牛骨骼肌分化相关DNA甲基化酶TET1与m6A甲基化酶METTL3的互作鉴定和应用
牛骨骼肌分化相关dna甲基化酶tet1与m6a甲基化酶mettl3的互作鉴定和应用
技术领域
1.本发明属于分子生物学技术领域,涉及dna甲基化修饰与rna甲基化修饰互作的鉴定、功能及应用,具体涉及牛骨骼肌分化相关dna甲基化酶tet1和m6a甲基转移酶mettl3的互作鉴定、功能及应用。
背景技术:
2.研究表明,畜禽的产肉量和肉品质离不开骨骼肌的生长发育及其遗传特性,一方面,骨骼肌肌纤维的数量和直径是决定动物产肉量的关键因素;另一方面,骨骼肌中肌内脂肪含量、肌纤维内蛋白质种类和数量与肌肉的嫩度密切相关,并影响肉品质。除此以外,骨骼肌是机体重要的运动和能量代谢组织,对于维持机体代谢平衡和稳态起到重要作用。因此,研究骨骼肌的生长发育规律对于提高肉用动物生产性能和研究肌肉生理病理学十分必要。同时,由于我国肉牛业起步较晚,本土的地方黄牛品种一直饱受生长速度慢、产肉性能较国外优良肉牛品种差距较大等因素的困扰,如何提高地方黄牛的产肉量和肉品质是国内肉牛育种工作者亟待解决的共同问题。
3.肌生成是一个非常复杂的生物学过程,从肌卫星细胞激活到成肌细胞增殖最终到终末分化阶段,这一过程除了受到关键的转录因子调控外,一些表观遗传如dna甲基化、组蛋白修饰等也发挥着重要的作用。而m6a作为真核生物最为普遍的mrna甲基化修饰,具有动态可逆性,由m6a甲基转移酶复合物核心组分mettl3、mettl14和wtap催化形成,同时能够被m6a去甲基化酶fto和alkbh5去除。哺乳动物中mrna上m6a甲基化修饰的基序通常具有物种保守性,一般发生为“rrach”序列,其中r表示鸟嘌呤(g)或者腺嘌呤(a);h表示腺嘌呤(a)、胞嘧啶(c)或尿嘧啶(u)。m6a修饰通过介导mrna代谢,包括影响rna翻译效率、稳定、剪切等调控多种生物学过程。近年来的研究已经发现m6a修饰参与肌肉生长发育。在小鼠骨骼肌原代成肌细胞和c2c12细胞中,m6a修饰调控其成肌分化。同时,随着m6a特异性甲基化rna免疫沉淀高通量测序技术(merip-seq/m6a-seq)的发明,越来越多的研究表明m6a修饰在调控猪、牛、鸡、鹅、羊等畜禽的脂肪和肌肉发育等方面有重要作用。然而,关于m6a修饰在牛骨骼肌生成中的功能、作用机制以及潜在靶基因仍未报道,关于m6a甲基化与其他表观遗传修饰的相互关系也鲜少发现。
4.众所周知,dna甲基化对骨骼肌分化过程来说是必不可少的。tet1是一种dna去甲基化酶,根据前期的m6a测序发现tet1 mrna m6a甲基化水平在秦川牛骨骼肌成肌细胞分化前后显著差异,表明tet1可能受到m6a甲基化修饰的调控,同时暗示了dna甲基化和rna甲基化在骨骼肌分化中存在潜在的互作机制。因此,有必要对tet1 mrna的m6a甲基化位点及与mettl3的互作调控功能进行分析鉴定,为研究m6a调控肌肉生长发育的功能研究提供参考,为肉牛的定向选育提供一种新的分子标记。
技术实现要素:
5.本发明的目的在于,提供一种牛骨骼肌分化相关tet1 mrna m6a甲基化位点鉴定方法、一种改变tet1 mrna的m6a甲基化修饰水平的方法、一种检测dna去甲基化酶tet1受m6a甲基化修饰调控的方法、一种tet1 mrna m6a甲基化修饰影响骨骼肌分化的鉴定方法、一种检测dna去甲基化酶tet1影响m6a甲基化修饰的方法。
6.为了实现上述任务,本发明采取如下的技术解决方案:
7.一种tet1 mrna m6a甲基化位点,tet1 mrna在ensembl数据库中的序列号为ensbtat00000010614.6,其特征在于,所述tet1 mrna m6a甲基化位点位于ensbtat00000010614.6序列的第3999位核苷酸,并通过m6a-ip-qpcr验证tet1 mrna的m6a甲基化水平,用于作为预测牛骨骼肌分化的分子标记。
8.一种与牛骨骼肌分化相关tet1 mrna m6a甲基化位点鉴定方法,根据在秦川牛骨骼肌成肌细胞分化过程中的m6a-seq测序结果分析得到相关rna片段中存在tet1 mrna的m6a甲基化修饰位点,其特征在于:
9.(1)根据哺乳动物中高度保守的rrach(r=g,a;h=a,c,u)序列进行tet1 mrna上m6a位点位置的确定;
10.(2)根据测序结果中的m6a甲基化位点在tet1 mrna上的位置,设计并合成含有m6a位点的定量引物:
11.tet1-m6a-f:5
’‑
ctccaaccaaccagtgtgct-3’12.tet1-m6a-r:5
’‑
ggtctggtaacggggtctca-3’13.通过m6a免疫沉淀后的实时荧光定量pcr检测tet1 mrna的m6a甲基化水平在牛成肌细胞分化过程中的变化,进而精确验证m6a-seq测序的结果,进一步证明tet1 mrna m6a甲基化位点在ensembl数据库中的序列号为ensbtat00000010614.6,位于ensbtat00000010614.6序列的第3999位核苷酸。
