):632-8;levy和darnell(2002)natrev mol cell biol.3(9):651-62以及murray,(2007)j immunol.178(5):2623-9。激活的jak和stat蛋白的选择性相互作用,统称为jak/stat途径,提供了响应细胞因子结合而观察到的多种胞内反应。
3、人基因组编码大约四十种不同的jak/stat细胞因子受体。原则上,可以产生大约1600种独特的同源二聚体和异二聚体细胞因子受体对,并有可能通过不同的jak/tyk/stat组合发出信号(bazan,proc natl acad sci u s a.87(18):6934-8,1990;huising等,jendocrinol.189(1):1-25,2006)。然而,就目前所知,人基因组编码的不同细胞因子配体少于50种(bazan,proc natl acad sci u s a.87(18):6934-8,1990;huising等,jendocrinol.189(1):1-25,2006),将细胞因子受体复合物的范围限制在那些可由天然配体组装的复合物。鉴于细胞因子配体与细胞因子受体亚基的胞外结构域的相互作用决定了受体复合物中受体亚基的组成,并且细胞内jak/tyk和rtk酶是简并的,细胞因子和生长因子受体二聚体天然发生配对的数量仅代表系统理论上允许的信号转导-能力型受体配对总数的一小部分。
4、天然存在的细胞因子配体介导多种细胞反应。在一些情况下,异源多聚细胞因子受体由受体复合物特有的一个或多个受体亚基构成,称为“专有”亚基,其与多个细胞因子受体共享的其他受体亚基相互作用,通常称为“共有”受体亚基。例如,il7受体是il7ra专有亚基和cd132亚基的异二聚体受体复合物,cd132亚基也称为“共有γ”亚基,因为它是多种细胞因子受体复合物(包括il2、il4、il19、il15和il21)的共享受体亚基。细胞因子与受体亚基的ecd相互作用的相对亲和力和动力学以及响应于细胞因子结合而形成的复合物的稳定性介导胞内信号转导的性质和强度。在一些情况下,细胞因子与专有亚基的结合增强了完整受体的形成,其中细胞因子对共有亚基的亲和力在不与专有亚基缔合时可能会显著降低。
5、细胞因子配体和受体亚基之间相互作用的细微差别是一个重要的科学研究问题。例如,天然存在的细胞因子的许多特性表明,它们在人疾病的治疗中具有潜在用途,但此类天然存在的细胞因子也可能引发不利和不良影响。在许多情况下,疾病与表达潜在治疗性细胞因子受体的具体细胞类型有关。然而,细胞因子受体也表达在其他不希望成为治疗干预靶标的细胞类型上。天然配体的给予激活两种细胞类型,导致不期望的副作用。
6、为了尝试产生能够选择性激活所需细胞类型的细胞因子类似物,已经产生了各种工程改造的细胞因子配体(或其组分),以选择性地调节它们与受体亚基的胞外结构域的亲和力。这些努力已经产生了细胞因子变体,这些变体已显示可提供部分活性,其导致配体的有益特性与不期望影响的解偶联。参见,例如,mendoza等,(2019)567:56–60。然而,此类选择性细胞因子配体的工程改造是基于对配体和受体界面处个体氨基酸残基的选择性调节。这种用于调节细胞因子受体亲和力的蛋白质工程方法需要受体与细胞因子相互作用的三维(通常是x射线晶体学)图谱,以鉴定与受体亚基相互作用的细胞因子残基。此外,这些界面残基处的氨基酸取代的影响可以是高度可变的,通常需要大量耗时的反复试错来确认产生所需活性概况所需的特定氨基酸取代。然而,即使实现了具有所需信号转导概况的工程改造的细胞因子,许多蛋白质对氨基酸取代高度敏感,导致重组表达出现显著问题,无论是在哺乳动物表达系统还是原核系统中,此类氨基酸取代表达于包涵体中时都可以影响蛋白质重折叠。当细胞因子本身是异二聚体时,上述问题尤为突出。例如,il12是由p35和p40亚基组成的异二聚体。在与受体亚基的界面(例如p40/il12rb2界面)处对细胞因子亚基配体工程改造的一个亚基界面的一个残基进行工程改造可能会导致细胞因子配体亚基(例如p35和p40)的缔合问题。此外,il12的p40亚基作为单体和同二聚体发挥作用,其活性也可能受到此类氨基酸取代的影响。因此,本领域需要提供具有细胞因子活性的分子,其可以很容易地生成和工程改造以获得受体亲和力,并且不具有独立的脱靶活性。
