一种发酵生产长春质碱的基因工程菌及其应用
1.本发明涉及生物工程技术领域,具体涉及一种发酵生产长春质碱的基因工程菌及其应用。
背景技术:
2.长春花(catharanthus roseus)是夹竹桃科(apocynaceae)植物,它含有100多种蚓噪生物碱,多数具有生物活性。其中长春碱(vin-blastine)和长春新碱(vincristine)可用于治疗何杰金氏病,恶性淋巴肿瘤、急性淋巴细胞型白血病、绒毛上皮细胞癌及其它癌症,其硫酸盐已经广泛应用于临床,是目前应用最广的天然植物抗肿瘤药物。但是长春碱和长春新碱在植物体中的含量极其微小,仅为百万分之几至十万分之几,化学合成和半合成也不太理想。所以长春碱和长春新碱供应紧张,因而寻找长春花生物碱的生物合成途径具很大的可能性,以及其催化酶乃至基因的表达与调控也是改观长春碱生产量的一种途径。由于长春花生物碱包含两种生物碱即蚓生物碱和单帖生物碱合成途径的调控,所以它的生物合成途径已经作为一个研究植物次生代谢的模式。
3.单帖吲哚生物碱是一大类具有重要药用价值的植物天然产物,包含抗癌药物,抗血管疾病药物,抗疟疾药物等。其中,从长春花中提取的长春碱是最有效的治疗癌症的药物之一。长春碱是长春质碱和文多灵的异质二聚体,其在植物中的含量极低。由于结构复杂,化学全合成存在环境污染,收率低,成本高等问题。随着合成生物学的发展,利用微生物尤其是酵母合成这类高价值化合物的越来越具备可行性。
4.毕赤酵母(pichia pastoris)被认为是基础生物学研究中的重要模型,它能在以甲醇为唯一碳源和能源的培养基中生长,具有蛋白高表达的特性,已被认为是最具发展前景的生产蛋白质的宿主之一。毕赤酵母对重组蛋白的加工水平与哺乳动物类似,甚至部分人源蛋白可以在毕赤酵母中找到同源的蛋白。毕赤酵母不仅拥有已知最强的真核启动子paox1,还可以进行高密度发酵,有利于工业规模化的生产。毕赤酵母基因冗余较小,在利用甲醇为唯一碳源时,甲醇诱导型启动子可以高水平的表达外源蛋白,而将内源蛋白控制在较低表达水平。基于以上优点毕赤酵母表达系统得到了广泛的应用,已成功表达了酶、疫苗、细胞毒素等1000多种外源蛋白。
5.毕赤酵母相比于其他酵母,表达的p450酶活性更高,是一个潜在的能够高效合成天然产物的细胞工厂。与外源蛋白表达相比,天然产物的生物合成是一个相当复杂的过程,不仅包含多步骤的生物合成途径,还需要对整个代谢网络进行协调控制。然而,毕赤酵母常用的整合位点只有少数几个,缺乏足够的中性位点用于外源基因整合。同时在毕赤酵母中重构多基因外源途径的研究较少,也是重构长春质碱合成途径需要克服的难点。
6.工业化生产长春花生物碱不仅靠各种工程技术的发展和培养条件的优化,也要靠生物碱合成规律的理论指导。只有了解了长春花生物碱的合成途径和一些关键限速酶的调节规律,乃至它们的基因的表达调控,才可能用细胞工程或基因工程手段生产长春花生物碱。
技术实现要素:
7.本发明利用已经表征过的整合位点,基于模块化的合成生物学策略,在毕赤酵母中重构长春碱单体(长春质碱)的合成途径。实现了从简单碳源甲醇出发,以环保无污染的方式合成了单萜吲哚生物碱,提供了一种发酵生产长春质碱的基因工程菌及应用。
8.本发明构建了一株发酵生产长春质碱的酵母工程菌,以毕赤酵母作为出发菌株,通过导入外源途径,敲除歧化途径,解除限速步骤,优化发酵条件等方法,使得长春质碱的产量得到了显著的提升。首先,根据已有的文献确认了长春质碱的合成途径,对已经报道的途径和改造步骤进行总结对比,设计最适的构建方法。考虑到途径过长(图1),不利于整体探充。因此以相对稳定的中间产物作为关键节点,进行分模块(can模块、str模块、nep模块)构建。进一步地,对途径中的限速步骤进行了分模块的优化,通过增加拷贝数的方式,提高了每个模块的转化率。进一步地,通过代谢工程的方法来增加辅因子供应,增强p450酶活性,减少外源蛋白的错误折叠等,从而显著提高长春质碱的产量。