14.一种改变tet1 mrna m6a甲基化修饰水平的方法,其特征在于:所述tet1 mrna在ensembl数据库中的序列号为ensbtat00000010614.6,所述tet1 mrna m6a甲基化位点位于ensbtat00000010614.6序列的第3999位核苷酸,将ensbtat00000010614.6序列的第3999位腺苷酸(a),同义突变为鸟嘌呤(g)。
15.一种检测tet1 mrna受m6a甲基化调控的方法,其特征在于:
16.(1)将tet1 mrna m6a甲基化位点由腺苷酸(a)同义突变为鸟嘌呤(g)后,分别构建野生型和突变型tet1双荧光素酶表达质粒,利用双荧光素酶报告系统分别检测m6a甲基转移酶mettl3、m6a阅读蛋白ytdhf2与tet1 mrna的结合;
17.(2)利用rna结合蛋白免疫共沉淀技术验证ythdf2蛋白与tet1 mrna m6a甲基化位点区域的结合。
18.一种tet1 mrna m6a甲基化修饰影响骨骼肌分化的鉴定方法,其特征在于:
19.(1)分别将tet1-wt和tet1-mut表达质粒转染进牛骨骼肌成肌细胞;
20.(2)24小时后,诱导骨骼肌成肌细胞进行成肌分化;
21.(3)利用免疫荧光和rt-qpcr技术检验tet1-wt和tet1-mut对成肌分化的影响,证明tet1 mrna的m6a甲基化修饰对骨骼肌成肌分化具有促进作用。
22.一种检测dna去甲基化酶tet1影响m6a甲基化修饰的方法,其特征在于:
23.(1)分别将tet1-wt和tet1-mut表达质粒转染进牛骨骼肌成肌细胞,检测tet1 mrna的m6a甲基化修饰影响mettl3的表达;
24.(2)利用甲基化岛分析网站methprimer,设计mettl3启动子中cpg岛的特异性亚硫酸氢盐测序pcr(bsp)引物:
25.mettl3-bsp-f:5
’‑
ttttgggagttggtgatggatagatag-3’26.mettl3-bsp-r:5
’‑
atcccaacaaatactataaaaaaaa-3’27.(3)通过亚硫酸氢盐测序技术检测tet1影响mettl3启动子的dna甲基化水平,调控mettl3的表达,进而影响m6a水平。
28.本发明带来的有益技术效果在于:
29.1、利用对m6a-seq的结果分析,根据哺乳动物中m6a的保守序列,鉴定了m6a在tet1 mrna上的具体位点;
30.2、通过同义突变m6a位点,在不改变氨基酸序列前提下改变了tet1 mrna的m6a水平,并发现tet1表达随之改变,证明了tet1 mrna的m6a甲基化位点影响骨骼肌生成;但这些并不限于tet1基因,其他发生m6a甲基化修饰的基因同样适用此方法;
31.3、证明了tet1的表达受到mettl3-m6a-ythdf2的调控,表明m6a甲基化修饰在骨骼肌生成中影响dna甲基化;
32.4、鉴定了tet1也可以影响m6a甲基转移酶mettl3的表达,证明了dna甲基化也可以影响rna m6a甲基化水平。
33.5、为肉牛的定向选育和分子育种提供有益的基因资源,鉴定了骨骼肌发育中一种新的互作机制,为肌肉相关机理和病理提供新的潜在分子标记和治疗靶点。
附图说明
34.图1:秦川牛tet1 mrna m6a甲基化位点和在骨骼肌成肌分化过程中甲基化水平的差异;
35.图2:秦川牛tet1 mrna m6a甲基化位点在mrna上的具体位置及同义点突变;
36.图3:tet1 mrna m6a甲基化位点突变后对骨骼肌成肌分化的影响;
37.图4:tet1 mrna m6a点突变后对tet1 mrna稳定性的影响;
38.图5:tet1 mrna m6a甲基化位点突变后影响tet1的表达;
39.图6:tet1 mrna m6a甲基化位点突变影响mettl3/ythdf2与tet1 mrna的结合;
40.图7:tet1影响mettl3的表达及其启动子的dna甲基化水平。
41.以下结合附图和实施例对本发明作进一步的详细说明。
具体实施方式
42.本实施例给出一种tet1 mrna m6a甲基化位点,tet1 mrna在ensembl数据库中的序列号为ensbtat00000010614.6,所述tet1 mrna m6a甲基化位点位于ensbtat00000010614.6序列的第3999位核苷酸,并通过m6a-ip-qpcr验证tet1 mrna的m6a甲基化水平,用于作为预测牛骨骼肌生成分子标记。
43.一种与牛骨骼肌分化相关tet1 mrna m6a甲基化位点鉴定方法,该方法根据在秦川牛骨骼肌成肌细胞分化过程中的m6a-seq测序结果,按以下步骤分析得到相关rna片段中
存在m6a甲基化修饰位点:
44.(1)根据哺乳动物中高度保守的rrach(r=g,a;h=a,c,u)序列进行tet1 mrna上m6a位点位置的确定;
45.(2)根据测序结果中的m6a甲基化位点在tet1 mrna上的位置,设计并合成含有m6a位点的定量引物:
46.tet1-m6a-f:5
’‑
ctccaaccaaccagtgtgct-3’47.tet1-m6a-r:5
’‑
ggtctggtaacggggtctca-3’48.通过m6a免疫沉淀后的实时荧光定量pcr(m6a-ip-qpcr)检测tet1 mrna的m6a甲基化水平在牛成肌细胞分化过程中的变化,进而精确验证了m6a-seq测序的结果,进一步证明tet1 mrna m6a甲基化位点的准确性。该tet1 mrna m6a甲基化位点在ensembl数据库中的序列号为ensbtat00000010614.6,位于ensbtat00000010614.6序列的第3999位核苷酸。