7、附图简要说明
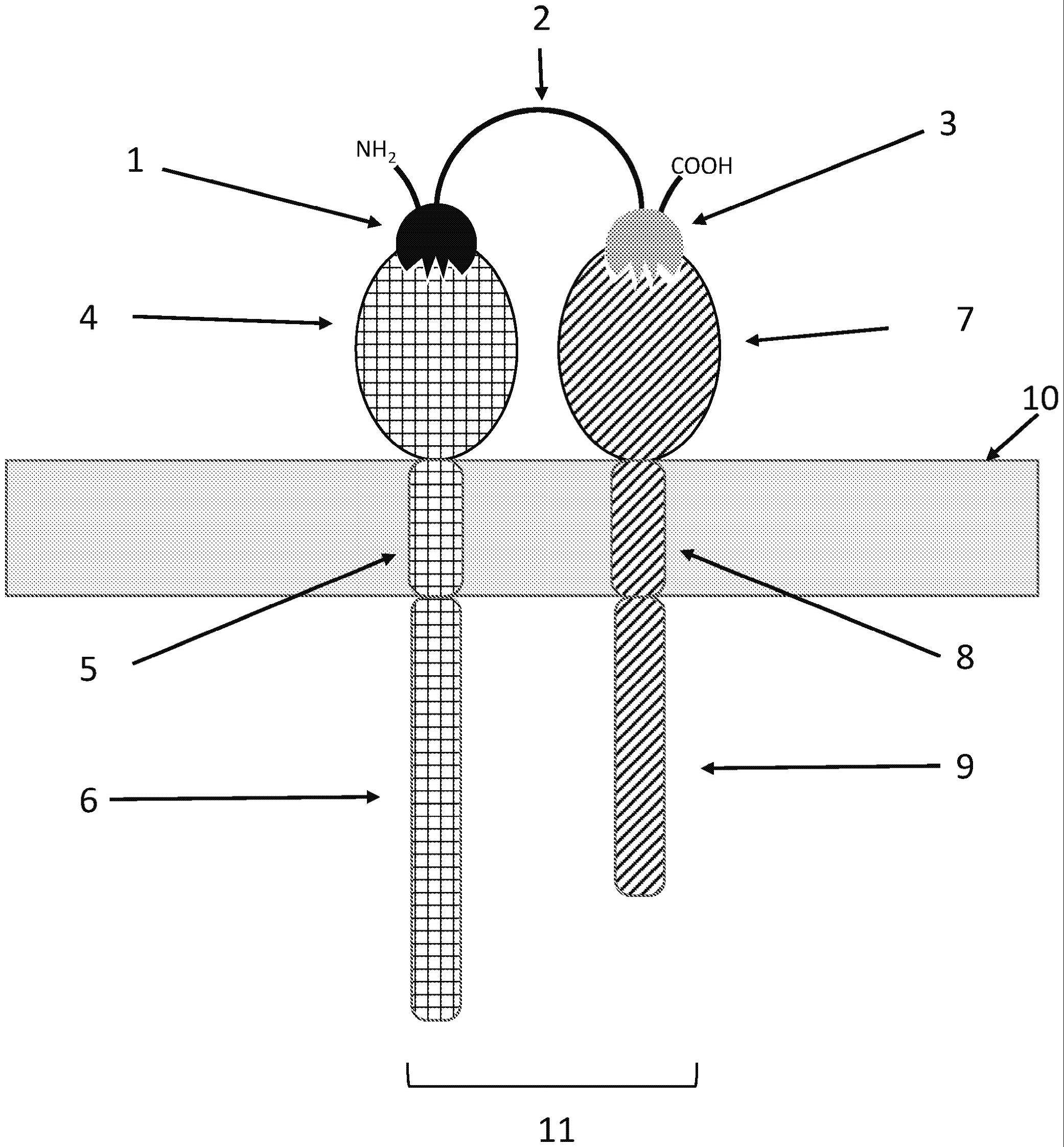
8、附图的图1提供了本公开的结合分子的一个实施方式的示意图,其包含第一单域抗体(1)和第二单域抗体(3)以及接头(2),其描述为与细胞膜(10)相关的异二聚体受体相互作用,该异二聚体受体包含含有胞外结构域(4)、跨膜结构域(5)和与结合分子相互作用的胞内结构域(6)的第一受体亚基,以及包含含有胞外结构域(7)、跨膜结构域(8)和胞内结构域(9)的第二第一受体亚基,其中结合分子的第一受体(6)的胞内结构域和第二受体(9)的胞内结构域在接近距离(11)内。
9、附图的图2提供了本公开的结合分子的两种说明性构型的示意图。分图a提供了说明性单条多肽链结合分子的示意图,其包含从氨基到羧基的第一单域抗体(1)和第二单域抗体(3)和接头(2)。分图b提供了包含第一单域抗体(1)和第二单域抗体(3)和接头(2)和杵臼(knob-into-hole)fc结构域的结合分子的示意图,其包含作为fc杵(13)的第一亚基和作为fc臼(14)的第二亚基,其中单域抗体通过igg铰链序列(12)与fc结构域稳定缔合。
10、附图的图2提供了本公开的结合分子的两种说明性构型的示意图。分图a提供了包含第一单域抗体(1)和第二单域抗体(3)和接头(2)的说明性结合分子的示意图。分图b提供了包含两条多肽链的结合分子的示意图,第一多肽链包含(从氨基到羧基)第一单域抗体(1)、接头序列、第二单域抗体(3)、igg铰链序列(12)和fc杵结构域(13),第二多肽包含fc臼(14),其中第一和第二多肽通过杵臼fc结构域的相互作用稳定缔合。
11、附图的图3提供了本公开的结合分子的两种说明性构型的示意图。分图a提供了一个示例性结合分子构建体的示意图,该结合分子构建体包含两个结合分子,每个结合分子都连接到杵臼fc结构域的亚基上,该构建体包含两条多肽链,第一多肽链(从氨基到羧基)包含第一单域抗体(1)、接头(2)和第二单域抗体(3)、igg铰链序列(12)和fc杵亚基(13),第二多肽链(从氨基到羧基)包含第一单域抗体(1)、接头(2)和第二单域抗体(3)、igg铰链序列(12)和fc臼亚基(14),其中第一和第二多肽通过杵臼fc结构域的相互作用稳定缔合。分图b提供了结合分子构建体的替代排列的示意图,该结合分子构建体包含两条多肽,第一多肽链(从氨基到羧基)包含第一单域抗体(1)、接头(2)和第二单域抗体(3)、igg铰链序列(12)和fc杵亚基(13),第二多肽链(从氨基到羧基)包含第一第二域抗体(3)、接头(2)和第一单域抗体(1)、igg铰链序列(12)和fc臼亚基(14),其中第一和第二多肽通过杵臼fc结构域的相互作用稳定缔合。
12、图4,分图a提供了本公开的结合分子的构型的替代示意图,其中一个单域抗体连接至包含两个多肽的杵臼fc结构域的每个亚基,第一多肽从氨基到羧基包含第一单域抗体(1)、igg铰链序列(12)和fc杵亚基(13),第二多肽从氨基到羧基包含第二单域抗体(3)、igg铰链序列(12)和fc臼亚基(13),其中第一和第二单域抗体通过杵臼fc结构域的相互作用稳定缔合。
13、图4,分图b提供了结合分子的示意图,结合结构域是通过过渡金属配位共价复合物缔合的单域抗体。如图所示,结合分子包含两个多肽亚基:第一亚基包含第一单域抗体(1)的通过第一接头(15)连接至第一螯合肽(17),第二亚基包含第二单域抗体(3)的通过第二接头(16)连接至第二螯合肽(18),其中第一螯合肽(17)和第二螯合肽(18)与单个过渡金属离子(“m”)形成配位共价复合物。过渡金属离子可以处于动力学不稳定或动力学惰性氧化状态。