9.本发明提供了一种发酵生产长春质碱的基因工程菌,以酵母作为出发菌株,通过基因改造构建获得所述基因工程菌,所述基因改造方式为以下任意一种:
10.(1)整合can模块;
11.(2)整合can模块和str模块;
12.(3)整合can模块、str模块和nep模块;
13.其中,can模块中导入的外源基因包括:异胡豆苷-β-d-葡萄糖苷酶sgd的编码基因、缝籽木榛合成酶gs的编码基因、缝籽木榛氧化酶go的编码基因、氧化还原酶1 redox1的编码基因、氧化还原酶2redox2的编码基因、花冠木碱-o-酰基转移酶sat的编码基因、前二酮乙酸酯合成酶pas的编码基因、dihydroprecondylocarpine合成酶dpas的编码基因、长春胺合成酶cs的编码基因;
14.str模块中导入的外源基因包括:开环马钱子苷合成酶sls的编码基因、异胡豆苷合酶str的编码基因、马钱子酸甲基转移酶lamt的编码基因、7-脱氧番木鳖酸糖基转移酶7dlgt的编码基因、7-脱氧马钱子酸羟化酶7dlh的编码基因、琉乙二醛氧化酶io的编码基因、色氨酸脱羧酶tdc的编码基因、醇脱氢酶cypadh2的编码基因;
15.nep模块中导入的外源基因包括:香叶醇合酶ges的编码基因、法尼基焦磷酸合成酶突变体erg20
ww
的编码基因、香叶醇氧化酶g8h的编码基因、8-羟基香叶醇还原酶gor的编码基因、琉蚁二醛合酶isy的编码基因、乳胶类蛋白mlpla的编码基因、碳源响应转录因子mxr1的编码基因、异戊烯基焦磷酸异构酶idi1的编码基因、截短的羟甲基戊二酰辅酶a还原酶thmgr的编码基因、特异性牻牛儿焦磷酸合酶aggpps2的编码基因;nep模块中敲除的基因包括:老黄酶oye3a的编码基因、老黄酶oye3b的编码基因。
16.优选的,香叶醇合酶ges的编码基因为n端截短35个氨基酸的香叶醇合酶ges截短体的编码基因,所述香叶醇合酶ges截短体的编码基因的genbank号为afd64744。前二酮乙酸酯合成酶pas的n端与麦芽糖结合蛋白mbp的c端通过3个ggggs序列连接,命名为mbp3-pas,进行融合表达,所述麦芽糖结合蛋白mbp的氨基酸序列的genbank号为3q26_a,mbp3-pas的基因序列如seq id no.2所示。
17.(a)整合can模块时:增加1个单拷贝的异胡豆苷-β-d-葡萄糖苷酶sgd的编码基因、缝籽木榛合成酶gs的编码基因、mbp3-pas的编码基因、dihydroprecondylocarpine合成酶
dpas的编码基因、长春胺合成酶cs的编码基因;
18.(b)整合can模块和str模块时:(a)的基础上,增加1个单拷贝的开环马钱子苷合成酶sls的编码基因、异胡豆苷合酶str的编码基因;
19.(c)整合can模块、str模块和nep模块时:(b)的基础上,增加1个单拷贝的香叶醇氧化酶g8h的编码基因、香叶醇合酶ges的编码基因、法尼基焦磷酸合成酶突变体erg20
ww
的编码基因、乳胶类蛋白mlpla的编码基因、碳源响应转录因子mxr1的编码基因;
20.(d)(c)的基础上,导入的基因为重组酶rad52的编码基因、内质网巯基氧化酶ero1的编码基因、内质网分子伴侣pdi1的编码基因、腺苷甲硫氨酸合酶sam2的编码基因、细胞色素b5的编码基因,敲除的基因为液泡蛋白酶pep4的编码基因。
21.其中,所述老黄酶oye3a的编码基因的genbank号为xm_002492555.1,老黄酶oye3b的编码基因的genbank号为xm_002492556.1,液泡蛋白酶pep4的编码基因的genbank号为8200047,异胡豆苷-β-d-葡萄糖苷酶sgd的编码基因包括来自萝芙木(rauvolfia serpentina)的异胡豆苷-β-d-葡萄糖苷酶sgd rvsgd的编码基因和来自长春花(catharanthus roseus)的异胡豆苷-β-d-葡萄糖苷酶sgd的编码基因,其中,异胡豆苷-β-d-葡萄糖苷酶sgd