生物合成方法
生物合成方法
1.本发明一般涉及通过细菌细胞生产产品的方法,具体涉及工业生物技术或微生物学领域,其中活细胞用于生产有用的化学品和产品,例如生物制品。更具体地,本发明涉及一种通过细菌细胞生产产品的方法,该方法包括不可逆地使细菌细胞中的复制起点失活的步骤。本发明还涉及包含复制起点的细菌细胞,其中该细菌细胞已被修饰,使得复制起点可以不可逆地失活。本发明还涉及缺乏功能性复制起点序列的经修饰的细菌细胞,以及经修饰的细菌细胞的各种载体和用途。
2.工业生物技术使用活细胞(主要是微生物)来生产有用的化学品和材料。其源于传统的生物技术,因为几千年来人们一直使用微生物来生产酸奶、啤酒和腌菜。最近,由于基因工程和合成生物学的进步,可能产品的选择范围大幅扩大。
3.科学文献中有通过微生物生产数百种不同化学品的报告,但其中只有一小部分达到了商业生产。主要问题是许多候选菌株不能生产足够的产品;即效价、产量和/或生产率太低而无法实现有利可图的商业生产(van dien,2013)。低生产的根本原因是微生物的生长和产品的生产之间存在权衡(venayak et al.,2015)。一般而言,微生物可以利用其可用的碳和能源通过生产生物量来生长,或合成目标产品。如果更多的资源被分配用于生长,生产就会受到影响,反之亦然。这种生长与生产的权衡严重限制了生产菌株的开发,并限制了可以商业生产的产品清单。
4.有人提议将两阶段生物过程作为绕过这种权衡的一种方式(burg et al.,2016)。在两阶段生物过程中,生物过程分为两个阶段——首先细胞以最大速率生长且不进行(显著)生产,当积累了足够的生物量时,关闭生长并诱导产品生产。有几个详细描述的系统可用于诱导产品合成所需的基因表达并诱导产品生产(marschall,sagmeister and herwig,2017),但关闭生长更难实现。迄今为止所描述的在两阶段生物过程中关闭生长的解决方案依赖于代谢途径的重排(soma et al.,2014)(brockman and prather,2015)。在从生长到生产的转换过程中,生长所需的代谢途径被关闭,积累的代谢中间体被引向目标产品。然而,这些解决方案是产品特异性的,必须为每种新产品设计和构建停止生长的新途径。因此,目前缺乏一种将细菌从生长阶段有效地转换到生产阶段的通用方法。
5.本发明人开发了一种新颖的两阶段生物过程,从而开发了一种新颖的通过细菌生产产品的方法。其优势在于,本发明的解决方案是通用的,从某种意义上它可应用于所有细菌,并且独立于任何目标代谢途径(例如,它独立于任何与生产所需产品有关的目标代谢途径)。它基于细菌染色体的复制起点的不可逆失活。令人惊奇的是,这不可逆地停止了细胞的分裂和生长,却保持了细胞的代谢活性。
6.本发明的可能应用有很多。它可用于微生物生物生产,以提高产品(例如蛋白质和小分子产品)的效价、产量和生产率。它还可用于生成具有固定菌株比例的微生物聚生体或微生物共培养物,因为非分裂细胞会相互促进生长。当途径在两个(或更多)菌株或菌种之间分割时,此类聚生体可用于生产产品。在这种情况下,第一个菌株产生中间体分子,然后由第二个(或更多)菌株转化为最终产品(wang et al.,2018)(yuan et al.,2020)。使用本发明,这些菌株的生长期可以单独进行,并且可以在每个菌株的复制起点不可逆地失活后,
例如在细胞生长和分裂完成后,通过将菌株按预定比例混合在一起来创建聚生体。或者,可以在每个菌株的复制起点不可逆地失活前,例如在细胞生长和分裂开始或完成前将菌株混合。
7.生长和生产阶段的分离也将使其在不同的时间或设施进行。可以在一个设施中培养细胞,然后将其在同一地点或运送到其他地方储存以供以后使用。
8.本发明也可用于工业微生物学以外的领域。在活体治疗领域,将活体微生物细胞作为治疗剂施用于患者,以产生必要的分子或分解有害的分子。这些活体治疗剂应具有代谢活性以实现其功能,但应避免其增殖失控。本发明具有这种双重功能。
9.本发明的方法促进并涉及从基因组中永久去除复制起点或使复制起点失活。一旦复制起点被去除或失活,基因组就不能再起始复制。由于转换(例如复制起点切除)效率非常高,研究人员可以精确地选择在诱导产品生产前细胞培养应达到的最终细胞密度(参见例如图3)。
10.本发明中使用的复制起点去除或失活的永久性的另一个优点是,不需要维持将细菌从生长期转换到生产期所需的条件。因此,一旦复制起点被去除(或删除或以其他方式不可逆地失活),细胞可以恢复到诱导前条件(或实际上任何其他条件),而不需要其再次恢复生长。这为操作人员提供了更多自由,并且不会限制可能的生产条件。
11.本技术的一个意想不到的优点是,已转换细胞即使在高培养密度下也能持续生产产品。图4b显示,在对照和转换菌株(转换后)同时(约10小时)达到相同的最终细胞密度的实验装置中,已转换细胞中的蛋白生产持续时间远远超过对照细胞。在对照细胞中,于8h的时间点停止红色荧光蛋白(rfp)的产生,此后不再积累rfp。然而,在转换细胞中(转换后),rfp信号持续增加到14h,导致与对照相比,rfp水平增加高达400%。尽管两种培养物同时达到相同的最大细胞密度,但仍会发生这种情况。
12.本发明具有广泛的可能应用范围。例如,它可用于生成人体治疗用途的非分裂细胞制剂。在此类应用中,复制起点的永久去除或失活至关重要,因为当细胞制剂在人体内时,它允许复制起点的失活状态的维持。这对于防止体内细胞不必要的生长的同时仍允许产品(治疗产品)生产并因此达到治疗效果很重要。
13.本发明还可以应用于生物防护领域,防止细胞增殖失控,这些细胞可能意外地从例如生物反应器(或其他受限环境)逃逸至自然(或非受限)环境(lee et al.,2018)。由于本发明的已转换细胞不能增殖,因此如果从受限(例如生物反应器)环境中释放,其构成的威胁会减少。相比之下,例如,暂时受阻的细胞一旦摆脱了诱导条件就可以恢复生长。
14.一方面,本发明提供了一种通过细菌细胞生产产品的方法,该方法包括不可逆地(或永久地)使细菌细胞中的复制起点失活的步骤。
15.术语“复制起点”是本领域的术语,并且技术人员可以理解。它一般指任何序列,例如核苷酸序列,在该序列上可以起始dna复制,例如通过结合适当的蛋白和酶,这些蛋白和酶是复制机制的一部分或复制过程所必需的。特定细菌菌株中的复制起点在本领域中可以用特定名称来指代。例如,在大肠杆菌染色体中发现的复制起点称为oric,并且在一些实施方案中优选该复制起点的不可逆失活。复制起点可以存在于染色体上,也可以存在于染色体外遗传物质上,例如附加体、质粒或多核苷酸载体。通常,复制起点位于染色体上。
16.术语“附加体”、“质粒”和“多核苷酸载体”是指通常以环状双链dna片段形式存在
的染色体外元件。此类元件可以是来自任何来源的线性或环状单链或双链dna或rna的自主复制序列、基因组整合序列、噬菌体或核苷酸序列,其中一些核苷酸序列被连接或重组为独特结构,其能够将启动子片段和目标或相关dna序列,例如对于选定的基因产物,连同适当的3’非翻译序列引入细胞。
17.本文所用的术语“不可逆地失活”或“永久地失活”复制起点或类似术语是指复制起点不可逆地(或永久地)不能充当dna复制的起始位点,或以其他方式作为功能上失活的复制起点。
18.类似地,本文所用的术语“不可逆失活”或“永久失活”的复制起点是指具有活性或功能但可以(或能够)以不可逆(或永久)的方式失活的复制起点。
19.类似地,本文所用的术语“不可逆去除”或“永久去除”的复制起点是指具有活性或功能但可以(或能够)以不可逆(或永久)的方式去除的复制起点。
20.在实施方案中,不可逆地(或永久地)使复制起点失活可通过永久改变,优选复制起点的永久遗传改变(或永久核苷酸改变)来实现。不可逆(或永久)失活的复制起点是指不再作为复制起点发挥作用的复制起点,例如不再用作起始dna复制的位点。
21.在实施方案中,不可逆地(或永久地)使细菌细胞中的复制起点失活包括或由复制起点的遗传修饰(或核苷酸修饰)组成。
22.在实施方案中,遗传修饰(或核苷酸修饰)包括或由复制起点的突变组成,使其不再作为复制起点发挥作用。
23.在实施方案中,复制起点的突变包括通过从复制起点插入、去除或替换核苷酸而引起的复制起点的突变。示例性突变或修饰或其他改变是指那些需要与复制起点相互作用以起始或进行复制的蛋白、酶或其他分子实体不再与复制起点相互作用,或者不再以发生复制,例如可测量的或显著的复制的方式与复制起点相互作用。例如,此类突变等可以是任何减少或抑制或阻止复制起点功能的突变等,例如减少或抑制(例如显著减少或抑制)或阻止复制起点处的dna复制。
24.不同细菌菌株的细菌复制起点的共同特征包括促进dna链分离的富含at的dna解链元件(due),以及作为dnaa结合位点的9个核苷酸长度的元件(所谓的“dnaa盒”)。在dna复制起始过程中,dnaa单体与复制起点内的dnaa盒结合。这些dnaa单体在与dnaa盒结合时,会寡聚化从而形成dnaa复合物。dnaa复合物足以解链due并协助将dna解旋酶(dnab)加载到暴露的单链上,从而完成细菌前复制复合物的形成。因此,本领域通常理解,due和dnaa盒是细菌中dna复制起始所必需的。
25.因此,在实施方案中,复制起点的不可逆(或永久)失活包括突变或其他改变或修饰,使得dnaa单体与复制起点的结合被减少或阻止(例如显著减少或阻止),或due的解旋被减少或阻止(例如显著减少或阻止),或dna解旋酶的结合(例如dnab分子的结合)被减少或阻止(例如显著减少或阻止)。换句话说,突变、改变或修饰使得dna复制不能在复制起点处起始或显著起始,或者使得复制起点失活或基本失活。更具体地,复制起点的不可逆(或永久)失活可包括复制起点的至少一个、至少两个、至少三个、至少四个或所有、或最多两个、三个或四个dnaa盒(或dnaa结合位点)的突变等。
26.在实施方案中,复制起点的不可逆(或永久)失活包括复制起点的至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少11个、至少12个、至少13个、至少14个、至少
15个或所有dnaa盒的突变,或最多5、6、7、8、9、10、11、12、13、14或15个dnaa盒,例如至少或最多13个dnaa盒的突变。在实施方案中,复制起点的不可逆(或永久)失活包括去除(或缺失或消除或切除)复制起点的至少或最多1、2、3、4、5、6、7、8、9、10、11、12、13、14、15或所有dnaa盒,例如至少或最多13个dnaa盒。优选地,该突变为去除(或缺失或消除或切除)。
27.在实施方案中,该产品是在细菌细胞中生产或可由细菌细胞生产的产品(或任何产品或任何目标产品),例如在本领域的两阶段生物过程方法中生产或可由该方法生产。因此,此类产品是希望以提升的、可测量的或显著的量制备或生产的产品,例如是所需的、有用的、工业的或商业的产品,或有价值的产品,例如对工业或研究而言。因此,在实施方案中,该产品是小分子。备选地或另外地,该产品可以是多肽或蛋白或蛋白复合物(或目标蛋白),例如抗体、抗原、酶、生长因子或细胞因子。优选地,该产品是药物化合物、医药产品、食品添加剂、着色剂化合物、香料成分、工业化学品、精细化学品或生物燃料。优选地,生物燃料是醇,例如丁醇或异丁醇。
28.使细菌能够产生所需产品的必要步骤,例如诱导或允许产物合成或调节所必需的基因表达,对于本领域技术人员来说是众所周知的。例如,在该产品是小分子的情况下,可以表达一种或多种酶,其可以从给定的底物生产产品。所述底物可以在细菌细胞中均匀地或内源地获得,例如其中底物是细菌细胞内代谢途径的中间体。备选地或另外地,可以异源地(例如通过在细菌细胞中表达异源基因)或外源地(例如通过向培养基施用)提供所述底物。
29.可以通过增加产品生产的调节蛋白的表达和/或通过消除编码减少产品生产的酶和/或调节蛋白的基因来提高产品生产。这些步骤通常通过向细胞dna中引入变化来进行,例如通过基因组和/或质粒的变化。当该产品是异源蛋白时,通常需要提供编码异源蛋白的核苷酸序列。任选地,还可以提供编码伴侣蛋白和/或其他辅助因子的核苷酸序列以帮助目标蛋白的折叠和加工。当小规模测试从细菌细胞生产产品时,通常首先在质粒或附加体上提供必需的异源基因。然而,对于工业规模而言,它几乎总是并且优选通过基因组表达来完成。
30.在实施方案中,本发明的方法增加或改善了细菌细胞的产品或产品生产(或生产的产品)的水平或量或浓度,优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率。优选地,所述增加是可测量的或显著的增加,例如在统计学或临床上是显著的。举例来说,优选缺乏功能性复制起点序列的本发明的细菌细胞(例如复制起点已不可逆(或永久)失活的本发明的细菌细胞),其可在产品生产或生产的产品的水平方面(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率)或生产所述产品或在生产所述产品中具有积极作用的一种或多种酶的活性方面引起至少或最多1.5倍、2倍、2.5倍、3倍、3.5倍、4倍、4.5倍、5倍、6倍、7倍、8倍、9倍或10倍的增加。可以使用任何适当的比较,例如当不存在或不诱导复制起点的不可逆(或永久)去除时与从相同类型的细菌细胞(例如其中复制起点未不可逆地失活的等效细菌细胞)观察到的所述水平相比,或者与例如如别处所述的对照菌株的所述水平相比增加。