14、附图的图5提供了用于评估il10r结合分子在分离自pbmc的单核细胞、cd4t细胞和cd8 t细胞中的活性受接头长度影响的方案示意图。
15、附图的图6提供了来自评估il10r多肽结合分子和诱导il10活性的能力的数据,如磷酸化stat3(y轴)和x轴上不同浓度的il10r结合分子测试品所测,以评估il10ra和il10rbsdab之间距离的变化对il10活性的影响。在所有实验中,多肽il10r结合分子的n端il10rasdab是dr241。在分图a中,c端il2rb sdab是dr244。在分图b中,c端il2rb sdab是dr246。在分图c中,c端il2rb sdab是dr247。各种线条代表il10r结合分子,其中il10ra和il10rbsdab之间的距离随接头长度而变化。
16、发明概述
17、本公开提供了可用于细胞受体的配对以产生可用于治疗哺乳动物对象疾病的所需效果的组合物。
18、本公开提供了至少包含特异性结合第一受体亚基的第一结构域和特异性结合第二受体亚基的第二结构域的组合物,使得在与表达第一和第二受体的细胞接触时,该组合物引起第一和第二受体的功能性缔合,从而引发它们的相互作用并导致下游信号转导。在一些实施方式中,第一和第二受体响应同源配体结合而发生接近,并且在本文中被称为“天然”细胞因子受体对。
19、在一个实施方式中,本公开提供结合分子,其包含结合il10受体的il10ra的第一结构域和结合il10受体的il10rb的第二结构域,使得在与表达il10ra受体和il10rb受体的细胞接触后,il10r结合分子引起il10ra和il10rb的功能性缔合,从而导致受体和下游信号转导的功能性二聚化。
20、本公开提供公开提供作为细胞因子受体的配体的细胞因子受体结合分子,所述细胞因子受体结合分子包含:
21、(a)特异性结合细胞因子受体的第一亚基胞外结构域的第一单域抗体(sdab);和
22、(b)特异性结合细胞因子受体亚基的第二亚基的胞外结构域的第二单域抗体;
23、其中:
24、·第一sdab和第二sdab稳定缔合;
25、·响应于与细胞因子受体的同源配体接触,细胞因子受体的第一和第二亚基二聚化;和
26、·将表达细胞因子受体的第一和第二亚基的细胞与有效量的细胞因子受体结合分子接触,导致细胞因子受体的第一和第二亚基的胞内结构域接近并导致胞内信号转导。
27、本公开的il10r结合分子包含两个或更多单域抗体,其选择性结合il10ra和il10rb受体亚基的胞外结构域。在一个实施方式中,本公开提供了一种il10受体(il10r)结合分子,其为il10r受体的配体,该il10r受体结合分子包含:
28、(c)特异性结合il10受体的il10ra亚基胞外结构域的第一单域抗体(sdab)("il10ra sdab"),和
29、(d)特异性结合il10受体的il10rb亚基胞外结构域的第二单域抗体(sdab)("il10rb sdab"),
30、其中:
31、·第一sdab和第二sdab稳定缔合;
32、·il10受体的il10ra和il10rb亚基响应于与il10r结合分子的接触而二聚化;和
33、·将表达il10ra和il10rb的细胞与有效量的il10r结合分子接触导致il10ra和il10rb的胞内结构域接近和胞内信号转导。
34、在一个实施方式中,细胞因子受体结合分子是il10受体的配体并且包含特异性结合il10ra受体亚基胞外结构域的第一sdab和特异性结合il10rb受体亚基胞外结构域的第二sdab。
35、在一些实施方式中,抗il10rαsdab是vhh抗体(抗il10rαvhh抗体)和/或抗il10rβsdab是vhh抗体(抗il10rβvhh抗体)。在一些实施方式中,抗-il10rαsdab和抗-il10rβsdab通过肽接头连接。在一些实施方式中,肽接头包含1至50个氨基酸。