rvsgd的编码基因的genbank号为aj302044.1,异胡豆苷-β-d-葡萄糖苷酶sgd的编码基因的核苷酸序列如seq id no.1所示;缝籽木榛合成酶gs的编码基因的genbank号为mf770507.1,缝籽木榛氧化酶go的编码基因的genbank号为mf770508.1、氧化还原酶1 redox1的编码基因的genbank号为mf770509.1,氧化还原酶2redox2的编码基因的genbank号为mf770510.1,花冠木碱-o-酰基转移酶sat的编码基因的genbank号为mf770511.1,前二酮乙酸酯合成酶pas的编码基因的genbank号为mh136588.1,dihydroprecondylocarpine合成酶dpas的genbank号为ku865331.1,长春胺合成酶cs的编码基因的genbank号为mf770512.1,开环马钱子苷合成酶sls的编码基因的genbank号为l10081.1,异胡豆苷合酶str的编码基因的核苷酸序列如seq id no.3所示,马钱子酸甲基转移酶lamt的编码基因的genbank号为eu057974.1,7-脱氧番木鳖酸糖基转移酶7dlgt的编码基因的genbank号为kf302068,7-脱氧马钱子酸羟化酶7dlh的编码基因的genbank号为kf302067.1,琉乙二醛氧化酶io的编码基因的genbank号为kf591593.1,色氨酸脱羧酶tdc的编码基因的genbank号为m25151,醇脱氢酶cypadh2的编码基因的genbank号为kp411012,法尼基焦磷酸合成酶突变体erg20
ww
的编码基因的genbank号为853272,香叶醇氧化酶g8h的编码基因的核苷酸序列如seq id no.4所示,8-羟基香叶醇还原酶gor的编码基因的核苷酸序列如seq id no.5所示,琉蚁二醛合酶isy的编码基因的genbank号为jx974564,乳胶类蛋白mlpla的编码基因的genbank号为qke59462.1,碳源响应转录因子mxr1的编码基因的genbank号为xm_002493877,异戊烯基焦磷酸异构酶idi1的编码基因的genbank号为abw98669.1,截短的羟甲基戊二酰辅酶a还原酶thmgr的编码基因的核苷酸序列如seq id no.76所示,特异性牻牛儿焦磷酸合酶aggpps2的编码基因的genbank号为xm_002490515.1,重组酶rad52的编码基因的genbank号为xm_002490995.1,内质网巯基氧化酶ero1的编码基因的genbank号为xm_002489600.1,内质网分子伴侣pdi1的编码基因的genbank号为xm_002494247.1;所述腺苷甲硫氨酸合酶sam2的编码基因来源于酿酒酵母(saccharomyces cerevisiae),genbank号为nm_001180810.3;细胞色素b5的编码基因的genbank号为xm_002493758.1。
22.优选的,在发酵培养时,碳源为甲醇或甲醇与以下至少一种:(1)甘露醇;(2)海藻糖;(3)丙氨酸;(4)山梨醇。
23.本发明还提供了所述发酵生产长春质碱的基因工程菌的构建方法,包括以下任意一种构建方法:
24.(i)使用前体原料异胡豆苷,通过can模块构建基因工程菌can4a;
25.(ii)在(i)的基因工程菌can4a中增加1个单拷贝的异胡豆苷-β-d-葡萄糖苷酶sgd的编码基因、缝籽木榛合成酶gs的编码基因、mbp3-pas的编码基因、dihydroprecondylocarpine合成酶dpas的编码基因、长春胺合成酶cs的编码基因和异胡豆苷-β-d-葡萄糖苷酶sgd rvsgd的编码基因序列,获得基因工程菌can7;
26.(iii)将前体原料异胡豆苷替换为荆芥内酯,在(ii)的基因工程菌can7的基础上,通过str模块构建基因工程菌can9;