31.本文所用的术语“对照菌株”(或对照细胞)一般可以指没有或不具有复制起点的不可逆(或永久)去除的细菌菌株(等效细菌菌株)。例如,该术语可以指不同于本发明的细菌细胞的细菌菌株(或细菌细胞),因为其不包含能够不可逆(或永久)失活的复制起点,例
如包含原生或野生型或未经修饰的复制起点,例如是原生或野生型或未经修饰的菌株或细胞。或者,当在不存在或不触发/诱导复制起点的不可逆(或永久)失活的条件下使用时,术语“对照菌株”可以表示细菌菌株或细菌细胞(例如本发明的细菌细胞)(例如如本文所定义的转换子或转换细胞,即其复制起点已被修饰以致其可以不可逆地失活(即能够不可逆地失活)但尚未不可逆地失活的经修饰的细菌细胞)。或者,术语“对照菌株”可以表示具有经修饰的复制起点的细菌菌株或细菌细胞(例如上文所述的转换细胞),但其中不能诱导复制起点的不可逆(或永久)失活,例如因为细菌细胞缺乏负责催化失活过程的酶(或所述酶的功能形式)(例如位点特异性重组酶,例如此类对照菌株可能含有缺陷或无功能重组酶,或缺乏活性重组酶)。
32.测量产品生产或生产的产品的水平(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率)或生产所述产品的酶的活性的适当方法对于本领域技术人员来说是众所周知的。因此,在本发明的一些实施方案中,该方法将涉及检测或确定由细菌菌株生产的产品的量或水平(例如浓度)(例如在确定效价或产量时),以及任选地测量由细菌细胞生产的产品的所述量或水平(例如浓度)所需时间的步骤。
33.因此,通常本发明的菌株或在本发明的方法中使用的菌株在转换时(即在复制起点不可逆地(或永久地)失活后)将表现出比上述对照菌株,例如转换菌株更高水平,有时显著更高水平的产品生产或生产的产品(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率)。如果合适,可以通过本领域已知的方法方便地测量或确定产品生产的水平。因此,例如在体外评估时,能够比对照具有更高(增加的)水平或显著更高(增加的)水平的产品生产或生产的产品(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率)的菌株,构成本发明的又一方面。
34.细菌细胞生产的产品可以分泌到培养基(或上清液)中,也可以保留在细胞内。因此,生产或生产的产品的水平可以根据培养基(或上清液)中存在的产品的水平、保留在细胞内的产品的水平(例如这是在标准蛋白提取和纯化后可测量的)、或两者来测量,具体取决于相关产品。
35.换句话说,例如在细菌细胞的培养基(或上清液)中和/或细胞内,例如在体外评估时,能够在转换时具有比对照菌株更高(增加的)水平或显著更高(增加的)水平的产品生产或生产的产品(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率)的菌株,构成本发明的又一方面。例如,菌株能够在转换时在受试者中具有比对照菌株更高(增加的)水平或显著更高(增加的)水平的产品生产或生产的产品(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率),例如在转换时在受试者中具有比对照菌株更高(增加的)局部水平的产品生产或生产的产品,例如在体内评估时,构成本发明的又一方面,特别是在将菌株施用于受试者时观察到这种效果的情况。
36.本发明的细菌细胞或经修饰的细菌细胞可以以任何适当的常规治疗方式施用于受试者,例如使用细菌细胞的治疗,以及用于指定疾病的治疗,包括但不限于口服、舌下、经皮、皮肤、直肠、鼻腔、阴道或眼部给药,或通过吸入给药或通过口腔给药或通过注射给药。此外,本发明的细胞或组合物可以配制用于肠胃外给药,例如通过注射或连续输注。给药途径可以是任何有效地将细菌细胞或经修饰的细菌细胞运送到所需位点而不伤害接受者的
途径。
37.优选地,细菌菌株中(产品生产、或生产的产品、或生产所述产品的酶的活性)的所述更高水平或增加是可测量的或显著的增加,例如在统计学或临床上是显著的。举例来说,在产品或产品生产或生产的产品的水平(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率),例如产品生产或生产的产品的局部或体外水平方面,或生产所述产品的酶的活性水平方面(例如在体外评估时),与上述对照的水平相比可引起至少或最多增加10%、20%、30%、40%、50%、60%、70%、80%、90%、100%、150%、200%、250%、300%、350%、400%、450%、500%、550%或更高的菌株为优选。换句话说,在产品或产品生产或生产的产品的水平(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率),例如产品或产品生产或生产的产品的局部或体外水平方面,或生产所述产品的酶的活性水平方面,与上述对照菌株的水平相比可引起至少或最多增加1.5倍、2倍、2.5倍、3倍、3.5倍、4倍、4.5倍、5倍、6倍、7倍、8倍、9倍或10倍的菌株为优选。可以如本文别处所述方便地测量体外水平,例如在细菌培养物中。优选地,此类增加是细胞外(例如分泌的)和/或细胞内产品水平的增加(优选根据效价或产量)或生产所述产品的酶的活性的增加,例如在体外,例如在细菌培养物的培养基(或上清液)中或在细胞内测量的增加。
38.测量产品或产品生产或生产的产品的水平(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率)或生产所述产品的酶的活性的适当方法是本领域技术人员已知的,并且可以使用其中任何一种方法。
39.在实施方案中,本发明的方法增加了产品或产品生产或通过细菌细胞生产的产品的水平(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率)。优选地,所述增加是可测量的或显著的增加,例如在统计学或临床上是显著的。举例来说,优选的是,当用于本发明的方法时,在产品生产或生产的产品的水平方面(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率),或生产所述产品的酶的活性方面,细菌细胞中复制起点的不可逆(或永久)失活(或细菌细胞中缺乏功能性复制起点序列)引起上述定义的倍数增加或百分比增加。可以使用任何适当的比较,例如与在相同细菌细胞(或相同类型的细菌细胞)中,当细菌细胞中的复制起点未失活(或未不可逆地失活)时观察到的水平相比时,水平(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率)的增加,或与对照细胞,例如本文别处所述的对照细胞的水平相比时,水平(优选根据效价、产量或生产率,或根据每个细胞的效价、每个细胞的产量或每个细胞的生产率)的增加。
40.任选地,可以按细菌细胞数量(或按细菌细胞群体大小或按细菌细胞群体密度)来量化产品或产品生产或细菌细胞生产的产品的水平(优选效价、产量或生产率)。换句话说,可以以考虑到用于生产产品的细菌细胞数量(或细菌细胞群体大小或细菌细胞群体密度)的方式来量化产品或产品生产或细菌细胞生产的产品的水平(优选效价、产量或生产率)。
41.测量细菌细胞群体大小或细菌细胞数量的适当方法在本领域是众所周知的,包括通过测量细菌细胞群体在600nm处的光密度(od
600
)。
42.在实施方案中,本发明的细菌细胞或本发明的方法中使用的细菌细胞在细菌细胞的复制起点不可逆地(或永久地)失活后的一段时间内保持生产产品(或被诱导/刺激以生
产产品)的能力(优选地,与对照细胞,例如本文别处所述的对照细胞相比,这种能力保持更长或显著更长的时间段)。本发明的细菌细胞或本发明的方法中使用的细菌细胞可以在细菌细胞的复制起点不可逆地(或永久地)失活后,或在细菌细胞的不可逆(或永久)失活被触发或诱导后,或在进行了使细菌细胞中的复制起点不可逆地失活的步骤后,可保持生产产品(或被诱导/刺激以生产产品)的能力长达(或至少)3小时、长达(或至少)4小时、长达(或至少)5小时、长达(或至少)6小时、长达(或至少)7小时、长达(或至少)8小时、长达(或至少)9小时、长达(或至少)10小时、长达(或至少)11小时、长达(或至少)12小时、长达(或至少)13小时、长达(或至少)14小时、长达(或至少)15小时、长达(或至少)20小时、长达(或至少)25小时、或长达(或至少)30小时。还优选的是,在任何特定时间点,本发明的细菌细胞或本发明的方法中使用的细菌细胞生产产品的能力大于,例如显著大于如本文别处所述的适当的对照细胞,例如与转换细胞(即复制起点还未失活的本发明的经修饰的细菌细胞)或复制起点不能发生不可逆失活的细胞或未经修饰的细胞或野生型细胞相比。
43.细菌细胞中复制起点的不可逆失活应导致细胞分裂和生长的停止。一旦诱导或触发细菌细胞中复制起点的不可逆失活,细菌细胞培养物逐渐达到明显的细胞密度平台期。可以通过调整细菌细胞培养物的起始细胞密度(例如,接种培养物的细胞密度和/或接种培养物的稀释因子)和/或诱导复制起点的不可逆失活时的细胞密度来选择细胞密度停止增加(即达到平台期)的值。
44.因此,在实施方案中,本发明的或在本发明的方法中使用的细菌细胞培养物的峰值细胞密度(或最大细胞密度,或最高细胞密度,或平台期细胞密度)为至少30%、至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少100%、约100%(或大致相同,或相同)、超过100%、至少100%、至少105%、至少110%、约110%、至少115%、至少120%、至少125%、至少130%、至少135%、至少140%、至少145%、至少150%的对照细菌细胞培养物的峰值细胞密度。还提供了“高达”这些百分比的值。
45.优选地,本发明的或在本发明的方法中使用的细菌细胞培养物的峰值细胞密度(或最大细胞密度,或最高细胞密度,或平台期细胞密度)为至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少100%、约100%(或大致相同,或相同)、超过100%、至少100%、至少105%、至少110%、约110%、至少115%或至少120%的对照细菌细胞培养物的峰值细胞密度。还提供了“高达”这些百分比的值。
46.优选地,本发明的或在本发明的方法中使用的细菌细胞培养物的峰值细胞密度(或最大细胞密度,或最高细胞密度,或平台期细胞密度)为至少90%、至少95%、至少100%、约100%(或大致相同,或相同)、超过100%、至少100%、至少105%、至少110%或约110%的对照细菌细胞培养物的峰值细胞密度。还提供了“高达”这些百分比的值。
47.适当的对照细菌细胞培养物如本文别处所述,并且可以例如是本发明的细菌细胞培养物,其中复制起点的不可逆失活未被诱导(例如复制起点的不可逆失活没有发生或不能发生)。
48.在实施方案中,由本发明的或在本发明的方法中使用的细菌细胞或经修饰的细菌细胞生产的产品水平或生产水平较高,其中细菌细胞培养物或经修饰的细菌细胞培养物的峰值细胞密度(或最大细胞密度,或最高细胞密度,或平台期细胞密度)更接近于对照细菌
细胞培养物的峰值细胞密度,例如上述对照细菌细胞培养物的峰值细胞密度的至少70%等。
49.在实施方案中,本发明的方法增加了细菌细胞生产的产品的产量或效价,或增加了细菌细胞的生产率。
50.术语“效价”意为在生物过程中或生物过程结束时生产的产品的量或浓度。因此,效价可以用“g/l”或任何其他适当的单位来测量。
51.术语“产量”意为提供给细菌用于生产方法或生物过程的每质量底物(例如碳和能源)所生产的产品质量。因此,产量可以用“g/g”或任何其他适当的单位来测量。
52.术语“生产率”与细菌在生物过程中生产产品的速度有关。因此,生产率为每单位时间生产的产品的量或浓度。因此,生产率可以用单位“g l-1
h-1”,即克每升每小时或任何其他适当的单位来测量。
53.在实施方案中,该产品是异源产品。
54.术语“异源”在本领域中是普遍理解的。因此,本发明上下文中的“异源产品”是指非正常地(即非天然地或非原生地)由细菌产生的产品。因此,对于生产异源产品的细菌,其可能需要进行遗传修饰或暴露于异源底物。因此,“异源”可以表示“外源”。术语“异源”与“同源”和“内源”形成对比,“同源”和“内源”在本领域中也是普遍理解的。
55.在实施方案中,该产品是同源产品。
56.术语“同源”在本领域中是普遍理解的。因此,本发明上下文中使用的术语“同源”是指正常地(即天然地或原生地)由细菌产生的产品。因此,对于产生同源产品的细菌,其可能不需要暴露于异源底物。因此,“同源”可以表示“内源”。
57.如果该产品是同源或内源产品,在复制起点被不可逆地去除前(即转换前),细菌细胞可能已经生产了该产品。在这种情况下,不可逆地去除复制起点可用于增加产品水平或过度表达产品,例如用于增加细菌细胞在生产同源或内源产品时的效价、产量和/或生产率,例如与对照细菌细胞相比,如本文别处所讨论的。