36、在另一个方面,本公开提供了一种在有此需要的对象中治疗肿瘤疾病(例如癌症)的方法,包括向对象给予本文所述的il10r结合蛋白,其中il10r结合蛋白结合并激活cd8+t细胞、cd4+t细胞、巨噬细胞和/或treg细胞。在一些实施方式中,il10r结合蛋白提供比聚乙二醇化il10更长的治疗功效。在一些实施方式中,癌症是实体瘤癌症。
37、在其他方面,本文所述的il10r结合蛋白也可用于治疗炎性疾病(例如克罗恩病和溃疡性结肠炎),以及自身免疫性疾病(例如银屑病、类风湿性关节炎和多发性硬化症)。
38、在一些实施方式中,结合分子的一个sdab是scfv,另一个sdab是vhh。
39、在一些实施方式中,第一和第二sdab通过化学连接共价结合。
40、在一些实施方式中,第一和第二sdab作为单条连续多肽提供。
41、本发明提供了il10r结合分子,其为il10受体的合成配体。
42、在一些实施方式中,本发明的il10r结合分子是下式[#1]的多肽:
43、h2n-(il10 vhh#1)–(l1)a–(il10 vhh#2)–(l2)b-(cp)c-cooh [#1]
44、其中:“—”代表共价键;l1和l2是接头;cp是螯合肽;a、b和c独立地选自整数0或1;“h2n”表示氨基末端;并且“cooh表示多肽的羧基末端”
45、在一些实施方式中,本发明提供的il10r结合分子是下式[#1]的多肽,其中il10vhh#1是ilra sdab,il10 vhh#2是il10rb sdab,il10 vhh#1是ilrb sdab并且il10 vhh#2是il10ra sdab。
46、在一些实施方式中,本发明提供了包含il10ra sdab的il10r结合分子,其包含:
47、·相对于seq id no:1、4、7、10、13、16、19、22、25、28、31、34、37、40、43、46、49和52中任一个的序列,cdr1具有至少90%(例如,91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列同一性,或具有0、1、2或3个氨基酸变化,任选保守氨基酸变化;
48、·相对于seq id no:2、5、8、11、14、17、20、23、26、29、32、35、38、41、44、47、50和53中任一个的序列,cdr2具有至少90%(例如,91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列同一性,或具有0、1、2或3个氨基酸变化,任选保守氨基酸变化;和
49、·相对于seq id no:3、6、9、12、15、18、21、24、27、30、33、36、39、42、45、48、51和54中任一个的序列,cdr3具有至少90%(例如,91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列同一性,或具有0、1、2或3个氨基酸变化,任选保守氨基酸变化。
50、在一些实施方式中,本发明提供了包含il10rb sdab的il10r结合分子,其包含:
51、·相对于seq id no:55、58、61、64、67、70、73、76、79、82、85、88、91、94、97、100、103、106、109、112、115、118、121、124、127、130、133、136、139、142、145、148和151中任一个的序列,cdr1具有至少90%(例如,91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列同一性,或具有0、1、2或3个氨基酸变化,任选保守氨基酸变化;
52、·相对于seq id no:56、59、62、65、68、71、74、77、80、83、86、89、92、95、98、101、104、107、110、113、116、119、122、125、128、131、134、137、140、143、146、149和152中任一个的序列,cdr2具有至少90%(例如,91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列同一性,或具有0、1、2或3个氨基酸变化,任选保守氨基酸变化;和