49.一种检测dna去甲基化酶tet1受m6a甲基化修饰调控的方法,包括:
50.(1)通过双荧光素酶报告系统检测m6a甲基转移酶mettl3与tet1 mrna m6a位点的结合,精确证明mettl3通过结合tet1 mrna,催化其形成m6a甲基化;
51.(2)m6a甲基化修饰发挥的具体作用一般是通过m6a阅读蛋白来表征,根据所述的改变tet1 mrna的m6a甲基化修饰水平的方法,进行tet1 mrna m6a甲基化位点对tet1 mrna稳定性的影响:
52.1)根据所述的改变tet1 mrna的m6a甲基化修饰水平的方法,将tet1基因中受到m6a修饰的第3999位腺苷酸(a)同义突变为鸟嘌呤(g)后,将野生型和同义突变后的tet1编码区序列克隆进表达载体中分别构建野生型(tet1-wt)和m6a同义突变型(tet1-mut)表达质粒;
53.2)分别将tet1-wt和tet1-mut表达质粒转染进牛骨骼肌成肌细胞;
54.3)24小时后,在细胞培养液中加入5μg/ml的放线菌素d(抑制mrna的转录),在处理后0、3、6、9个小时收集细胞,提取细胞总rna,进行反转录,利用rt-qpcr技术检测tet1的mrna表达量,比较tet1-wt组和tet1-mut组中tet1 mrna在放线菌素d处理后的表达情况,计算tet1 mrna稳定性,证明了m6a甲基化位点突变后提高了tet1mrna的稳定性,因此,推测m6a甲基化修饰抑制tet1表达;
55.4)m6a阅读蛋白ythdf2一般被认为识别靶标转录本的m6a甲基化位点,降低靶标的稳定性,促进其mrna降解。因此,利用双荧光素酶报告系统和rna结合蛋白免疫沉淀实验(rip)验证m6a阅读蛋白ythdf2与tet1 mrna的m6a位点结合,进而发挥抑制tet1表达的作用;
56.5)通过上述步骤证明了tet1的表达受到mettl3-m6a-ythdf2轴的调控,即确定了dna去甲基化酶tet1的表达受m6a甲基化修饰调控。
57.一种tet1 mrna m6a甲基化修饰影响骨骼肌分化的鉴定方法,包括:
58.(1)根据上述方法,分别将tet1-wt和tet1-mut表达质粒转染进牛骨骼肌成肌细胞;
59.(2)24小时后,诱导骨骼肌成肌细胞进行成肌分化;
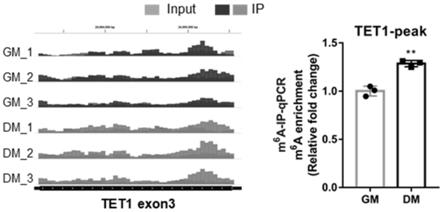
60.(3)利用免疫荧光和rt-qpcr技术检验tet1-wt和tet1-mut对成肌分化的影响,从而精确证明了tet1 mrna的m6a甲基化修饰对骨骼肌成肌分化具有促进作用。
61.tet1作为一个dna去甲基化酶,通过将靶基因的dna甲基化5mc转变成5hmc,达到dna去甲基化的作用。因此,猜测tet1也可以介导mettl3的dna甲基化水平。
62.一种检测dna去甲基化酶tet1影响m6a甲基化修饰的方法,包括:
63.(1)根据上述方法,分别将tet1-wt和tet1-mut表达质粒转染进牛骨骼肌成肌细胞,检测了tet1mrna的m6a甲基化修饰影响mettl3的表达;
64.(2)利用甲基化序列位置分析网站methprimer(http://www.urogene.org/cgi-bin/methprimer2),设计mettl3启动子中cpg岛的特异性亚硫酸氢盐测序pcr(bsp)引物:
65.mettl3-bsp-f:5
’‑
ttttgggagttggtgatggatagatag-3’66.mettl3-bsp-r:5
’‑
atcccaacaaatactataaaaaaaa-3’67.(3)通过亚硫酸氢盐测序技术检测tet1影响mettl3启动子的dna甲基化水平,调控mettl3的表达,进而影响m6a水平。
68.以下是发明人给出的具体实施例。
69.实施例1:牛骨骼肌成肌细胞分化过程中tet1mrnam6a甲基化水平的差异
70.试验材料为秦川牛成肌细胞分化前后m6a-seq测序后的数据和剩余的m6a免疫沉淀后的mrna样品。
71.具体操作如下:
72.1、m6a-seq测序数据的分析和可视化
73.(1)对测序数据进一步进行生物信息学分析,在igv软件中将tet1上的m6apeak峰图可视化,表明成肌分化前后tet1mrna的m6a甲基化位置以及m6a甲基化水平存在差异(图1左图);
74.(2)根据测序结果中tet1的m6apeak序列,在m6a修饰位点预测网站(http://www.cuilab.cn/sramp)确定tet1mrna的m6a位点位于编码区第3999位核苷酸;
75.2、利用m6a-ip-qpcr方法检测并验证tet1mrnam6a甲基化水平
76.(1)将测序后剩余的m6a免疫沉淀后的mrna反转录成cdna;
77.(2)根据上述测序结果中的m6a甲基化位点在tet1mrna上的位置,在上下游设计含有m6a位点的定量引物,有上海生工生物有限公司合成,序列如下:
78.tet1-m6a-f:5
’‑