58.在实施方案中,不可逆地(或永久地)使复制起点失活可通过遗传(或核苷酸)修饰来实现,其中遗传(或核苷酸)修饰包括或由复制起点的全部或完整核苷酸序列的去除或修饰或突变组成。换句话说,不可逆地(或永久地)使复制起点失活可通过去除复制起点的全部核苷酸序列来实现。换句话说,不可逆地(或永久地)使复制起点失活可通过去除(或缺失或消除或切除)复制起点的全部序列,即通过去除(或完全缺失或消除或切除)所有的复制起点来实现。在大肠杆菌中,ori序列被称为oric,其在普通实验室菌株中位于mnmg和mioc基因之间(messer,2002)。因此,举例来说,去除(或缺失或消除或切除)这两个基因之间的全部或部分核苷酸序列可导致复制起点的去除以及不可逆失活。
59.在另一实施方案中,不可逆地(或永久地)使复制起点失活可通过遗传(或核苷酸)修饰来实现,其中遗传(或核苷酸)修饰包括或由至少部分复制起点或仅一部分复制起点的去除或修饰或突变组成。换句话说,不可逆地(或永久地)使复制起点失活可通过去除(或缺失或消除或切除)至少部分复制起点或仅一部分复制起点来实现。
60.至少部分或仅一部分复制起点被修饰或去除(例如部分去除等)的实施方案包括导致复制起点作为复制起点发挥作用的能力丧失或显著降低的任何去除或修饰,例如在该位点起始dna复制的能力丧失或显著降低,或导致复制起点不可逆失活的任何去除或修饰。
61.本文所用的术语“去除”(或缺失或消除或切除)或类似术语表示复制起点(或其一部分)已从包含其的多核苷酸中去除(或缺失或消除或切除),从而使细菌细胞不能(或能力显著降低)复制包含其的多核苷酸。因此,如果复制起点(或其一部分)从最初包含其的多核苷酸(例如从细菌染色体)中去除,则可被视为“去除”(或缺失或消除或切除)。因此,复制起点(或其一部分)已从包含或最初包含其的多核苷酸中缺失,但仍保留在细菌细胞中(例如作为细胞内的线性序列或环状序列或质粒或附加体),则仍可被视为“去除”。任选地,术语“去除”是指从细菌细胞中去除,使其不再包含在细菌细胞中。
62.去除(或缺失或消除或切除)或修饰至少部分复制起点或仅一部分复制起点(例如部分去除)可包括去除复制起点内的多个非连续(或间断或非连贯)序列,例如修饰(例如突变)或去除一个或多个、或两个或更多等来自复制起点的dnaa盒序列,例如如本文别处所述。或者,提及复制起点的“部分”去除,或去除“部分”复制起点,可能仅包括复制起点中单个连续(或不间断或连贯)序列的修饰(例如突变)或去除。
63.在实施方案中,遗传修饰包括或由部分去除复制起点的核苷酸序列组成。
64.在实施方案中,复制起点的部分去除(或部分缺失或部分消除或部分切除)包括或由去除(或缺失或消除或切除)复制起点的至少5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、或95%、或100%的dnaa盒的核苷酸序列组成。优选地,复制起点的部分去除(或部分缺失或部分消除或部分切除)包括或由去除(或缺失或消除或切除)复制起点的至少三分之一(或至少33%)、至少一半(或至少50%)、至少三分之二(或至少67%)或全部dnaa盒的核苷酸序列组成。还提供了“高达”这些百分比的值。
65.在实施方案中,复制起点的部分去除(或部分缺失或部分消除或部分切除)包括或由去除(或缺失或消除或切除)复制起点的至少5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、或95%、或100%的dna解旋元件(due)的核苷酸序列组成。优选地,复制起点的部分去除(或部分删除或部分消除或部分切除)包括去除(或删除或消除或切除)复制起点的至少三分之一(或至少33%)、至少一半(或至少50%)、至少三分之二(或至少67%)或全部dna解链元件的核苷酸序列。还提供了“高达”这些百分比的值。
66.在实施方案中,复制起点的部分去除(或部分缺失或部分消除或部分切除)优选包括缺失复制起点的至少三分之一(或至少33%)、至少一半(或至少50%)、至少三分之二(或至少67%)或全部核苷酸序列。优选地,复制起点的部分去除(或部分缺失或部分消除或部分切除)包括去除(或缺失或消除或切除)复制起点的至少5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、或95%、或100%的核苷酸序列。还提供了“高达”这些百分比的值。
67.在实施方案中,本发明的方法包括:
68.(i)提供能够生产或生产所述产品的细菌细胞;
69.(ii)不可逆地(或永久地)使细菌细胞中的复制起点失活;以及任选地,
70.(iii)诱导细菌细胞中的产品生产。
71.用于进行步骤(ii)的适当的技术对于本领域技术人员来说是众所周知的,并且可以使用其中任何一种技术。优选技术,例如涉及使用重组或位点特异性重组的技术在本文
别处描述。
72.在实施方案中,本发明的方法的步骤(ii)和(iii)同时或依次进行。如果依次进行,步骤(ii)和(iii)可以按任何适当的顺序进行。
73.在实施方案中,细菌细胞存在于细菌细胞群体中。实际上,在本发明的所有实施方案中,细菌细胞通常是细菌细胞群体的一部分,或包括或由细菌细胞群体组成。
74.在本发明的优选方法中,本发明的方法的步骤(ii)和/或(iii)是在细菌细胞群体的指数生长期进行的。可以很容易地确定在指数生长期期间进行步骤(ii)和/或(iii)的适当的时间点,例如用于所需或最大的产品生产。例如,在一些实施方案中,在细胞密度较低(或相对较低)的指数生长期的初期(或前半部分),或者,在一些实施方案中,在细胞密度较高(例如高于或显著高于指数生长期的初期)的指数生长期的后期(例如后半部分)进行步骤(ii),即激活转换,可能是有益的。在许多实施方案中,转换的最佳时机很大程度上取决于所需达到的最终(或最大)细胞密度。如果所需的最终细胞密度较高,则很可能需要在指数生长期的后期诱导转换,而如果所需的最终细胞密度较低(或不需要那么高),则可以在指数生长期的早期诱导转换。
75.本领域中众所周知的是,细菌在培养物中的生长通常可以用四个不同的阶段来建模:(1)迟滞期,(2)指数期,(3)稳定期,以及(4)衰亡期。在迟滞期,细菌适应培养条件,细胞分裂极少或没有发生。在指数期,细胞分裂发生,因此细胞数量随每个连续的时间段而翻倍。由于生长限制因素,例如必需营养素的耗尽,达到了稳定期。在稳定期,细胞生长速率与细胞死亡速率相匹配(例如两者可以实际上都不存在),即细胞数量大致保持不变。在衰亡期,更多细菌死亡,因此(活)细胞数量减少。这些典型阶段可见于例如本文的图3、4和5。
76.在本领域中应当理解,指数生长期开始的时间可以事先预测/知道(例如通过了解相关菌株在特定条件/接种水平下的生长模式)或通过定期监测生长,例如通过测量细菌细胞培养物在600nm处的光密度(od
600
)。
77.本文所用的术语“转换”是指细菌细胞中复制起点的不可逆(或永久)失活的过程或步骤。
78.本文所用的术语“转换”可以指细菌细胞从生长阶段转变为生产阶段的过程,例如在两阶段生物过程方法中。换句话说,本文所用的术语“转换”可以指细菌细胞从以下状态转变的过程:(i)生长(例如细胞生长、增殖和/或增加生物量)优于产品生产,优选没有产品生产的状态(例如代谢状态);(ii)产品生产优于生长(例如细胞生长、增殖和/或增加生物量),优选没有生长(例如细胞生长、增殖和/或增加生物量)的状态(例如代谢状态)。
79.在本发明的方法中,细菌细胞从生长阶段转变为生产阶段的过程可由细菌细胞的复制起点的不可逆(或永久)失活诱导(或可由其引起,或可与之伴随)。令人惊讶的是,这种失活可以提供一种停止或显著减少细菌细胞生长的便捷方式。此外,这种失活可以提供一种便捷方式以将细菌细胞转换为其中产品生产优于生长(例如细胞生长、增殖和/或增加生物量)的状态,优选其中没有生长(例如细胞生长、增殖和/或增加生物量)的状态。因此,在本发明的方法中,这种不可逆地使复制起点失活的步骤优选伴随产品生产(所需产品生产),例如内源产品生产或在适当的时间点诱导产品(所需产品)生产的步骤。
80.类似地,“转换子”或“转换细胞”或“转换菌株”是指一种包含复制起点的细菌细胞,其中该细菌细胞已被修饰,使得复制起点可以不可逆地(或永久地)失活。本文所用的术
语“已转换”或“已转换细胞”或菌株是指其复制起点已不可逆地(或永久地)失活的细胞。
81.可以在任何适当的时间点诱导或进行转换或复制起点的不可逆(或永久)失活。
82.在实施方案中,在细菌细胞群体的指数生长期期间诱导或进行转换或复制起点的不可逆(或永久)失活。例如,可以在细菌细胞群体的指数生长期开始时、或大约开始时、或开始后立即或开始后不久诱导或进行复制起点的不可逆(或永久)失活。换句话说,例如可以在指数生长期的前半部分,或在细胞密度达到最大细胞密度(或od)的50%时或之前的时间点诱导或进行复制起点的失活。在其他实施方案中,在细胞密度较高的细菌细胞群体的指数生长期期间,例如在指数生长期的后半部分,或在细胞密度达到最大细胞密度(或od)的50%之后的时间点诱导或进行转换或复制起点的不可逆(或永久)失活。
83.在另一个实施方案中,可以在细菌细胞群体的指数生长期之前诱导或进行转换或复制起点的不可逆(或永久)失活。例如,可以在细菌细胞群体的指数生长期开始前立即(或不久)诱导或进行复制起点的不可逆(或永久)失活。
84.因此,在将细菌接种到培养基中后,或例如在细菌培养物的细胞密度(光密度)达到一定或所需的阈值水平后立即或不久后,在预测的或方便的或合适的时间诱导或进行转换或复制起点的不可逆(或永久)失活。
85.优选地,在其后例如紧随其后应起始或诱导转换(即复制起点的不可逆失活)的阈值光密度水平为转换后的细菌细胞培养物或对照细菌细胞培养物的峰值细胞密度(或最大细胞密度,或最高细胞密度,或平台期细胞密度),例如在没有转换的情况下细菌细胞培养物的峰值细胞密度的至少1%、至少2%、至少3%、至少4%、至少5%、至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、或至少90%。
86.优选地,紧随其后应起始或诱导转换(即复制起点的不可逆失活)的阈值光密度水平为转换后的细菌细胞培养物或对照细菌细胞培养物的峰值细胞密度(或最大细胞密度,或最高细胞密度,或平台期细胞密度),例如在没有转换的情况下细菌细胞培养物的峰值细胞密度的1至5%、5至10%、10至15%、15至20%、20至25%、25至30%、30至35%、35至40%、40至45%、45至50%、50至55%、55至60%、60至65%、65至70%、70至75%、75至80%、80至85%、或85至90%。
87.峰值或最大值的光密度水平等,如上文和本文别处所讨论的细胞密度可以预先确定,但对于不同的细菌细胞在不同的条件下,例如不同的生长条件,如培养基和温度,或生产不同的产品,会有所不同。因此,使用在选定或所需条件下达到的最大细胞密度的百分比,特别便于确定应何时起始或诱导转换(或复制起点的不可逆失活)。
88.对照细菌细胞培养物可以例如是本发明的细菌细胞培养物,其中未诱导复制起点的不可逆失活(即复制起点的不可逆失活没有发生或不能发生)。
89.所需产品的生产可以在本领域技术人员可以很容易地确定的任何适当的时间点进行。
90.在实施方案中,可以在诱导或进行转换或复制起点的不可逆(或永久)失活的同时(或大约在转换或复制起点失活同时,或在转换或复制起点的失活之前立即,或在转换或复制起点的失活之后立即,或在转换或复制起点的失活之前或之后不久)起始或诱导所需产品的生产(例如通过诱导负责产生所需产品的酶或代谢途径的生产和/或激活或诱导产品
的生产,例如当产品是蛋白质时,通过诱导编码产品的基因或核酸序列的表达)。
91.或者,起始或诱导产品(所需产品)生产的时间不一定与诱导转换或复制起点的失活的时间相关,且可以在任何适当的时间点进行,只要细菌细胞能够生产可测量的或显著水平的产品(所需产品)。例如,在实施方案中,在细菌细胞群体的指数生长期期间(或大约开始时,或开始前立即,或开始后立即,或开始前不久,或开始后不久)起始或诱导产品生产(例如通过诱导负责产生所需产品的酶或代谢途径的生产和/或激活或诱导产品的生产,例如当产品是蛋白质时,通过诱导编码产品的基因或核酸序列的表达)。换句话说,例如可以在指数生长期的前半部分,或在细胞密度达到最大细胞密度(或od)的50%时或之前的时间点诱导或进行产品生产。在其他实施方案中,可以例如在细胞密度较高的细菌细胞群体的指数生长期期间,例如在指数生长期的后半部分,或在细胞密度达到最大细胞密度(或od)的50%之后的时间点诱导或进行产品生产。
92.或者,产品的生产可能不需要与指数期的时间相关。例如,该产品可以是组成型或内源产生的(在这种情况下不一定需要起始或诱导步骤),或者起始或诱导可以在细菌细胞培养物生长的指数期开始之前或之后开始。
93.在备选实施方案中,在指数生长期期间或甚至在稳定期期间的其他时间点起始或诱导产品的生产。实际上,本发明的方法的一个令人惊讶的优点是,当细菌细胞处于生长稳定期时,其仍可被起始或诱导以生产产品(所需产品)。
94.因此,可以在将细菌接种到培养基中后,或例如在细菌培养物的光密度达到一定或所需的阈值水平后立即或不久后,在预测的或方便的或合适的时间诱导或进行产品的生产。