53、·相对于seq id no:57、60、63、66、69、72、75、78、81、84、87、90、93、96、99、102、105、108、111、114、117、120、123、126、129、132、135、138、141、144、147、150和153中任一个的序列,cdr3具有至少90%(例如,91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列同一性,或具有0、1、2或3个氨基酸变化,任选保守氨基酸变化。
54、在一些实施方式中,本发明提供il10r结合分子,相对于seq id no:154-171中的任一个,其包含至少90%(例如91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列同一性的il10ra sdab,或具有0、1、2或3个氨基酸变化,任选保守氨基酸变化。
55、在一些实施方式中,本发明提供il10r结合分子,相对于seq id no:172-198或seqid no:199-201中的任一个,其包含至少90%(例如91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列同一性的il10rb sdab,或具有0、1、2或3个氨基酸变化,任选保守氨基酸变化。
56、在一些实施方式中,本发明提供il10r结合分子,相对于seq id no:256-353中的任一个,其包含至少90%(例如91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列同一性,或具有0、1、2或3个氨基酸变化,任选保守氨基酸变化。
57、在一些实施方式中,本发明提供il10r结合分子,相对于seq id no:172-198或seqid no:199-201中的任一个,其包含具有至少90%(例如91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列同一性的il10rb sdab,或具有0、1、2或3个氨基酸变化,任选保守氨基酸变化。
58、在一些实施方式中,本发明提供与表29的任何一种il10r结合分子的序列(seq idno:[dr1511-dr1525])具有至少90%(例如91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列同一性的il10r结合分子。在一些实施方式中,本发明提供与表29的任何一种il10r结合分子(seq id no:[dr1511-dr1525])基本同一的il10r结合分子。在一些实施方式中,本发明提供与表29的任何一种il10r结合分子(seq id no:[dr1511-dr1525])的序列同一的il10r结合分子。
59、在一个实施方式中,本公开提供了一种il10ra结合分子,其相对于单核细胞优先激活t细胞,特别是cd8+t细胞。在一个实施方式中,本公开提供了式#1的il10ra结合分子,其中il10ra sdab对il10ra胞外结构域的亲和力高于il10rb sdab对il10rb胞外结构域的亲和力。
60、在一些实施方式中,本公开提供经修饰的il10r结合分子以在哺乳动物对象体内提供延长的作用时间,及其药学上可接受的制剂。在一些实施方式中,本发明提供聚乙二醇化的式#1的il10r结合分子,其中peg偶联至il10r结合分子,并且peg是直链或支链peg分子,其具有约2,000至约80,000道尔顿、或约2,000至约70,000道尔顿、或约5,000至约50,000道尔顿、或约10,000至约50,000道尔顿、或约20,000至约50,000道尔顿、或约30,000至约50,000道尔顿、或20,000至约40,000道尔顿或约30,000至约40,000道尔顿分子量。在本公开的一个实施方式中,peg为包含两个20kd臂的40kd支链peg。