27.(ⅳ)在(iii)的基因工程菌can9中增加1个单拷贝的开环马钱子苷合成酶sls的编码基因和异胡豆苷合酶str的编码基因序列,获得基因工程菌can11;
28.(v)不使用前体原料荆芥内酯,在(ⅳ)的基因工程菌can11的基础上,通过nep模块构建基因工程菌can14;
29.(vi)在(v)的基因工程菌can14上增加1个单拷贝的香叶醇氧化酶g8h的编码基因、香叶醇合酶ges的编码基因、法尼基焦磷酸合成酶突变体erg20ww的编码基因、乳胶类蛋白mlpla的编码基因、碳源响应转录因子mxr1的编码基因、香叶醇合酶ges的编码基因和法尼基焦磷酸合成酶突变体erg20ww的编码基因序列,获得基因工程菌can17;
30.(vii)在中将重组酶rad52的编码基因、细胞色素b5的编码基因、内质网巯基氧化酶ero1的编码基因和腺苷甲硫氨酸合酶sam2的编码基因序列整合到基因工程菌can17,并敲除液泡蛋白酶pep4的编码基因,获得基因工程菌can20。
31.优选的,发酵培养基因工程菌时,碳源为甲醇或甲醇与以下至少一种:(1)甘露醇;(2)海藻糖;(3)丙氨酸;(4)山梨醇。
32.本发明还提供了所述发酵生产长春质碱的基因工程菌在制备长春质碱中的应用。
33.本发明还提供了一种制备长春质碱的方法,发酵培养所述发酵生产长春质碱的基因工程菌,并提取获得长春质碱;
34.其中,(1)培养发酵通过can模块基因改造方式构建获得的所述基因工程菌时,增加前体异胡豆苷;
35.(2)培养发酵通过can模块和str模块基因改造方式构建获得的所述基因工程菌时,增加前体荆芥内酯
36.(3)培养发酵通过can模块、str模块和nep模块基因改造方式构建获得的所述基因工程菌时,仅使用碳源;所述碳源为甲醇或甲醇与以下至少一种:(1)甘露醇;(2)海藻糖;(3)丙氨酸;(4)山梨醇。
37.本发明实现利用微生物以简单碳源为底物发酵生产长春质碱。实现了从头合成长春质碱的关键技术难点,强化毕赤酵母中心代谢,为萜类吲哚生物碱的生物合成提供参考。本发明利用微生物一步法合成长春质碱,与植物提取和化学合成相比,不受气候地理等条件限制,环境友好,满足国家绿色生物制造的战略需求。本发明的酵母工程菌通过代谢甲醇直接合成长春质碱,其产量可达2.57mg/l,为其工业化生产奠定了基础。
附图说明
38.图1为长春质碱的生物合成途径图。
39.图2为can模块的产量优化图。
40.图3为str模块的产量优化图。
41.图4为nep模块的产量优化图。
42.图5为代谢工程优化对长春质碱产量的变化图。
43.图6为菌株can20采用甲醇(a)或混合碳源(b)进行上罐发酵的曲线图。
具体实施方式
44.毕赤酵母pichiapastoris gs115和大肠杆菌escherichia coli dh5α购买自中国工业微生物菌种保藏中心(cicc)。dna polymerase、限制性内切酶与t4连接酶购买自neb公司。质粒提取试剂盒购买自axygen有限公司,pcr产物核酸纯化试剂盒购买自thermo scientific公司。
45.大肠杆菌的培养使用的lb培养基配方为:胰化蛋白胨10g/l、酵母提取物5g/l、nacl10g/l。
46.毕赤酵母培养和发酵使用的ypd(或ypm或ypman)培养基的配方:酵母提取物10g/l,蛋白胨20g/l,葡萄糖20g/l(或甲醇20ml/l或甘露醇20g/l)。
47.抗生素的配方:博来霉素的配置浓度为0.1g/ml,使用浓度为0.1g/l。g418的配置浓度为0.2g/ml,使用浓度为0.2g/l。潮霉素的配置浓度为0.2g/ml,使用浓度为0.2g/l。
48.毕赤酵母电转化方法:
49.(1)取新鲜的单菌落接种于5mlypad试管,30℃,250rpm培养过夜。转接到50ml的ypad摇瓶中,初始od
600
在0.15-0.20范围内;
50.(2)摇瓶培养4-5h,使od
600
到达0.8-1.0之间(酵母传一代的时间在100-120min);