95.优选地,紧随其后应启动或诱导产品生产的阈值光密度水平为转换后的细菌细胞培养物或对照细菌细胞培养物的峰值细胞密度(或最大细胞密度,或最高细胞密度,或平台期细胞密度),例如在没有转换的情况下细菌细胞培养物的峰值细胞密度的至少1%、至少2%、至少3%、至少4%、至少5%、至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、或至少90%。
96.优选地,紧随其后应启动或诱导产品生产的阈值光密度水平为转换后的细菌细胞培养物或对照细菌细胞培养物的峰值细胞密度(或最大细胞密度,或最高细胞密度,或平台期细胞密度),例如在没有转换的情况下细菌细胞培养物的峰值细胞密度的1至5%、5至10%、10至15%、15至20%、20至25%、25至30%、30至35%、35至40%、40至45%、45至50%、50至55%、55至60%、60至65%、65至70%、70至75%、75至80%、80至85%、或85至90%。
97.峰值或最大值的光密度水平等,如上文和本文别处所讨论的细胞密度可以预先确定,但对于不同的细菌细胞在不同的条件下,例如不同的生长条件,如培养基和温度,或生产不同的产品,会有所不同。因此,使用在选定或所需条件下达到的最大细胞密度的百分比,特别便于确定应何时启动或诱导产品生产。
98.对照细菌细胞培养物可以例如是本发明的细菌细胞培养物,其中未诱导复制起点的不可逆失活(即复制起点的不可逆失活没有发生或不能发生)。
99.可以理解的是,在一些实施方案中,用户没有必要诱导产品的生产,该步骤可以是任选的,例如因为产品的诱导或生产于特定时间点在培养物中自动发生。例如,当产品的生
产与对内部刺激或内部条件敏感的启动子相关联时,就可能出现这种情况。例如,可以在细菌细胞培养物进入特定生长期时诱导启动子,例如pcp_2836启动子(ma et al.,2018)、phopr启动子(paul et al.,2004)和p170启动子(madsen et al.,1999)的情况。优选地,可以在细菌细胞培养物进入指数生长期时或在指数生长期期间诱导启动子。其他优选设置包括使用自动诱导培养基(例如如studier et al.2005,protein expression and purification,2005,41:207-234中所述)和自诱导系统(例如silex,如briand et al.2016,scientific reports,6:33037中所述)。
100.或者,如果产品是组成型或连续或恒定地生产的,例如在组成型或内源启动子的控制下,例如内源产品(尽管同样的组成型或连续或恒定的生产可应用于异源产品的生产),则用户可能没有必要在细菌细胞中诱导产品的生产。在这种情况下,细菌细胞中复制起点的不可逆失活可以使所需产品的生产或水平增加或提高,例如以增加细菌细胞生产所述产品的效价、产量和/或生产率。
101.在实施方案中,细菌细胞包含编码产品或编码生产产品所需的酶的基因(或核酸分子)。此类基因可以是同源的(内源的)或异源的。
102.在实施方案中,该基因可操作地与启动子连接。此类启动子可以控制产品或酶的表达,例如同源(内源)或异源产品或酶。因此,在一些实施方案中,此类启动子可以是内源启动子(例如基因或细菌细胞原生的启动子),也可以是组成型启动子,以能够连续或恒定表达。或者,此类启动子可以是诱导型或异源启动子,例如可用于能够于所需时间点在细菌细胞中诱导由该基因编码的产品的生产。
103.适当的诱导型启动子在本领域是众所周知的且有所描述,并且可以使用这些启动子中的任何一个。例如,在实施方案中,启动子可以是ph敏感的、光敏感的、温度敏感的或化学敏感的。此类启动子和系统的示例和更多细节将在本文别处讨论。示例性系统也在示例中示出。
104.在其他实施方案中,基因或启动子的表达可以通过靶向蛋白水解或其他适当的技术如crispr干扰(crispri)诱导或控制。
105.crispr干扰(crispri)的概念在本领域是众所周知的,例如在lei et al.2013(cell152,1173-1183)中有所描述。在crispri中,使用具有特定序列的sgrna,将催化失活版本的cas9靶向特定dna序列。催化失活cas9与目标dna区(例如启动子区)结合,以阻断rna聚合酶结合和转录延伸。
106.因此,在其中基因或启动子的表达由crispr干扰(crispri)诱导或控制的细菌细胞的实施方案中,细菌细胞包含编码催化失活cas9的基因,以及用于将催化失活cas9靶向该基因或启动子以生产所需产品,从而抑制基因或启动子的表达的sgrna。
107.在本发明的方法的替代观点中,或在另一方面,本发明提供了一种不可逆地(或永久地)停止细菌细胞的生长而不损害细菌细胞的代谢活性的方法,该方法包括不可逆地(或永久地)使细菌细胞中的复制起点失活。因此,本发明的方法还提供了一种停止细菌细胞的生长而不损害细菌细胞的代谢活性的方法。
108.短语“停止细菌细胞的生长而不损害细菌细胞的代谢活性”是指细菌细胞不能参与dna复制,但在其他方面基本不受影响,即细菌细胞的代谢途径,特别是与所需产品生产相关的途径,如产品或酶的基因表达,保持完整或功能(或至少基本完整或功能)。这与本领
域的两阶段生物过程方法不同,其中从生长阶段转变为生产阶段需要一种或多种特定代谢途径的靶向下调或失活(例如,通过靶向抑制代谢途径酶或靶向抑制编码所述酶的基因表达)和/或其他特定途径的靶向上调或激活(例如,通过靶向激活代谢途径酶或靶向诱导编码所述酶的基因表达)。
109.在另一方面,本发明提供了一种包含复制起点的细菌细胞,其中该细菌细胞已被修饰,使得复制起点可以不可逆地(或永久地)失活。因此,这些方面提供了经修饰的细菌细胞。
110.不可逆地使遗传元件失活的各种方法在本领域是已知的,并且技术人员会理解如何将这些方法应用于复制起点的失活。例如,所述细菌细胞可包含经遗传修饰的染色体,使得该染色体中的复制起点不能充当复制起点,例如不能充当dna复制的起始位点。可以通过复制起点的突变或缺失来执行失活。优选地,可以例如通过控制负责或参与突变或缺失事件的试剂来诱导或控制失活。例如,可以通过启动子诱导或控制失活,该启动子可操作地与编码负责或参与突变或缺失事件的试剂的基因连接。然后可以通过诱导所述启动子来触发复制起点的不可逆失活。因此,对此优选的启动子是诱导型启动子。
111.根据本发明,不可逆地使复制起点失活的优选方法是通过使用同源重组来去除或缺失全部或部分复制起点的核苷酸序列。例如,可以对含有复制起点的染色体进行遗传修饰,从而包括适当的位点以起始同源重组,例如通过位点特异性重组酶识别位点特异性重组位点,其定位和定向从而允许去除或缺失全部或部分复制起点的核苷酸序列。然后可以向细菌细胞提供位点特异性重组酶,方便地在细菌细胞内表达它们。这种表达理想地需要被控制,优选被严格控制,以便当需要触发同源重组(或转换或起点失活)时,可以在所需或适当的时间点诱导重组酶的表达。这可以方便地通过将重组酶的表达置于诱导型启动子的控制下来完成。
112.在包含不可逆失活的复制起点的细菌细胞或使用所述细菌细胞的方法的实施方案中,可以使用本领域的标准技术去除(或可去除)复制起点,例如使用重组酶,例如使用位点特异性重组或同源重组,例如使用丝氨酸重组酶或酪氨酸重组酶系统,或λ噬菌体red重组酶系统(参见例如progress in biophysics and molecular biology 147(2019)33e46)。
113.在包含不可逆失活的复制起点的细菌细胞或使用所述细菌细胞的方法的实施方案中,可通过用另一序列替换复制起点,例如通过同源重组来去除复制起点(即可发生转换)。在此类实施方案中,替换序列应该是在转换后不能使dna复制起始保留在该位点的序列。换句话说,替换序列不应该是能使dna复制起始从该位点保留的序列。例如,不可逆失活的复制起点不应被在所述细菌细胞中具有功能的另一复制起点替换。
114.在包含不可逆失活的复制起点的细菌细胞或使用所述细菌细胞的方法的实施方案中,可通过用另一序列替换复制起点,例如通过同源重组来去除复制起点(即可发生转换)。在此类实施方案中,替换序列应该是在转换后不能使dna复制起始保留在该位点的序列。换句话说,替换序列不应该是能使dna复制起始从该位点保留的序列。例如,不可逆失活的复制起点不应被在所述细菌细胞中具有功能的另一复制起点替换。
115.同样可以使用其他类型的基因组编辑来不可逆地(或永久地)使复制起点失活。可能的候选方案包括crispr/cas系统(单独使用或与另一重组酶系统如λ噬菌体red重组酶系
统结合使用)或使用任何其他类型的不可逆地(或永久地)改变的酶,例如去除或切除细菌中(例如细菌染色体中)的复制起点序列,并使所述复制起点不能起始dna复制,如本文别处所讨论的。
116.因此,示例性经修饰的细菌细胞可能包含旁侧有重组位点的复制起点,例如位点特异性重组位点,以使全部或部分复制起点序列不可逆地(或永久地)失活,例如通过允许去除或切除复制起点。使用本领域的标准技术可以很容易地引入此类重组位点。
117.包含复制起点的细菌细胞,其中该细菌细胞已被修饰使得复制起点可以不可逆地(或永久地)失活,其可以使用本领域的标准技术生成,例如使用重组酶,例如使用同源重组,例如使用λ噬菌体red重组酶系统(参见例如progress in biophysics and molecular biology 147(2019)33e46)。这种使用重组酶的同源重组可以例如使用载体进行,该载体包含旁侧有位点特异性重组位点的复制起点,其中位点特异性重组位点本身旁侧有(即嵌套在)与目标细菌细胞的染色体中的相应序列同源的核苷酸序列。
118.在生成包含不可逆失活的复制起点的细菌细胞的过程中,起始细菌细胞的原始(即原生、天然、野生型或内源)复制起点可以用不同的复制起点替换。例如,插入的(即不可逆失活)复制起点与起始细菌细胞的原始(即原生、天然、野生型或内源)复制起点相比,可具有不同的序列(例如复制起点可能来自不同的细菌种类或菌株,或可能是外源序列)。
119.类似地,在本文所述的本发明的生产产品的方法和其他方面,在细菌细胞中不可逆地失活(即不可逆失活)的复制起点与所使用的细菌细胞的原始(即原生、天然、野生型或内源)复制起点相比,可具有不同的序列(例如复制起点可能来自不同的细菌种类或菌株,或可能是外源序列)。
120.在实施方案中,复制起点旁侧有位点特异性重组位点(例如位点特异性识别和切割位点),用于通过位点特异性重组酶进行识别和切割。
121.然后相关的位点特异性重组酶可以结合并切割所述位点,以促进或使dna重组成为可能。
122.在实施方案中,复制起点旁侧有位点特异性重组位点,这些位点特异性重组位点可被位点特异性重组酶识别(例如可能存在两个位点特异性重组位点,一个位于复制起点的5’处,一个位于复制起点的3’处)。
123.在实施方案中,位点特异性重组位点具有不同的序列,例如attb和attp位点。或者,位点特异性重组位点具有相同的序列(即位点特异性重组位点是相同的),例如loxp位点。适当的位点特异性重组位点的选择取决于所使用的重组酶,并且对于本领域技术人员来说可以很容易地推导得出。
124.在实施方案中,每个位点特异性重组位点置于离复制起点的一端(5’端或3’端)适当的距离处,使其旁侧于期望去除的核苷酸区,包括全部或部分复制起点,并且间隔适当,以便可以成功发生同源重组。
125.大多数细菌和细菌细胞仅含有单一复制起点(例如单一染色体复制起点),尽管在某些情况下复制起点包含一段以上的核苷酸,例如起点不存在于单个连续的核苷酸序列中。
126.例如,在枯草芽孢杆菌中,复制起点是二部分的,即复制起点由两个核苷酸段组成,被不属于复制起点的内含子(dnaa基因)所分隔。因此,在枯草芽孢杆菌和其他具有二部
分复制起点的细菌菌株的情况下,短语“复制起点旁侧有位点特异性重组位点”或类似的短语可以表示重组位点旁侧于复制起点的一个或两个核苷酸段。
127.细菌细胞具有多个复制起点(例如多个染色体复制起点)的情况下,可通过不可逆地(或永久地)使细菌细胞中的一个、至少一个或全部复制起点(例如染色体复制起点)失活来诱导根据本发明的产品生产的增加(优选根据效价、产量或生产率)。因此,在实施方案中,如果复制起点是多部分的,即由两个单独的核苷酸段(即二部分)或更多个(例如三部分)组成,复制起点可通过去除(或缺失或消除或切除)复制起点的一个或两个或所有核苷酸段而不可逆地(或永久地)失活。
128.细菌细胞可在染色体外遗传物质如质粒或附加体上包含一个或多个复制起点。在这种情况下,仅不可逆地(或永久地)使染色体复制起点失活便足以促进转换。因此,在实施方案中,转换包括或由不可逆地(或永久地)使细菌细胞中的染色体复制起点失活组成(或由其诱导)。优选地,转换包括或由仅使细胞中的一个复制起点失活,优选仅使细胞中的一个染色体复制起点失活组成(或由其诱导)。
129.本文所用的术语“重组酶”包括能够通过切除、插入、倒位或易位进行dna操作的任何位点特异性酶。因此,术语“重组酶”不仅包括重组酶,还包括整合酶、转化酶和解离酶。
130.本文所用的术语“位点特异性重组酶”(或位点特异性整合酶或位点特异性转化酶或位点特异性解离酶)是指催化该重组酶特异性的靶位点序列之间的dna交换反应的酶。因此,本文所用的位点特异性重组酶(和位点特异性重组酶位点)可以促进去除(或缺失或消除或切除)至少一部分,优选全部细菌细胞的复制起点。