61、本公开的il10r结合分子可用于治疗或预防哺乳动物对象的疾病。在一些实施方式中,本公开通过给予本公开治疗有效量的il10r结合分子来治疗或预防哺乳动物对象的自身免疫性疾病。在一些实施方式中,本公开通过给予本公开治疗有效量的il10r结合分子来治疗或预防哺乳动物对象的传染性疾病(包括病毒和慢性病毒感染)。在一些实施方式中,本公开通过给予本公开治疗有效量的il10r结合分子来治疗或预防哺乳动物对象的肿瘤性疾病。在一些实施方式中,本公开通过给予本公开治疗有效量的il10r结合分子与一种或多种辅助治疗剂的组合来治疗或预防哺乳动物对象的肿瘤、传染性或自身免疫。
62、本公开还提供了用于向哺乳动物对象给予的il10r结合分子的药学上可接受的制剂。本公开还提供了一种药学上可接受的组合物,用于向哺乳动物对象给药,该组合物包含编码多肽il10r结合分子的核酸序列、编码多肽il10r结合分子的重组病毒或非病毒载体、或包含编码多肽il10r结合分子的核酸序列的重组修饰的哺乳动物细胞,在每种情况下该核酸序列操作性地连接至在哺乳动物细胞中有功能的一种或多种表达控制元件。
63、本公开提供了编码多肽il10r结合分子的核酸序列。本公开还提供了包含编码多肽il10r结合分子的核酸序列的重组载体。本公开还提供了一种重组修饰的哺乳动物细胞,其包含编码多肽il10r结合分子的核酸。本公开还提供了重组载体的多肽il10r结合分子的重组产生、分离、纯化和表征的方法,其包含并提供了编码多肽il10r结合分子的核酸序列。
64、本公开还提供了一种表达载体,其包含编码操作性地连接至一个或多个表达控制序列的双特异性结合分子的核酸。本公开还提供了一种分离的宿主细胞,其包含表达载体,表达载体包含编码双特异性结合分子的核酸,其操作性地连接至在宿主细胞中有功能的一个或多个表达控制序列。
65、在另一个方面,本公开提供了一种药物组合物,其包含本文所述的il10r结合分子和药学上可接受的运载体。
66、在另一个方面,本公开提供了一种在有需要的对象中治疗自身免疫或炎性疾病、病症或病况或病毒感染的方法,包括向对象给予治疗有效量的本文所述的il10r结合分子或本文所述的药物组合物。
67、本文描述的结合分子有几个优点。il10受体的天然配体il10导致il10ra和il10rb接近(即通过它们对il10的同时结合)。然而,当il10在哺乳动物(特别是人)对象中被用作治疗剂时,它也可能通过多种机制引发许多不利和不良影响,包括il10ra和il10rb在其他细胞类型上的存在以及与其他细胞类型上的il10ra和il10rb的结合可能会对表达il10ra和il10rb的细胞产生不良影响和/或不期望的信号转导。本公开内容涉及调节il10ra和il10rb结合的多重效应的方法和组合物,以便发生所需的治疗信号转导,特别是在所需的细胞或组织亚型中,同时最小化不期望的活性和/或胞内信号转导。
68、在一些实施方式中,本文所述的il10r结合分子是il10受体的部分激动剂。在一些实施方式中,本文所述的结合分子被设计为使得结合分子是完全激动剂。在一些实施方式中,本文所述的结合分子被设计为使得结合分子是超级激动剂。
69、在一些实施方式中,结合分子通过与所需细胞类型上的il10ra和il10rb的结合提供最大所需的il10胞内信号转导,同时对其他不期望的细胞类型提供显著更少的il10信号转导。例如,这可以通过选择具有不同亲和力,或与il10对il10ra和il10rb的亲和力相比,对il10ra和il10rb造成不同的e最大的结合分子来实现。因为不同的细胞类型以不同的灵敏度响应配体与其同源受体的结合,通过调节二聚体配体(或其单个结合部分)对il10受体相对于野生型il10结合的亲和力促进所需活性的刺激同时减少对非靶标细胞的不期望的活性。
技术实现思路
- 还没有人留言评论。精彩留言会获得点赞!