ctccaaccaaccagtgtgct-3’79.tet1-m6a-r:5
’‑
ggtctggtaacggggtctca-3’80.(3)进行m6a-ip-qpcr检测tet1mrna的m6a甲基化水平变化,发现tet1的m6a甲基化水平在成肌细胞分化后升高(图1右图,gm表示分化前,dm表示成肌分化第4天)。
81.实施例2:改变tet1mrna的m6a甲基化修饰的方法,即对m6a位点进行同义突变;
82.牛tet1基因的编码区核苷酸序列如下:
83.atgtctcgatctcgccacgcaagaccttccaaattagtcaggagggaagatctaaacaag60
84.aagaaaaacaaccaactgaagaagggatctaagccagccaacaaaaatgtggcgactgtc120
85.aagactgtgagccctggaaaattgaagcaattcgtgcaagaaagagatgttaagaaaaaa180
86.acagaacccaaactgactgtgccagtcagaagccttctgacaagagctggagcagcacga240
87.atgaatttggataggaccgaggttctttttcagaacccagagtccttaacttgcaatggg300
88.tttacaatggctctgcgaagcacctctcttagcagacgactctcccaacccccaatggtc360
89.atagccagacccaagaaagttccaccacctaagaatctaggaaagcaacatgagtgtaat420
90.tataaggtacttacagacatgggagtaaagcactcagaaaatgattcagtcccaatgcaa480
91.gaccccccagtcccccctgacatagagaagctaattggcatacaaaatgcatctttcatt540
92.aaagacgagagccaagagacaacccagtcttggtcccaaggtgtccaggattccaaaata600
93.atttgctctactcctagtgaccctgcagcagagattctttctgggtcactggaagggaca660
94.tgtggtggagaaggactattctctagagagacattgaataatatcagtgattcccctaga720
95.atgtttgctcaggacactgtctgtgttcctttgctccaaagagcagccctcaaggttacc780
96.tctgaaggaaaccccagcattcagttagaagatttgggttcacgagtagaatctcttaag840
97.ctgtctgattcttacctggatcccatcaaaagtgaacatgattgctgccccacctccagc900
98.tttaataaggttgtacctgcgttggaccttagaaactgtttgtctattggtgggtcgata960
99.tatcctacctcattaataaaactcctcttggcaggttcagaacaagaaacccttggtgct1020
100.aaaccagatcatcaagaggtactcagagctaccccagatcaacaggaagttcctgatacc1080
101.acccctgtcctaggacaggcctttagtgctgtcccacatcaatgggaagttcctggtgct1140
102.aacccagttcatggagagcctttgggtgagactccagacccaccagagattcctggtgct1200
103.attccagtccaaggagaggtctttggtgctatcttaaatcaacaagtccttggtatgggt1260
104.ggtggtactgcctcagacccacctatttttcttcctgcccctcttaacccagttgctacc1320
105.tgtaatgctctaccagcatggcccgagccccagggcactgtttcatctgggcttgaggtc1380
106.ccgggcactatgcagattttgcctttgggttcaggtcacactcctcaaccatcatcaaac1440
107.tcagagataattttggtacctcctgtaatacctatgaacaatatagaaaatgagaagaag1500
108.gttcatataagctttctgccagctcacactcaggggttcacattagcccctgagagagga1560
109.cctcagccggctccacagggcattgcccagctccccctggtgagtcccagcacattggaa1620
110.ggagggagctcccagatcagcgccaccaggttggttgacatcaacaccacagtggtgtct1680
111.aggccagtgccacttgctagtacctcttctccttcctacacaactctgctacctaccttg1740