131.位点特异性重组位点的相对方向,例如一对位点特异性重组位点,决定了重组事件的结果。本文所用的术语位点特异性重组位点的“方向”(或方位或方向性)在本领域中是普遍理解的,并与多核苷酸中提供的位点特异性重组酶位点的方向有关。例如,当同一多核苷酸上的位点特异性重组位点,例如一对位点特异性重组位点具有相反的方向时,多核苷酸内的侧翼序列可能发生倒位(例如一对loxp位点(在cre-lox系统中),或由attp位点和attb位点(例如在丝氨酸重组酶系统中发现)组成的一对)。相反,如果两个位点特异性重组位点具有相同的方向(或平行方向),则可能发生侧翼序列从多核苷酸上切除。
132.因此,在实施方案中,位点特异性重组位点旁侧于细菌染色体中的复制起点(即位于其任一侧),其方向使得位点特异性重组导致复制起点从细菌染色体上切除。在实施方案中,位点特异性重组位点可以以相同的方向定位在多核苷酸上。或者,位点特异性重组位点可以以相反的方向定位在多核苷酸上(即其中一个位点处于“有义”方向,另一个位点处于“反义”方向)。
133.在实施方案中,位点特异性重组位点包含用于丝氨酸重组酶识别的位点特异性重组位点。优选地,所述丝氨酸重组酶为γδ、bxb1、或tp901。然而,本领域和实验中描述了许多不同的丝氨酸重组酶,生物信息学预测了超过4000种新的丝氨酸重组酶(yang et al.,2014)。可以使用其中任何一种。
134.在实施方案中,位点特异性重组位点包含用于丝氨酸整合酶识别的位点特异性重组位点。
135.丝氨酸位点特异性整合酶(丝氨酸位点特异性重组酶的亚家族)的重组机制在本领域中是普遍理解的。丝氨酸位点特异性整合酶催化attp和attb位点的重组,生成attl和
attr位点。attp和attb位点的重组结果取决于位点特异性重组位点的方向。方向相同并位于同一多核苷酸上的attp位点和attb位点的重组导致从多核苷酸上切除侧翼序列,在剩余的多核苷酸中留下attr或attl位点。与之相对,方向相反的attp位点和attb位点的重组导致dna序列倒位。
136.因此,在实施方案中,丝氨酸位点特异性重组位点旁侧于细菌染色体中的复制起点(即位于其任一侧),其方向使得重组(当由适当的酶催化时)导致复制起点从细菌染色体上切除。优选地,丝氨酸位点特异性重组位点可以以相同的方向定位在多核苷酸上。或者,丝氨酸位点特异性重组位点可以以相反的方向定位在多核苷酸上(即其中一个位点处于“有义”方向,另一个位点处于“反义”方向)。
137.在实施方案中,位点特异性重组位点包含用于酪氨酸重组酶识别的位点特异性重组位点,优选地,其中所述酪氨酸重组酶为cre、dre、flp、kd、b2或b3。
138.适用于本发明的位点特异性重组酶技术的示例包括丝氨酸位点特异性重组、酪氨酸位点特异性重组、cre-lox重组、flp-frt重组和同源重组。
139.在实施方案中,位点特异性重组位点包含一对位点特异性重组位点或由一对位点特异性重组位点组成。
140.本文所用的术语“一对”位点是指被同一位点特异性重组酶识别的两个位点,并且其中两个位点都必须存在以使重组事件发生。因此,一对位点特异性重组位点的示例是一个attp位点和一个attb位点(例如用于丝氨酸重组酶/整合酶的识别),或两个loxp位点(例如用于cre重组酶的识别)。
141.成对的位点特异性重组位点的其他示例在本领域是已知的。因此,成对的位点特异性重组位点的其他示例是:两个frt位点(例如用于flp重组酶的识别);两个rox位点(例如用于dre重组酶的识别);两个kd重组酶靶(kdrt)位点(例如用于kd重组酶的识别);两个b2重组酶靶(b2rt)位点(用于b2重组酶的识别);或两个b3重组酶靶(b3rt)位点(用于b3重组酶的识别)。
142.因此,本文所述的“一对”位点可以是两个不同的位点(具有两个不同的序列)或两个相同的位点(具有相同的序列),这取决于所使用的重组酶。
143.包含不可逆(或永久)可去除的复制起点的经修饰的细菌细胞有可能发生变化,无意中使复制起点变得不可去除(即不再可去除)。例如,一个或多个位点特异性重组位点的突变,或编码对所述位点具有特异性的位点特异性重组酶的一个或多个基因的突变,可导致复制起点可去除性的丧失。
144.因此,提供一种复制起点旁侧有两对、三对或四对(或至少或最多两对、三对或四对)位点特异性重组位点的细菌细胞可能是有益的,优选两对位点特异性重组位点。在不希望受理论约束的情况下,与其中复制起点仅旁侧有一对位点特异性重组位点的细菌细胞相比,此类细菌细胞失去不可逆地使其复制起点失活的能力的概率降低。
145.因此,在实施方案中,位点特异性重组位点包含或由两对或更多对,优选两对位点特异性重组位点组成。
146.换句话说,位点特异性重组位点包含或由三对、四对、五对或六对位点特异性重组位点组成。
147.随着成对的位点特异性重组位点数量的增加,本发明的细菌细胞失去不可逆地使
其复制起点失活的能力的概率降低。因此,例如,具有三对位点特异性重组位点的细菌细胞失去不可逆地使其复制起点失活的能力的概率低于具有两对位点特异性重组位点的细菌细胞失去不可逆地使其复制起点失活的能力的概率,以此类推。
148.在两对或更多对位点特异性重组位点旁侧于复制起点的情况下,可沿染色体以任何顺序提供成对的位点特异性重组位点,只要该顺序能使重组成功发生。
149.在实施方案中,存在两对位点特异性重组位点的情况下,其中一对位点特异性重组位点嵌套在另一对重组位点内,即一对重组位点旁侧有另一对重组位点。因此,对于两对重组位点a和b,这些位点沿多核苷酸以a-b-b-a的顺序排列,或以b-a-a-b的顺序排列。
150.或者,每对位点特异性重组位点沿多核苷酸交替提供,使得任何一对都不嵌套在另一对内;换句话说,对于两对重组位点a和b,这些位点在多核苷酸中以a-b-a-b的顺序排列,或以b-a-b-a的顺序排列。
151.两对或更多对重组位点的各个组分在空间上以能够发生重组的方式排列。因此,在一个优选的实施方案中,适当数量的核苷酸将相邻的a和b位点(即每对的相邻位点)分开。在另一个优选的实施方案中,没有核苷酸将相邻的a和b位点(即每对的相邻位点)分开;换句话说,相邻的a和b位点(即每对的相邻位点)形成一个连续序列。
152.上述段落中提供的实施方案加以必要的修正适用于更多组成对的位点特异性重组位点,例如三个位点特异性重组位点。
153.在实施方案中,本发明的细菌细胞进一步包含编码位点特异性重组酶的基因。
154.在实施方案中,编码位点特异性重组酶的基因可操作地与启动子连接。
155.适当地选择位点特异性重组酶,使其可以与选定的位点特异性重组位点一起发挥作用。因此,在使用两对或更多对重组位点的实施方案中,细菌细胞可进一步包含编码每对重组位点的位点特异性重组酶的基因(视情况而定),例如编码两种或更多种重组酶,例如3、4、5或6种重组酶的基因(视情况而定)。此外,编码每个位点特异性重组酶的基因可以可操作地与相同或不同的启动子连接。此外,编码每个位点特异性重组酶的基因可以在相同或不同的核酸序列上,例如在相同或不同的染色体外元件上,例如质粒或构建体。
156.本文所用的术语“启动子”具有其本领域公认的含义,并且通常被理解为指与蛋白结合以控制、起始和/或抑制启动子下游dna序列的转录的dna序列。当指出编码重组酶的基因与启动子“可操作地连接”时,这意味着启动子相对于基因处于正确的功能位置和/或方向,以通过启动子的操作控制与基因连接的序列的转录。
157.在实施方案中,启动子对内部刺激或内部条件敏感。例如,可以在细菌细胞培养物进入特定生长期时诱导启动子,如pcp_2836启动子(ma et al.,2018)、phopr启动子(paul et al.,2004)和p170启动子(madsen et al.,1999)的情况。优选地,可以在细菌细胞培养物进入指数生长期时或在指数生长期期间诱导启动子。其他优选设置包括使用自动诱导培养基(例如如studier et al.2005中所述,同上)和自诱导系统(例如silex,如briand et al.2016,scientific reports,6:33037中所述)。
158.在工业环境中对内部条件敏感的启动子有时是优选的,因为这些启动子不需要用户的干预(例如施用化学诱导剂)来诱导,并且在本发明中由此诱导转换或复制起点的不可逆失活。
159.在实施方案中,启动子对一种或多种外部刺激敏感,例如是诱导型启动子。因此,
可以根据外部刺激或用户希望起始转换(或复制起点的失活)的刺激来选择所使用的启动子。
160.在一些此类实施方案中,启动子是温度敏感的、ph敏感的、光敏感的或化学敏感的。具有这些和其他特性(例如其他可诱导的特性)的启动子在本领域是众所周知的且有所描述,并且可以很容易地选择用于本发明的适当的启动子。
161.例如,可用于相同目的的其他类型的基因表达控制方法包括但不限于诱导剂(iptg、阿拉伯糖、高丝氨酸内酯、无水四环素等)诱导(marschall,sagmeister and herwig,2017)(lutz and bujard,1997)(cox,surette and elowitz,2007)、靶向蛋白水解(cameron and collins,2014)、crispr/cas9的不同变体(qi et al.,2013)等。本领域技术人员会知道如何使用此类其他类型的基因表达控制方法。例如,在化学诱导启动子的情况下,适当的化学品被添加到生长培养基中,通过扩散或促进运输进入细胞。在细胞内,该化学品与其目标转录调控因子结合,这导致来自所述转录调控因子的目标启动子的转录激活。如果转录阻遏因子控制来自启动子的表达并且使所述转录阻遏因子对蛋白水解敏感,则可以通过诱导所述转录调控因子的蛋白水解来诱导来自启动子的表达。可以通过在细胞中表达适当的向导rna将催化失活crispr/cas9靶向启动子,并且根据确切的配置,这可以导致所述启动子的激活或阻遏。
162.因此,在实施方案中,启动子是化学敏感的,并且合适的启动子在本领域中会很容易获得。例如,优选的启动子是那些其中启动子可使用异丙基β-d-1-硫代半乳糖苷(iptg)、阿拉伯糖、高丝氨酸内酯或无水四环素诱导的启动子,例如其中启动子是来自lac操纵子、ara操纵子、tet操纵子或lux操纵子的启动子,例如其中启动子是lac启动子(lac p)、arabad启动子、ppui启动子、tet启动子(p
tet
启动子)或luxi启动子。
163.例如,细菌细胞可包含编码位点特异性重组酶的基因,并且所述基因的表达可在lac操纵子表达系统下控制。所述基因的表达可由iptg或异乳糖诱导。例如,可以在质粒上提供该基因,其中在该基因的上游提供lac启动子和lac操纵子,并且该质粒进一步包含lac阻遏因子(lac i)。适合在该系统下控制靶基因的质粒在本领域是已知的。
164.在另一个示例中,细菌细胞可包含编码位点特异性重组酶的基因,并且所述基因的表达可在ara操纵子表达系统下控制。所述基因的表达可由阿拉伯糖诱导。例如,细菌细胞包含质粒上提供的基因,其中arabad启动子位于该基因的上游,并且该质粒还包含使用ara操纵子进行基因调控所必需的其他遗传元件。基因调控所必需的ara操纵子的遗传元件是本领域技术人员已知的并在上文提供。
165.在另一个示例中,细菌细胞可包含编码位点特异性重组酶的基因,并且所述基因的表达可在lux操纵子表达系统下控制。所述基因的表达可由ahl诱导。例如,细菌细胞包含质粒上提供的基因,其中luxi启动子位于该基因的上游,并且该质粒还包含luxr基因,优选地,其中luxr基因为组成型表达。
166.在另一个示例中,细菌细胞可包含编码位点特异性重组酶的基因,并且所述基因的表达可在tet-off或tet-on表达系统下控制。所述基因的表达可由四环素(或无水四环素或多西环素)诱导。例如,细菌细胞包含(i)质粒上提供的基因,其中四环素(或无水四环素或多西环素)依赖型启动子位于该基因的上游,和(ii)tta表达质粒或rtta表达质粒。适合在该系统下控制靶基因的质粒在本领域是已知的。
167.在实施方案中,基因或启动子的表达可以通过靶向蛋白水解来诱导或控制。
168.在实施方案中,基因或启动子的表达由crispr干扰(crispri)诱导或控制。
169.在优选的实施方案中,启动子是温度敏感的。
170.可以使用任何适当的温度敏感性启动子,其中在本领域有许多示例。
171.可以选择在任何合适的温度下激活(和/或处于其最高活性水平)本发明中使用的温度敏感性启动子。优选地,该启动子可以是温度敏感性启动子,其在20至25℃、25至30℃、30至35℃、35至40℃、40至45℃、45至50℃、50至55℃、或55至60℃、或高于60℃被激活(和/或处于其最高活性水平)。优选地,该启动子可以在约37℃被激活(和/或处于其最高活性水平)。
172.因此,在实施方案中,可以通过改变(即升高或降低)细菌细胞(或细菌细胞培养物)的温度,使其达到温度敏感性启动子被激活(和/或处于其最高活性水平)的温度(或温度范围)来诱导本发明中使用的温度敏感性启动子。因此,为了诱导转换,温度可以改变为20至25℃、25至30℃、30至35℃、35至40℃、40至45℃、45至50℃、50至55℃、或55至60℃、或高于60℃。