51.(3)培养液转移到50ml离心管中,5000rpm离心5min。然后弃去上清;
52.(4)用8.5ml预冷的beds溶液(10mm bicine-naoh,ph8.3;3%ethylene glycol(v/v),1m sorbitol),450μl dmso和1ml dithiothreitol(dtt,1.0m)将沉淀重悬。重悬液置于30℃,100rpm的环境中反应5min;
53.(5)5000rpm离心5min,然后弃去上清。用0.95ml beds溶液和50μl dmso重悬沉淀;
54.(6)5000rpm离心30s,然后弃去上清;用0.19ml beds溶液和10μl dmso重悬沉淀;感受态细胞至此制备完成(将细胞放入聚苯乙烯泡沫盒中,在-80℃的温度下将细胞缓慢冷冻。在这种温度下,感受态细胞至少可以保存6个月);
55.(7)取40μl感受态细胞,添加1000ng donordna,500ng质粒dna,混合均匀。转移到预冷的电转杯中;
56.(8)电转的参数如下:gene ii electroporator(bio-rad laboratories,hercules,美国加州),试管间隙2.0mm;充电电压1500v;抵抗200ω;电容25μf;
57.(9)电转后,加入1ml山梨醇和ypad按1∶1混匀的混合液,30℃,250rpm孵育2-4h,取200μl涂布于选择性培养基平板。
58.实施例1:毕赤酵母高效整合位点
59.1、毕赤酵母整合位点:
60.基因组上两个表达框中间的区域作为待测整合位点。以整合效率、稳定性和表达强度作为筛选条件,得到了一系列具有高整合效率或高表达强度的整合位点(表1),并根据整合位点序列构建了一系列grna质粒。结合crispr/cas9基因组编辑技术,这些整合位点可用于外源基因或途径的高效导入,根据不同需求可选用不同的位点。
61.表1整合位点
62.整合位点上游基因下游基因sgrna对应结合到酵母基因组的靶点序列int2pas_chr1-1_0053pas_chr1-1_0054ccgtcactatgaggacaaagint3pas_chr1-4_0182pas_chr1-4_0183tgtggttaactgactaacagint4pas_chr1-4_0202pas_chr1-4_0203gaaatgttgctcggtgcgcgint5pas_chr1-4_0248pas_chr1-4_0249gaaaaatggtccaaattgtginta6pas_chr1-4_0276pas_chr1-4_0277tagaatgcaactgggcaccaint17pas_chr2-1_0727pas_chr2-1_0728agacacagaagagaactgggint18pas_chr2-1_0797pas_chr2-1_0798agtcaatcaggatggtgtggint19pas_chr2-2_0086pas_chr2-2_0087tcgaattttactcgcaatggint20pas_chr2-2_0412pas_chr2-2_0413gtgacgaaagagatgaggtgint21pas_chr2-2_0498pas_chr2-2_0420aattacttcgggaataatggint22pas_chr3_0053pas_chr3_0054gatcagttcattgatagacaint23pas_chr3_0267pas_chr3_0268ttatgggtgttatctctttgint25pas_chr3_1114pas_chr3_1115actgatttgccagaagaacaint26pas_chr3_1202pas_chr3_0618ggggtttgaataacagacacint31pas_fragb_0066pas_fragb_0067tatctgaagtatttactgggint32pas_chr1-