112.gaaaagaagaaacgaaagcgatgtggggtctgtgagccctgccagcagaagaccaattgt1800
113.ggtgaatgtacttactgtaagaatagaaagaacagccatcagatctgtaagaagaggaaa1860
114.tgtgaggaactgaaaaagaagccatctgtcattgtgtctttggaggttgtaaaggaaaac1920
115.aagaggccccagagggaaaagaagcccaaagttttaaaggcagattttgacaacaaactg1980
116.gtaaatggccccaagtcagaatccatggaatacagtagttgtgctcatgaggaagaacag2040
117.agattggaattgaattcatgtaccttagaaaatgtaactaaaaatgaagaaagtatgaca2100
118.ggtattgaagtggagaattggacacaaaacaagaaatcacacttaactgatcatgtcaaa2160
119.ggagaatttaatacaattaaaacagaagttgaaaatttgaaaaactctgaagatgacagg2220
120.aaaaaaattctacttactgacctccttgaacctcaaaaattgtttgcacaaactgtaaga2280
121.aatggcattaaaaatgtacactatttaccaactgaaacaaatttgtcttttaaaaaaatc2340
122.aacattgaagaacttggcaaggcatttgaaaacaatccttataaattcctgaaagacacc2400
123.gcaaaccataacaatgtcatgagcaccattgcttctagtgcgagctgtgatcatctcaag2460
124.gggaagagtaatgttttggttattcagaagcctggctttaattgtaagtctttcccagat2520
125.cctgtgaactgcaactcaaatagtcatactactatacacaatgaaagtgatcaaccaaaa2580
126.tccctagaaattatagcaaatagagaactaaaagaaggatcaacacttcaaccaagtctc2640
127.ctatccctaatgaaagataggagattaacactagaacaagtagtggccatagaagctctc2700
128.acacaactttcagaagccccatcagaaaattcctctccatcaaagtcagagaaggatgaa2760
129.gaaacagaacagagaacagctagtttgcttagcagctgtaaagctattctgtattctgtg2820
130.agaaaagacttccaagacccaaacttaaaaggggagtcacagagccttcatcactgtccg2880
131.tctttggaaaagcaaagctcgtgcaatgctgtagtgttcaatggccaaaatagcatttct2940
132.aagtcacacaacagctcacctagtgaccaagcatccacaaaggcacaggaaaattcaacc3000
133.gtcacaaattcactttttataccaaattcaaattcatctaaaactgataccaataaaaat3060
134.actgcgcaaggtaggattactcttgacagttctaggaatttgcagcagttgaccacaata3120
135.actagcaaatcagagtattgtaaccagtcactggacaacagcatcaaggtggattcaaag3180
136.gatgattcatcatgtcaagattcagttcacagtaaaatagaagaagatgttgccgcacaa3240
137.ttaacacaacttgcatcaatgattaaattcaattatgtgaaaccagaggacaaaaaagtt3300
138.gaaagtacaccaacaagccttgttgcatataatgtacagcaaaagtataaccaggagatg3360
139.ggcacaccaccacagaaaccaccttccagtgttcaaaataatcaaggttcatcagtaata3420
140.aagcaaaagaacacaatccagaaaaagacaaaatctactccatcaagagatagacggaaa3480
141.aaaaagcacatggtcataagttgtcaagaaaatgaccagaaaaagcaggaacagttgtca3540
142.tatgagtatagtaaattacatgacatttggatagcatctaaatttcaaagatttggtcaa3600
143.tttggtccacatgattttcccattctgttgggaaaaattcctcccctaactaaagtatgg3660
144.aaaccactgactcaaacaagttcagcattagaacacaaaaaattatttcctccacttgct3720