173.使用的温度敏感性启动子可以在较高温度下被激活,在这种情况下,细菌细胞(或细菌细胞培养物)的温度最初可能需要较低(例如30℃或低于30℃),然后升高(例如升至37℃或高于37℃)以诱导激活并因此发生转换(或起点失活)。相反,使用的温度敏感性启动子可能在较低温度下被激活,在这种情况下,细菌细胞(或细菌细胞培养物)的温度最初可能需要较高(例如37℃或高于37℃),然后降低(例如降至30℃或低于30℃)以诱导转换(或起点失活)。优选地,该启动子可以在或约37℃(或高于37℃)被激活(和/或处于其最高活性水平)。因此,方便地,细菌细胞(或细菌细胞培养物)的温度最初可以较低,例如在或约30℃,例如25℃至33℃,然后升高(例如在或约37℃,例如36℃至39℃,或高于37℃)以诱导转换(或起点失活)。
174.用于本发明的优选温度敏感性启动子是可由温度敏感性试剂(或由温度敏感性阻遏因子,或由温度敏感性激活剂)控制或调节的启动子,例如λ噬菌体cl857阻遏因子。在实施方案中,温度敏感性启动子是λ噬菌体pr或p
l
启动子(或pr或p
l
启动子的修饰形式),由λ噬菌体cl857阻遏因子控制,这些启动子在或约在37℃被激活(和/或具有其最高活性水平)(jechlinger et al.,1999)。因此,使用该启动子可以方便地通过将细菌细胞(或细菌细胞培养物)的温度从30℃或约30℃(例如25℃至33℃)改变至37℃或约37℃(例如36℃至39℃,或高于37℃)以诱导转换(或起点失活)。
175.在实施方案中,功能元件,例如在复制起点的不可逆失活中使用(或必需)的一个或多个或所有功能元件(即遗传元件)位于(或整合在)基因组上,例如染色体(即细菌染色体)上。例如,在实施方案中,编码位点特异性重组酶的遗传元件(或基因)位于(或整合在)基因组上,例如染色体(即细菌染色体)上。或者,整个重组酶表达系统(或转换系统)可以位于(或整合在)基因组上。例如,编码位点特异性重组酶的遗传元件和编码重组酶调控因子组分的遗传元件可以位于(或整合在)基因组上,例如染色体上。重组酶调控因子组分可包括如本文定义的温度敏感性启动子(例如pr或p
l
启动子或其修饰形式),以及任选地在适当情况下编码促进如本文定义的敏感性的试剂的基因,例如编码温度敏感性阻遏因子的基因,例如编码ci857阻遏因子的基因。
176.在实施方案中,功能元件,例如在复制起点的不可逆失活中使用(或必需)的一个或多个或所有功能元件(即遗传元件)位于(或整合在)一个或多个附加体或质粒上,或位于细菌细胞内的一些其他适当的染色体外元件上。例如,在实施方案中,编码位点特异性重组酶的遗传元件(或基因)位于(或整合在)一个或多个附加体或质粒上,或位于细菌细胞内的一个或多个其他适当的染色体外元件上。或者,整个重组酶表达系统(或转换系统)可以位于(或整合在)一个或多个附加体或质粒(或其他适当的元件)上。例如,编码位点特异性重组酶的遗传元件(或基因)和编码重组酶调控因子组分的遗传元件(或基因)可以位于(或整合在)一个或多个附加体或质粒(或其他适当的元件)上。重组酶调控因子组分可包括如本文定义的温度敏感性启动子(例如pr或p
l
启动子或其修饰形式),以及任选地在适当情况下编码促进如本文定义的该敏感性的试剂的遗传元件(或基因),例如编码温度敏感性阻遏因子的基因,例如编码ci857阻遏因子的基因。
177.或者,编码位点特异性重组酶的遗传元件(或基因)可以位于基因组上,而编码相关重组酶调控因子组分的遗传元件可以位于一个或多个附加体或质粒上;或反之亦然。
178.在实施方案中,复制起点的不可逆失活中使用(或必需)的一个或多个功能元件(即遗传元件)是密码子优化的。例如,编码位点特异性重组酶的遗传元件(或基因)可以是密码子优化的。备选地或另外地,在使用此类温度敏感性启动子系统的情况下,编码促进温度敏感性启动子的敏感性的温度敏感性试剂(例如编码温度敏感性阻遏因子,例如编码cl857)的遗传元件(或基因)是密码子优化的。
179.在另一方面,本发明提供了一种经修饰的细菌细胞,其中所述经修饰的细菌细胞缺乏功能性复制起点序列。因此,在优选的实施方案中,此类细菌细胞是经遗传修饰的。换句话说,此类细菌细胞的基因组或染色体经修饰,使其缺乏功能性复制起点。生产此类细菌细胞的方法在本文别处与本发明的方法相关描述,其涉及不可逆地使细菌细胞中的复制起点失活的步骤。
180.因此,又一方面提供了一种经修饰的细菌细胞,其中所述经修饰的细菌细胞缺乏功能性复制起点序列,其中所述经修饰的细菌细胞通过一种方法获得、生产或可获得,该方法包括不可逆地使细菌细胞中的复制起点失活的步骤。进行这种不可逆失活的适当方法在本文别处有所描述,例如包括复制起点遗传修饰的方法,例如部分或完全去除复制起点的核苷酸序列。
181.在实施方案中,本发明提供了一种经修饰的细菌细胞,其中所述经修饰的细菌细胞不含有一个(或任何)功能性复制起点序列。优选地,经修饰的细菌细胞不具备dna复制能力(即不具备任何染色体dna复制能力)。
182.在实施方案中,本发明提供了一种经修饰的细菌细胞,其中所述经修饰的细菌细胞不含有一个(或任何)功能性染色体复制起点序列。优选地,经修饰的细菌细胞不具备染色体dna复制能力(即不具备任何染色体dna复制能力)。
183.在实施方案中,细菌细胞或经修饰的细菌细胞可以生产产品(或能够生产产品或正在生产产品),例如根据本发明的方法生产所需产品。所需产品的示例在本文别处描述。因此,生产产品的细菌细胞是本发明的又一方面。
184.在实施方案中,细菌细胞或经修饰的细菌细胞包含编码产品或编码生产产品所需的酶的基因。编码产品或酶的适当基因也在本文别处描述,例如产品(和编码基因)可以是
内源、同源或异源产品。
185.在实施方案中,该基因可操作地与启动子连接。适当的启动子的示例在本文别处描述,例如可以是原生或异源启动子,或者可以是组成型或诱导型启动子。
186.在另一方面,本发明提供了一种多核苷酸载体,其包含旁侧有位点特异性重组位点的复制起点。适当的和优选的位点特异性重组位点在本文别处描述。
187.在实施方案中,位点特异性重组位点包含用于重组酶,优选丝氨酸重组酶识别的位点特异性重组位点,优选地,其中所述丝氨酸重组酶为γδ、bxb1、或tp901。
188.在实施方案中,位点特异性重组位点包含用于酪氨酸重组酶识别的位点特异性重组位点,优选地,其中所述酪氨酸位点特异性重组酶为cre、dre、flp、kd、b2或b3。
189.在实施方案中,本发明的多核苷酸载体或细菌细胞(经修饰的细菌细胞)进一步包含用于选择不可逆(或永久)失活的复制起点的标记。
190.用于选择的标记通常是编码蛋白的核苷酸序列,该蛋白将选择性抗性赋予表达其的细胞。用于选择的标记优选是抗生素抗性基因。氯霉素抗性基因、氨苄青霉素抗性基因和卡那霉素抗性基因是优选的选择性标记,尽管许多其他选择性标记在本领域中有所描述并且可以使用。氯霉素抗性基因、氨苄青霉素抗性基因和卡那霉素抗性基因是本领域中常规使用的众所周知且充分表征的用于选择的标记。或者,用于选择的标记可以是赋予使用人工氮源能力的基因或赋予使用人工碳源能力的基因。选择性标记能够选择(或识别)已整合/并入不可逆(或永久)失活的复制起点的细胞。
191.在实施方案中,用于选择的标记与复制起点一起旁侧于位点特异性重组位点之间,以便作为将复制起点并入染色体(例如通过同源重组)的阳性标记。因此,在一些实施方案中,不可逆(或永久)失活的复制起点在5’到3’方向包括:(i)位点特异性重组位点,(ii)复制起点,(iii)阳性选择性标记基因,和(iv)位点特异性重组位点;或者,不可逆(或永久)失活的复制起点在5’到3’方向包括:(i)位点特异性重组位点,(ii)阳性选择性标记基因,(iii)复制起点,和(iv)位点特异性重组位点。
192.用于选择的标记可用于将失活的复制起点插入细菌染色体(例如通过同源重组或不同的方法)。然而,稍后可以去除用于选择的标记,同时在染色体中保留失活的复制起点(例如旁侧有重组位点的复制起点)。通常,用于选择的标记的存在对于本发明的细菌菌株(例如本发明的转换菌株)的下游应用不是必需的,例如对于产品生产不是必需的。因此,一旦将失活的复制起点并入细菌染色体中,通常就没有必要在细胞内保持选择性标记。或者,可以不使用用于选择的标记,例如使用另一种阳性选择工具,或根本不使用阳性选择工具来生成本发明的菌株(例如转换菌株)。
193.在实施方案中,不可逆(或永久)失活的复制起点可包含一种(或进一步的)用于选择的标记,其中该标记被安排成使得只有已转换(即已使其复制起点不可逆地(或永久地)失活)的本发明的细菌细胞可以表达该标记。例如,用于选择的标记可被安排成使得其被旁侧于复制起点的位点特异性重组位点打断,使得用于选择的标记的全部序列仅通过去除(或缺失或消除或切除)复制起点被连续组装。或者,用于选择的标记及其启动子可被安排成使得用于选择的标记及其启动子被旁侧于复制起点的位点特异性重组位点打断;以这种方式,只有发生转换(即不可逆地去除复制起点)才可操作地与用于选择的标记及其启动子连接(因此才可能表达用于选择的标记)。
194.在不可逆(或永久)失活的复制起点的实施方案中,阳性选择性标记基因旁侧有位点特异性重组位点,这些位点特异性重组位点可被位点特异性重组酶识别,但不包括复制起点(即可能存在两个位点特异性重组位点,一个位于阳性选择性标记基因的5’处,一个位于阳性选择性标记基因的3’处)。
195.在一些实施方案中,希望仅包括对已从基因组中进行了复制起点消除的细胞进行阳性选择。如果复制起点消除开启了生存所必需的基因的表达(例如抗生素抗性基因或必需营养素代谢所必需的基因),则只有已转换(或具有失活的复制起点)的细胞存活。仅对已消除复制起点的细胞使用阳性选择步骤提供了一种机制以减少或防止稀有突变体的存活,这些突变体出现并失去了响应适当的诱导条件(例如温度变化)而停止生长的能力,例如根据本发明,复制起点不再可去除或不再是不可逆失活的。此类突变体在本文中也称为“作弊者”,并有可能在诱导条件(例如37℃)下继续生长和繁殖,且可能成为培养物中的优势群体。在一些实施方案中,这种阳性选择步骤可以用来避免这种潜在的问题。
196.避免这种潜在的问题的另一种方法是通过使用多种(例如两种或更多种)位点特异性重组酶及其各自的位点特异性重组位点,如本文别处所述。
197.如上所述的本发明的细菌细胞可以很容易且方便地用于本发明的生产方法,例如能够生产产品(所需产品)的细胞。实际上,优选使用此类细胞,并且优选的方法包括提供能够生产所述产品的本发明的此类细菌细胞的步骤。
198.从上述讨论中还可以看出,有许多方法可用于不可逆地使细菌细胞中的复制起点失活的步骤,这些方法中的任何一种,或者实际上任何适当的替代方法,都可用于本发明的生产方法中的该步骤。优选的方法也如上所述,包括使用复制起点旁侧有重组位点(例如如上所述的位点特异性重组位点)的方法。
199.在本文所述的生产产品(或用于生产产品)的方法(产品生产方法)中,可以包括产品生产所需的任何进一步的步骤。例如,生产产品的细菌细胞可以通过本领域众所周知的适当方法进行培养、生长或以其他方式繁殖或生产,以便生产产品,例如适当或足够量的产品,或使细菌细胞群体得到生长或扩增。因此,该方法可进一步包括培养、生长或以其他方式繁殖细菌细胞的步骤。该方法还可进一步包括收集、分离、纯化或以其他方式从细菌细胞(例如从培养基(或上清液)或从细菌细胞的细胞内环境)中获得产品的步骤。同样,进行这些步骤的方法在本领域是众所周知的。
200.还提供了分离或纯化的细菌细胞(或细菌细胞群体)。在一些实施方案中,此类细菌细胞将不是或将不对应于天然存在的细菌菌株。一些实施方案将涉及进一步的步骤,即培养或繁殖或生产此类细菌菌株,并任选地将所述培养或繁殖或生产的菌株配制成包含所述菌株的组合物,例如用于产品生产的稳定配方,例如用于工业或商业产品生产,或配制成药物组合物。或者,可以储存此类细菌细胞以备将来使用,例如通过冻干或冷冻。
201.在另一方面,本发明提供了本发明的细菌细胞、经修饰的细菌细胞、或多核苷酸载体或方法用于生产产品(例如所需产品)的用途。示例性产品在本文别处描述。
202.由于本发明的细菌细胞(经修饰的细菌细胞)可以被设计成生产目标产品,包括治疗上有效和有用的产品,并且以有用的或显著的量,此类细菌具有明确的治疗用途,例如可以施用于受试者或患者,特别是人类患者,其中该产品的施用在治疗上是有效或有用的。细菌作为活体药物,例如益生菌的使用是本领域所接受的,本发明的细菌细胞在任何使用此
类合适或适当的活体药物的治疗中的用途都是本发明所考虑的。
203.因此,在另一方面,本发明提供了用于治疗的本发明的细菌细胞或本发明的经修饰的细菌细胞。
204.在另一方面,本发明提供了用于治疗或预防疾病或病理的本发明的细菌细胞或本发明的经修饰的细菌细胞。在优选的实施方案中,该疾病或病理为癌症、代谢疾病或免疫失调。
205.在另一方面,本发明提供了本发明的细菌细胞或本发明的经修饰的细菌细胞在制备用于治疗或预防疾病或病理的药物中的用途。在优选的实施方案中,该疾病或病理为癌症、代谢疾病或免疫失调。
206.在另一方面,本发明提供了一种治疗或预防疾病的方法,包括向受试者施用有效量的本发明的细菌细胞或本发明的经修饰的细菌细胞,优选地,其中该疾病或病理为癌症、代谢疾病或免疫失调。
207.在实施方案中,该疾病或病理为代谢疾病(或代谢紊乱)。优选地,代谢疾病(或代谢紊乱)为酸碱失衡、代谢性脑病、钙代谢紊乱、dna修复缺陷紊乱、葡萄糖代谢紊乱、高乳酸血症、铁代谢紊乱、脂质代谢紊乱、吸收不良综合征、代谢综合征、先天性代谢异常、线粒体疾病、磷代谢紊乱、卟啉症、蛋白稳态缺陷、代谢性皮肤病、消耗综合征(或恶病质)或水电解质失衡。
208.在实施方案中,该疾病或病理为免疫失调(或免疫疾病)。优选地,免疫失调(或免疫疾病)是过敏、哮喘、自身免疫性疾病(例如狼疮、硬皮病、溶血性贫血、血管炎、1型糖尿病、格雷夫斯病、类风湿性关节炎、多发性硬化症、肺出血肾炎综合征、恶性贫血,肌病或莱姆病)、自身炎症综合征或免疫缺陷综合征。
209.本文所用的术语“受试者”(或“患者”)包括任何哺乳动物,例如人类和任何牲畜、家养或实验室动物。具体示例包括小鼠、大鼠、猪、猫、狗、马、羊、兔、牛和猴(或其他灵长类动物)。然而,优选地,所述患者是人类受试者。
210.在与本文所述的治疗方法和用途有关的实施方案中,适当的受试者是指那些患有、疑似患有或有风险患有(或易患)待治疗疾病的。
211.本发明的所述治疗方法和用途中的细菌菌株是以药学、治疗或生理有效量施用于需要治疗的受试者。因此,所述方法和用途可能涉及识别需要治疗的受试者的额外步骤。
212.根据本发明的疾病的治疗(例如已存在疾病的治疗)包括治愈所述疾病或病症,或任何减轻或缓解疾病(例如减轻疾病的严重程度)或疾病的症状。
213.本发明的方法和用途适用于疾病的预防以及疾病的积极治疗(例如已存在疾病的治疗)。因此,预防性治疗也包括在本发明的范围内。为此,在本发明的方法和用途中,治疗还包括适当的预防或防止。
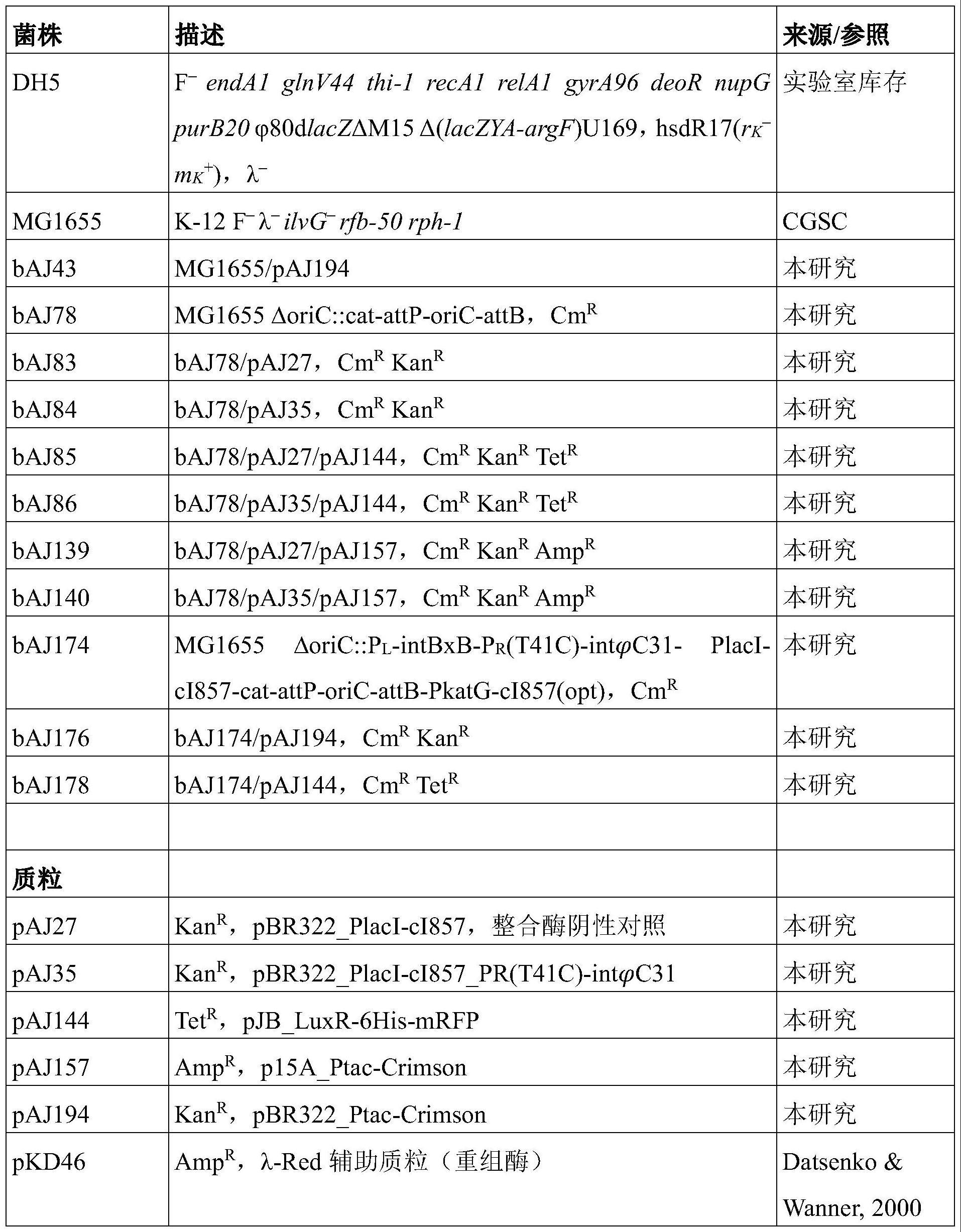
214.在实施方案中,细菌细胞为革兰氏阴性细菌细胞。或者,细菌细胞为革兰氏阳性细菌细胞。
215.在实施方案中,细菌细胞为埃希氏杆菌属、芽孢杆菌属、乳球菌属、链球菌属、乳杆菌属、棒状杆菌属、链霉菌属、假单胞菌属、梭菌属、黄单胞菌属或肠杆菌科。
216.在实施方案中,细菌细胞为埃希氏杆菌属,优选大肠杆菌,更优选大肠杆菌mg1655。
217.在实施方案中,细菌细胞为芽孢杆菌属,优选枯草芽孢杆菌。
218.如在整个申请中所使用的,术语“一”和“一个”是指“至少一个”、“至少第一个”、“一个或多个”或“多个”所提及的组分或步骤,除非在此后具体说明上限的情况下。
219.此外,如果本文使用术语“包含”、“包括”、“具有”或“含有”,或其他等同术语,则在一些更具体的实施方案中,这些术语包括术语“由...组成”或“基本上由...组成”,或其他等同术语。包括某些步骤的方法在适当时还包括由这些步骤组成的方法。
220.本文所述的术语“增加”或“提高”(或等同术语)包括当与适当的对照相比时任何可测量的增加或提高。本领域技术人员可以很容易地识别出适当的对照,且适当的示例在本文中有所描述。优选地,当与适当的对照水平或值相比时,增加将是显著的,例如临床或统计学上显著的,例如概率值≤0.05。
221.本文所述的术语“减少”或“降低”(或等同术语)包括当与适当的对照相比时任何可测量的减少或降低。本领域技术人员可以很容易地识别出适当的对照,且适当的示例在本文中有所描述。优选地,当与适当的对照水平或值相比时,减少将是显著的,例如临床或统计学上显著的,例如概率值≤0.05。
222.确定受试者测试组之间的差异或特定参数的水平或值的差异的统计显著性的方法在本领域是众所周知的,并有文献记载。例如,本文中如果使用显著性检验如t检验、曼-惠特尼u(秩和)检验、卡方检验或费舍尔精确检验、单因素方差分析或双因素方差分析检验(视情况而定)进行的统计比较显示概率值≤0.05,则通常将减少或增加视为统计显著。
223.现在将参照以下附图在以下非限制性实施例中进一步描述本发明。
224.图1。转换菌株的遗传基础。(a)丝氨酸重组酶识别位点attb和attp整合在细菌染色体中oric的两侧,其方向使得重组导致oric从细菌染色体上切除。(b)将丝氨酸重组酶置于λ噬菌体pr或p
l
启动子的控制下,该启动子受组成型表达的温度敏感性λ噬菌体ci阻遏因子的阻遏。(c)双重组酶设置,其中两种重组酶都在λ噬菌体ci阻遏因子的控制下。
225.图2。温度诱导的整合酶的表达导致转换细胞中oric的切除。对照组(baj83,空心符号)和转换组(baj84,实心符号)培养物由过夜培养物稀释后于30℃(白色绘图区域)在摇瓶中生长两小时。在时间点0小时,温度改变为37℃(灰色绘图区域)。(a)测量温度变化前后的菌落形成单位数量。绘制三个独立实验的几何平均值,误差线表示标准偏差。(b)使用未重组(oric保留)或重组(oric切除)dna序列特异性引物,通过pcr检测oric从染色体dna上切除的情况,并通过琼脂糖凝胶电泳进行可视化。(c)在整合酶切除oric后,attb和attp位点之间形成的连接的dna序列。
226.图3。最终细胞密度可以通过转换的时间来调整。将对照菌株(baj83,空心符号)和转换菌株(baj84,实心符号)的过夜培养物的各种稀释液在30℃(白色绘图区域)的微孔板培养箱上接种。在时间点0小时,温度改变为37℃(灰色绘图区域),以诱导转换。转换细胞在产生不同水平的生物量(600nm处的光密度)时停止生长,具体取决于转换时的密度。绘制两个独立实验的六个生物重复的平均值,阴影表示标准偏差。
227.图4。与对照菌株相比,转换菌株的蛋白质生产能力的评估。对照组(baj85,空心符号)和转换组(baj86,实心符号)培养物在30℃(白色绘图区域)的微孔板培养箱上预生长,以不同的稀释度(a为2400倍,b为1600倍)从过夜培养物中接种。在时间点0小时,温度改变为37℃(灰色绘图区域),以诱导转换菌株中oric的切除。通过在时间点0小时添加hsl来诱
导mrfp1荧光蛋白的产生。(a、b)监测mrfp1蛋白在600nm处的od变化(左轴)以及荧光(激发波长:584nm,发射波长:607nm)增加(右轴)。(c、d)对照组(空心)和转换组(实心)培养物的荧光与光密度(生物量)之比基于图a和b的值计算并绘制(c为2400倍,d为1600倍)。绘制三个生物重复的平均值,阴影表示标准偏差。
228.图5。转换菌株的蛋白质生产能力在很长一段时间内保持活跃。对照组(baj85,空心符号)和转换组(baj86,实心符号)培养物在30℃(白色绘图区域)的微孔板培养箱上预生长。在时间点0小时,温度改变为37℃(灰色绘图区域),以诱导转换菌株中oric的切除。通过在指定时间点向对照组和转换组培养物中添加hsl来诱导mrfp1荧光报告蛋白的产生。监测光密度(a)和荧光(b,激发波长:584nm,发射波长:607nm)的变化。绘制三个生物重复的平均值,阴影(图b)表示标准偏差。
229.图6。使用的dna构建体和质粒的示意图。a.并入转换菌株的基因组的线性dna片段。基因组中mnmg和mioc基因之间的区域被f_baj78 dna片段替换。t
–
转录终止子,t_bi
–
双向转录终止子,frt
–
翻转酶识别靶位点,cat
–
氯霉素抗性基因,gfpmut2
–
gfp报告基因。b.本研究中使用的质粒图谱。λpr(t41c)
–
λ噬菌体pr(t41c)启动子,bom、rop、trfa、oriv、rk2
–
质粒dna复制起点元件,pbr322_origin
–
质粒复制起点,teta、tetr
–
四环素抗性基因,kanr
–
卡那霉素抗性基因,lux-box
–
受luxr调控的启动子。
230.图7a。与对照菌株相比,转换菌株的蛋白质生产能力的评估。对照组(baj139,空心符号)和转换组(baj140,实心符号)培养物在30℃(白色绘图区域)的微孔板培养箱上预生长,以2000倍稀释度从过夜培养物中接种。在时间点0小时,温度改变为37℃(灰色绘图区域),以诱导转换菌株中oric的切除。通过在时间点0小时添加iptg来诱导e2-crimson荧光蛋白的产生。监测e2-crimson蛋白在600nm处的od变化(左轴)以及荧光(激发波长:610nm,发射波长:650nm)增加(右轴)。绘制三个生物重复的平均值,阴影表示标准偏差。
231.图7b。与对照菌株相比,基因组转换菌株的蛋白质生产能力的评估。对照组(baj85,空心符号)和转换组(baj178,实心符号)培养物在30℃(白色绘图区域)的微孔板培养箱上预生长,以8000倍稀释度从过夜培养物中接种。在时间点0小时,温度改变为37℃(灰色绘图区域),以诱导基因组转换菌株中oric的切除。通过在时间点0小时添加hsl来诱导mrfp1荧光蛋白的产生。监测mrfp1蛋白在600nm处的od变化(左轴)以及荧光(激发波长:584nm,发射波长:607nm)增加(右轴)。绘制三个生物重复的平均值,阴影表示标准偏差。
232.图7c。与对照菌株相比,基因组转换菌株的蛋白质生产能力的评估。对照组(baj43,空心符号)和转换组(baj176,实心符号)培养物在30℃(白色绘图区域)的微孔板培养箱上预生长,以16000倍稀释度从过夜培养物中接种。在时间点0小时,温度改变为37℃(灰色绘图区域),以诱导全基因组转换菌株中oric的切除。通过在时间点0小时添加iptg来诱导e2-crimson荧光蛋白的产生。监测e2-crimson蛋白在600nm处的od变化(左轴)以及荧光(激发波长:610nm,发射波长:650nm)增加(右轴)。绘制三个生物重复的平均值,阴影表示标准偏差。
233.图8。使用的dna构建体和质粒的示意图。a.并入基因组转换菌株baj174的基因组的线性dna片段。基因组中mnmg和mioc基因之间的区域被baj174 dna片段替换。t
–
转录终止子,t_bi
–
双向转录终止子,frt
–
翻转酶识别靶位点,cat
–
氯霉素抗性基因,gfpmut2
–
gfp报告基因。b.本研究中使用的质粒图谱。bom、rop
–
质粒dna复制起点元件,p15a_orign、
pbr322_origin
–
质粒复制起点,lac操纵子
–
laci结合序列,ampr
–
氨苄青霉素抗性基因,kanr
–
卡那霉素抗性基因。
234.实施例1
235.材料和方法
236.细菌菌株、质粒和生长培养基
237.菌株和质粒列于表1。大肠杆菌dh5α用于质粒克隆和繁殖。在大肠杆菌mg1655中进行了基因组改变和基于质粒的转换实验。大肠杆菌在溶原性肉汤(lb)中生长,必要时添加适量抗生素(100μg/ml氨苄青霉素、25μg/ml氯霉素、25μg/ml卡那霉素、10μg/ml四环素),以选择菌株和维持质粒。
238.dna操作
239.用于克隆和测序的短寡核苷酸序列购自metabion international ag。
240.使用重组工程进行大肠杆菌mg1655的基因组原位工程。λ噬菌体red重组系统由pkd46质粒表达(datsenko and wanner,2000)。旁侧有attb和attp位点的oric片段(表2)作为合成dna购自twist bioscience。从大肠杆菌mg1655的基因组dna中扩增出侧翼的400bp长的同源区。从pkd3中扩增出具有相邻frt位点的氯霉素抗性标记。使用重叠延伸pcr组装重组工程片段f_baj78(最终序列列于表3、图6a),进行凝胶纯化,并通过电穿孔转化为大肠杆菌细胞。
241.为了构建基因组转换菌株,整合酶、具有λp
l
启动子的bxb1整合酶、具有pkatg启动子的ci857阻遏因子、attb和attp位点以及cat选择性标记的密码子优化序列作为合成dna片段购自twist bioscience(表2、5)。通过pcr从大肠杆菌mg1655的基因组dna中扩增出重组工程所需的oric和同源区。通过重叠延伸pcr组装片段,进行凝胶纯化,并用于重组工程(datsenko and wanner,2000)(整合到基因组中的baj174的最终序列列于表3、图8a)。
242.质粒paj35用于在转换菌株中表达整合酶。在paj35中,整合酶(merrick,zhao and rosser,2018)置于突变的λpr(t41c)启动子的控制下(表2;(jechlinger et al.,1999)),其活性受pet24骨架中由组成型启动子表达的λ噬菌体阻遏因子ci587的温度敏感性突变体控制(表4、图6b)。在对照菌株中,除了整合酶orf被翻译为mdtyagaydrqsrerenssaaspatqrsa的小orf取代之外,含有与paj35所有相同元件的质粒paj27用作阴性对照(表4、图6b)。
243.为了测定蛋白合成活性,将红色荧光蛋白mrfp1(campbell et al.,2002)置于质粒paj144中hsl诱导型lux启动子的控制下(表4、图6b)。
244.为了测定iptg诱导的蛋白合成活性,将荧光蛋白e2-crimson置于具有pbr322复制起点(paj194)或p15a复制起点(paj157)的质粒中iptg诱导型ptac启动子的控制下(表4、图8b)。
245.验证oric切除
246.转换菌株和对照菌株的培养物于30℃下生长过夜,稀释2000倍到20ml新鲜lb中,并于30℃下生长2小时。此后将温度转变为37℃。每小时取样,对lb琼脂平板上每体积培养物的菌落形成单位(cfu/ml)进行计数,并通过pcr分析oric的切除情况。模板的量按od值进行归一化。使用特异性引物对检测oric的切除(oaj297和oaj32)或未切除(oaj91和oaj32)版本。通过测序验证oric切除版本的dna序列。