145.cagataaaatttgagagatattatcctgaattagcacaggaaaagatgaaggttgaaccc3780
146.ttggattctatccccataatcccgttaaaaacagaatccactgggcaggcattttcagaa3840
147.aatgcatatactcaggtacagccaacagtgaatgtcaatcagaaagctcatcctttactc3900
148.cagccagcctctccaaccaaccagtgtgctaatgtcctggatggcagtgaccaaagacag3960
149.ttccaagaggatgttaaagaccaactcatgcatcagagactgccaacatttcctggtatc4020
150.tctcatgagaccccgttaccagacccagcacaaattctcaggaacctaaatgtagtatgt4080
151.tcaggtggaattacagtggtttctaccaaaaatgaagaggatgtctgttcatctagtgtt4140
152.ggaacatcagaattttccccaataaacagtgcacagaaaacttttaatgattatgcgatg4200
153.aacttctttaccaatcctacaaaaacccttgtgtccacaacgaaagattctgaattgcct4260
154.acttgcaactgtcttgatcgagttatacaaaaagacaaaggcccatattatacacacctt4320
155.ggggcaggaccaagtgttgctgctgtcagggaaatcatggagaataggtatggtcaaaaa4380
156.ggaaatgcagtaaggatagaaatagtagtgtacacggggaaagaaggaaaaagctctaat4440
157.gggtgtccagttgctaagtgggttttaagaagaagcagtgacgaagagaaagttctttgt4500
158.ctggttcgacagcgtacaggccaccactgcccaacagctgtgatggtggtgctcatcatg4560
159.gtatgggacggcattcctctcccgatggctgacaaattgtactcacagcttaccgagagt4620
160.ctcaagtcatacaatggtcatcccacagaccgaagatgcacgctcaatgaaaatcgtacc4680
161.tgtacatgtcaaggaattgatccagagacttgtggagcttccttctcttttggctgttca4740
162.tggagtatgtatttcaatggctgcaaatttggtagaagcccaagtcctagaagatttaga4800
163.attgatccaagttctcccttacatgaaaaaaaccttgaagataatttacagagtttggct4860
164.acagaattagctccaatttataagcagtatgctccagctgcttatcaaaaccaggtggcg4920
165.cttgaacatattgcccgagaatgtcggcttgggaagaaagaaggtcgtcctttctctggg4980
166.gtcactgcttgcctagacttctgtgcccatccccacagggacattcacaacatgaataat5040
167.gggagcacagtggtctgcactttaacacgagaagataaccgctcatttggtgttattcct5100
168.caagatgagcagctgcatgtgcttcctctttataaactttcagacacagatgagtttgga5160
169.tcaagggaaggaatggaagccaagatcaaatctggggccatcatggtccttgcaccccgc5220
170.cagagaaaaagaatgcgatttactcaacctgttcctcgttctgggaagaagagagcagcg5280
171.atgatgacagaggttctggcacataagataagggctgtggaaaagaaatttattcctcga5340
172.atcaagcggaagaataattcaatggcaaacaatagtaaggcttcatcactgcccctttta5400
173.ggaaataaaacagagcctgcgcaacctgaaataaaaagtgaaactgaaccccatcttatc5460
174.ttcaaaggttcggacaacactaaaagctactcaccaaccccgtccattcctcacccaggg5520
175.aaagaggcaaacttcgctccagccttctcctggtcctctaagactgctttggtcacaaca5580
176.gctccttttaagaatgatggatcactttcgtacgggttttcagaaagaggaagcaatccc5640