247.生长和荧光测量
248.转换菌株和对照菌株的培养物于30℃下生长过夜,如图例所示稀释,在96孔板上于30℃下继续生长3小时(对于基因组转换菌株)或4小时,直到温度转变为37℃(时间点0小时)。在适当的情况下,通过在温度转变前立即或在指定时间点添加12m hsl(最终浓度)来诱导mrfp1的合成。使用biotek synergy mx酶标仪测定生长曲线和mrfp1相关荧光(反映蛋白水平)。在100μl体积的培养物中测量600nm处的光密度和荧光强度(激发波长为584/13,5nm;发射波长为607/13,5nm)。通过在温度转变前立即添加0.5mm iptg(最终浓度)来诱导e2-crimson的合成,并在100μl体积的培养物中测量荧光强度(激发波长为610/20nm;发射波长为650/20nm)。
249.表1:菌株和质粒
250.251.a cmr:氯霉素抗性;kanr:卡那霉素抗性;ampr:氨苄青霉素抗性;tetr:四环素抗性
252.表2:功能元件的核苷酸序列和寡核苷酸序列
[0253][0254]at41c突变以粗体表示
[0255]
表3:转换菌株的oric区的序列
[0256]
[0257]
[0258]
[0259]
[0260]
[0261]
[0262]
[0263]
[0264][0265]
表4:质粒序列
[0266]
[0267]
[0268]
[0269]
[0270]
[0271]
[0272]
[0273]
[0274]
[0275]
[0276]
[0277]
[0278]
[0279]
[0280]
[0281]
[0282]
[0283]
[0284]
[0285]
[0286]
[0287]
[0288]
[0289][0290]
表5:密码子优化蛋白的序列
[0291]
[0292]
[0293][0294]
结果
[0295]
dna复制是所有生物体中细胞分裂的先决条件。在细菌中,染色体携带复制起点(ori)的单一dna序列,复制复合物在该序列上组装并开始复制。在大肠杆菌中,ori序列称为oric,其在普通实验室菌株中位于mnmg和mioc基因之间(messer,2002)。
[0296]
我们试图通过抑制其dna复制来阻断细胞生长和分裂。为了实现这一点,我们在oric的两侧整合了丝氨酸重组酶识别位点attb和attp(图1a)。当相应的丝氨酸重组酶在细
胞中表达时,其会在位点上催化切割dna,通过链交换切除其间的dna序列,并重新连接dna以形成基本上不可逆的杂合att位点(merrick,zhao and rosser,2018)。结果,oric从细菌染色体上消除。以下,在本实施例部分和本文别处,我们将oric的切除称为“转换”或“已转换的”或“转换子”,指的是能够转换的菌株。oric的切除将排除任何进一步的dna复制起始,同时允许正在进行的复制完成。在这些实施例中,我们使用丝氨酸重组酶来构建转换,但很明显,其他类型的基因组编辑也可用于相同目的。可能的候选方案包括但不限于酪氨酸重组酶(fogg et al.,2014)、crispr/cas(yao et al.,2018)或任何其他类型的酶,这些酶可永久地改变染色体中的oric序列,并使其无法起始dna复制。
[0297]
基本上,丝氨酸重组酶(或其他类型的编辑)的活性/表达受到严格控制,以避免过早消除oric和停止生长。为此,我们使用了温度敏感性λ噬菌体ci857阻遏因子(图1b)。在30℃(或更低)下,该阻遏因子与其目标启动子(pr或p
l
)结合并对其进行阻遏,而在37℃(或更高)时,阻遏被解除,且表达开始。其他类型的基因表达控制方法也可用于相同目的,包括但不限于诱导剂(iptg、阿拉伯糖、高丝氨酸内酯、无水四环素等)诱导(marschall,sagmeister and herwig,2017)(lutz and bujard,1997)(cox,surette and elowitz,2007)、靶向蛋白水解(cameron and collins,2014)、crispr/cas9的不同变体(qi et al.,2013)等。
[0298]
我们构建了一种大肠杆菌菌株,该菌株携带经修饰的oric附近(图1a)和丝氨酸重组酶表达系统,在附加体质粒上表达(尽管丝氨酸重组酶表达系统同样可以整合到基因组中——如图7b和7c所评估的)。在我们的系统中,我们可以在同一ci阻遏因子的控制下,以单一或双重设置表达不同的丝氨酸重组酶(图1b和1c)。如有必要,可以添加具有相同或不同的表达控制机制的额外重组酶。我们在实验中使用了整合酶,但很明显,其他丝氨酸重组酶,连同其同源attp和attb位点,也可用于相同目的。实验中描述了数十种不同的丝氨酸重组酶,生物信息学预测了超过4000种新的丝氨酸重组酶(yang et al.,2014)。
[0299]
有可能出现稀有突变体,这些突变体失去了响应温度变化而停止生长的能力。此类突变体(“作弊者”)将在37℃下继续生长和繁殖,并可能成为培养物中的优势群体。为了避免这种潜在的问题,可以仅包括对已从基因组中进行了复制起点(oric)消除的细胞进行阳性选择。如果后者开启了生存所必需的基因的表达(例如抗生素抗性基因或必需营养素代谢所必需的基因或人工氮源代谢所必需的基因),则只有已转换的细胞才能存活。
[0300]
我们在时间进程实验中测试了转换子从基因组中消除oric的能力,首先在30℃下培养细胞(无切除),然后将培养物转移至37℃(切除)(图2)。温度转变后,转换菌株的cfu/ml数量下降了两个数量级以上(图2a),且重组迅速消除了oric(图2b)。在缺乏活性重组酶的对照菌株(对照)中,这些变化均未发生。为了进一步确认oric的切除,我们对已转换菌株中attb和attp位点之间的连接处进行了测序,并验证了其与预测完全一致(图2c)。
[0301]
研究发现,oric的切除和基因组复制的消除最终导致了细胞分裂和生长的停止。为了研究间歇培养系统中细胞密度的变化,我们在不同的细胞密度下诱导了转换(图3)。转换后,od值继续增加了几个小时,然后开始放缓。根据初始细胞密度,转换菌株逐渐达到明显的平台期密度(图3中对照菌株平台期的大约65%、95%和110%),该平台期密度甚至可以高于对照细胞。转换培养物的最终密度低于对照培养物,表明培养基中的营养物质并未耗尽,并且转换培养物的生长可通过其他方式停止。这使我们可以通过在转换时选择适当
glutamicum’,microbial cell factories.biomed central ltd.,17(1),p.185.doi:10.1186/s12934-018-1031-7.
[0321]
madsen,s.m.et al.(1999)
‘
molecular characterization of the ph-inducible and growth phase-dependent promoter p170 of lactococcus lactis’,molecular microbiology.blackwell publishing ltd.,32(1),pp.75
–
87.doi:10.1046/j.1365-2958.1999.01326.x.
[0322]
marschall,l.,sagmeister,p.and herwig,c.(2017)
‘
tunable recombinant proteinexpression in e.coli:promoter systems and genetic constraints’,applied microbiology andbiotechnology.springer verlag,101(2),pp.501
–
512.doi:10.1007/s00253-016-8045-z.
[0323]
merrick,c.,zhao,j.and rosser,s.(2018)
‘
serine integrases:advancing syntheticbiology’,acs synthetic biology.acs.doi:10.1021/acssynbio.7b00308.
[0324]
messer,w.(2002)
‘
the bacterial replication initiator dnaa.dnaa and oric,the bacterialmode to initiate dna replication tl-26’,fems microbiology reviews.wiley,26 vn-r(4),pp.355
–
374.doi:10.1111/j.1574-6976.2002.tb00620.x.
[0325]
paul,s.et al.(2004)
‘
autoinduction of bacillus subtilis phopr operon transcription resultsfrom enhanced transcription from eσa-and eσe-responsive promoters by phosphorylatedphop’,journal of bacteriology.american society for microbiology journals,186(13),pp.4262
–
4275.doi:10.1128/jb.186.13.4262-4275.2004.
[0326]
qi,l.s.et al.(2013)
‘
repurposing crispr as an rna-γuided platform for sequence-specific control of gene expression’,cell.cell press,152(5),pp.1173
–
1183.doi:10.1016/j.cell.2013.02.022.
[0327]
soma,y.et al.(2014)
‘
metabolic flux redirection from a central metabolic pathway towarda synthetic pathway using a metabolic toggle switch.’,metab.eng.doi:10.1016/j.ymben.2014.02.008.
[0328]
venayak,n.et al.(2015)
‘
engineering metabolism through dynamic control’,curr opinbiotech,34,pp.142
–
152.doi:10.1016/j.copbio.2014.12.022.
[0329]
wang,j.et al.(2018)
‘
a novel process for cadaverine bio-production using a consortiumof two engineered escherichia coli’,frontiers in microbiology.frontiers media s.a.,9(jun),p.1312.doi:10.3389/fmicb.2018.01312.
[0330]
yang,l.et al.(2014)
‘
permanent genetic memory with》1-byte capacity’,nature methods.nature publishing group,11(12),pp.1261
–
1266.doi:10.1038/nmeth.3147.
[0331]
yao,r.et al.(2018)
‘
crispr-cas9/cas12a biotechnology and application in bacteria’,synthetic and systems biotechnology.keai communications co.,pp.135
–
149.doi:10.1016/j.synbio.2018.09.004.
[0332]
yuan,s.f.et al.(2020)
‘
de novo resveratrol production through modular engineering ofan escherichia coli-saccharomyces cerevisiae co-culture’,
microbial cell factories.biomedcentral,19(1),p.143.doi:10.1186/s12934-020-01401-5.
[0333]
strack,r.l.et al.(2009)
‘
a rapidly maturing far-red derivative of dsred-express2 forwhole-cell labeling.’,biochemistry,48(35),pp.8279
–
8281.doi:10.1021/bi900870u.
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1