177.cactgcgtaatgccttctgcgagacacagtggtgctaatgtagctgctggggaatgcgct5700
178.gaaattgcacagcctagtgaagtggttcctcttcccaccttgtctactcctatgacaaat5760
179.tccctgggttattctgagcctcccattggcccatctgagcagctaacttccaatcagcca5820
180.aaccagcagcctccattcattacttctcctcatgagcttgcttcctctctggtggaagaa5880
181.gacgagcagccttctgaagcagatgagcctccatcagacgaccccctgtcagacgacccc5940
182.ttgtcacctgctgaggataaactgccccacattgttgagtattggtcagacagtgagcac6000
183.atcttcctggatgccaatgttggcggcgtggctatagcgccttcccacggctcggttctg6060
184.atcgagtgcgcccggcgggagcttcacgccacgactcctgttgaacaccccaaccgaaat6120
185.caccctacgcgcctctctcttgtcttctaccagcacaaaaacctgaataagccccaacac6180
186.ggtttcgaactaaacaagattaaatttgaggcaaaagaagctaagaataagaaaactaaa6240
187.gcctccgaacagaaagaccaggcagctaacgagggtattgaactgtccacagaagttaat6300
188.gaatttagccagattccttctcacaaagcgttaacattaacccatgacaatgttgtcact6360
189.gtgtccccttatgctctcacacacgttgcggggccctataaccattgggtctga6414
190.tet1的m6a甲基化修饰发生在rna分子腺嘌呤(a)的第六位氮原子上,因此,在不改变tet1氨基酸序列的前提下,将tet1基因中受到m6a修饰的第3999位腺苷酸(a)同义突变为鸟嘌呤(g),以达到降低tet1mrna的m6a甲基化修饰水平的目的(图2);
191.实施例3:tet1mrnam6a甲基化位点突变后对tet1mrna稳定性的影响
192.(1)分别将牛tet1基因((ensbtat00000086206.1)编码区序列和同义点突变后的序列克隆到pcdna3.1-3xflag-egfp-c哺乳动物表达载体中,获得野生型(mef2c-wt)和突变型(mef2c-mut)表达质粒;
193.(2)待秦川牛骨骼肌成肌细胞生长至80~90%密度时,将细胞传代接种至6孔板;
194.(3)成肌细胞生长至80%密度时,使用lipofectamine3000试剂盒(invitrogen公司)将mef2c-wt和mef2c-mut表达质粒转染进成肌细胞,主要参照lipofectamine3000试剂盒说明书进行,具体操作如下:
195.1)每孔细胞,在120μl的opti-memi培养基中加入5μl的p3000试剂和2500ng质粒dna;
196.2)每孔细胞,在120μlopti-memi培养基中加入5μllipofectamine3000试剂;
197.3)分别混合均匀后,放置5分钟;
198.4)将上述两种稀释后的dna和lipofectamine3000试剂混合,轻揉摇匀,放置10-15分钟;
199.5)直接将混合物滴加到每孔细胞的培养液中,轻轻摇动混匀,每次实验设置3个生物学重复孔;
200.(4)转染后,继续在37℃、5%co2的培养箱中培养6-8小时后,更换不含抗生素的完全培养基;
201.(5)转染24小时后,在细胞培养液中加入5μg/ml的放线菌素d(抑制mrna转录),在处理后0、3、6、9个小时收集细胞,提取细胞总rna,进行反转录,利用rt-qpcr技术检测tet1的mrna表达量,比较tet1-wt组和tet1-mut组中tet1基因在放线菌素d处理后的表达情况,计算tet1mrna的稳定性;
202.结果如图3所示,tet1-wt组中tet1基因的mrna表达水平下降更快,mrna稳定性明显降低,表明m6a甲基化位点降低tet1mrna的稳定性,促进其mrna降解。
203.实施例4:dna去甲基化酶tet1受m6a甲基化修饰调控,tet1 mrna的m6a甲基化修饰被m6a甲基转移酶mettl3和m6a阅读蛋白ythdf2结合;
204.(1)分别将tet1的编码区野生型序列和m6a位点突变序列克隆进带有renilla荧光素酶和firefly荧光素酶的psicheck2双荧光素酶载体中,构建psicheck2-mef2c-wt和psicheck2-mef2c-mut质粒;
205.(2)待293a细胞生长至90%时,分别将上述双荧光素酶质粒和mettl3的干扰rna或表达质粒共转染进293a细胞中,转染操作同实施例3;
206.(3)转染48小时后,使用双荧光素酶报告系统试剂盒(promega公司)检测双荧光素酶活(图4);
207.(4)m6a甲基化修饰的作用一般是用过m6a阅读蛋白来表征,已证明m6a甲基化位点抑制了tet1 mrna的稳定性(图3),因此猜测m6a阅读蛋白ythdf2结合tet1 mrna的m6a修饰位点,发挥促mrna降解的作用;
208.(5)同样地,利用双荧光素酶报告系统检测转染ythdf2的干扰rna或过表达质粒后的双荧光素酶活(图4);
209.(6)使用rna结合蛋白免疫沉淀(rip)实验进一步检测ythdf2与tet1 mrna的结合(图4),具体步骤参照magna rip
tm
rna-binding protein immunoprecipitation试剂盒(17-700,sigma-aldrich公司)的说明书进行。
210.结果如图4所示,在野生型tet1双荧光素酶报告质粒存在的情况下,mettl3敲除和过表达分别导致荧光素酶活性升高和降低,而并没有影响m6a突变后的荧光素酶活性,表明mettl3与tet1 mrna结合并调控tet1 mrna的m6a修饰水平;相似地,在野生型mef2c双荧光素酶报告质粒存在的情况下,ythdf2的敲除和过表达分别促进和降低了荧光素酶的活性,而并没有影响突变型tet1双荧光素酶报告质粒组中的荧光素酶活性,表明ythdf2与tet1 mrna的m6a位点的结合;rip实验进一步证实了m6a阅读蛋白ythdf2与tet1 mrna的m6a甲基化位点的结合,进而发挥促进tet1 mrna降解的作用。
211.实施例5:tet1 mrna m6a点突变后对tet1表达的影响
212.(1)分别将tet1-wt和tet1-mut表达质粒转染进牛骨骼肌成肌细胞,操作步骤同实施例3;
213.(2)转染24小时后,更换为分化培养基,诱导细胞成肌分化;
214.(3)分化第3天,收集细胞,提取rna和蛋白质,通过rt-qpcr和western blot检测tet1基因的mrna和蛋白表达情况(图5)。
215.结果如图5所示,tet1的m6a位点突变后较野生型明显促进了tet1的mrna和蛋白表达,表明m6a甲基化修饰抑制tet1的表达。
216.实施例6:tet1 mrna m6a甲基化位点对骨骼肌分化的影响
217.(1)分别将野生型(tet1-wt)和突变型(tet1-mut)表达质粒转染进成肌细胞,操作步骤同实施例3;
218.(2)在分化第3天,收集细胞,进行myhc蛋白的免疫荧光试验检测肌管的生成(图6);
219.(3)同时收集分化第3天的细胞,提取总rna、反转录、rt-qpcr检测相关基因的表达(图6)。
220.结果如图6所示,与对照组和野生型组相比,tet1的m6a位点突变后更加增强了成肌分化,促进了肌管生成和融合指数,同时相对于tet1-wt组,tet1-mut显著促进了骨骼肌生成特异性基因myog、myh3和mymk的表达。
221.实施例7:tet1介导mettl3启动子的dna去甲基化促进mettl3的表达进而调控m6a甲基化水平
222.(1)实施例6中的结果也发现了tet1过表达促进mettl3的表达(图7);tet1作为一个dna去甲基化酶,能够通过将靶基因dna的5-胞嘧啶甲基化(5mc)转变成5-胞嘧啶羟甲基化(5hmc),从而起到dna去甲基化的作用。因此,猜测tet1也可以介导mettl3的dna甲基化水平。
223.(2)在甲基化岛预测和引物设计网站methprimer(http://www.urogene.org/cgi-bin/methprimer2)预测并设计了mettl3启动子上cpg的特异性亚硫酸氢盐测序pcr(bsp)引物:
224.mettl3-bsp-f:5
’‑
ttttgggagttggtgatggatagatag-3’225.mettl3-bsp-r:5
’‑
atcccaacaaatactataaaaaaaa-3’226.(3)收集对照和tet1敲低后分化第3天的成肌细胞样品,提取dna,使用ez dna甲基化试剂盒(zymo research公司)将dna进行亚硫酸氢盐处理,作为pcr反应的模板;
227.(4)使用takara epitaq hs试剂盒(takara公司)进行pcr扩增反应;
228.(5)获得的pcr产物纯化后,连接pmd 19-t载体,挑取阳性单克隆进行测序分析;
229.(6)将测序结果在甲基化分析网站quma(http://quma.cdb.riken.jp)进行可视化,并计算dna甲基化(5mc)水平(图7)。
230.结果如图7所示,tet1-wt促进mettl3的表达,且m6a甲基化位同义突变后的tet1-mut组更加提高了mettl3的表达;同时,tet1敲低增加了mettl3启动子的dna甲基化水平;表明dna甲基化调控mettl3的表达进而调控m6a甲基化水平。
231.最后,需要说明的是,以上列举的若干个具体实施例是较优的例子,本发明不限于这些实施例,本领域技术人员可以对实施例中所示的技术方案做出等效变化或添加,应属于本发明技术方案限定的范围。
232.233.234.235.236.
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1