3_0003pas_chr1-3_0004ctacagataccattgaatgtint33pas_chr2-1_0010pas_chr2-1_0011tatctgtgtaaagcgacgagint34pas_chr2-1_0528pas_chr2-1_0530tgaggaggaaacctccaccgint35pas_chr3_1156pas_chr3_0154tattaaaaaagacgatcccgint36pas_chr4_0465pas_chr4_0467agaagaaaatgcgaaacaggint37pas_chr4_0575pas_chr4_0576gcaccatctggatagcattg
63.实施例2:产长春质碱的菌株构建及优化
64.利用基因编辑工具和代谢工程手段,在毕赤酵母中实现了长春质碱的全合成(图1)。合成途径中涉及的植物或微生物来源的酶,大部分可以通过美国国家生物信息中心ncbi(https://www.ncbi.nlm.nih.gov/)找到对应的氨基酸序列或基因序列。以异胡豆苷和荆芥内酯为中间产物,将长春质碱的合成途径分成can、str和nep三个模块,进行分步构建与优化。以甲醇为碳源,结合代谢工程的手段,有效的提升了长春质碱的产量。
65.1、can模块的构建及优化
66.(a)can模块的构建
67.can模块包含异胡豆苷到长春质碱的生物合成途径。利用构建的基因编辑工具,在菌株gs115-cas9(由毕赤酵母gs115菌株基因组在his4位点引入cas9蛋白编码基因后培养得到,his4位点为毕赤酵母gs115常用的外源基因整合位点)中重构can模块,利用40bp同源臂的扩增目的片段的表达盒(详见表2)。构建的含有目的基因表达盒的载体,可作为donor
的模板(表3)。将donor和相应的grna一起导入到毕赤酵母细胞中,孵育4h后涂布于带有相应筛选抗性的ypad平板上。
68.从筛选平板上获得的转化子,可进行破胞pcr验证。采用基因组上的引物(表2)扩增,如果大小正确,可进一步测序验证。若测序正确,可进行下一轮转化。3轮转化后,可得到菌株can4a。
69.将验证正确的菌株can4a接种于5mlypam培养24h(30℃,220rpm),然后转接到新鲜的5mlypam培养16h。将菌液转接到50mlypam中培养3天(25℃,220rpm),每24h补充2%的甲醇(v/v)和1.5mg/l的异胡豆苷。发酵液取上清进行lc-ms分析,得到长春质碱的产量为125μg/l。
70.长春质碱的分析方法为:取50ml菌液4000rpm离心5min。将菌液冷冻干燥后,重悬于1.5ml 20mm的tris-hcl(ph 8.0)中。用1ml的乙酸乙酯进行萃取,将萃取所得的上层液体用0.22μm的有机系滤头过滤。有机相采用agilent 1290-6470uhplc-msms分析,通过zorbax eclipse plus c18 hplc(2.1
×
50mm,1.8-micro)柱和esi离子源对长春质碱进行鉴定和定量。
71.用lc-ms多级反应监测模式(mrm)监测长春质碱,m/z从337.1跃迁到144.0,碰撞能量为22ev,m/z从337.1跃迁到93.1,碰撞能量为30ev。脱溶线(dl)温度为300,喷雾电压为4kv,雾化气体流量为3l/min。流动相a为0.1%甲酸溶液,流动相b为乙腈。洗脱程序:流动相a从95%降到20%,整个下降过程保持在10min。然后从20%上升到95%,整个上升过程保持在10min。流动相流速保持不变在0.3ml/min。
72.表2基因组整合can模块基因所用的引物
73.引物名称引物序列(5’到3’)str-gs-fccatacttcttttgcaacgtaaatactagtctaaaagcttttttttcagaccatatgacstr-gs-rtcaggccttaaaggaaatttggaactccagtgccactttcatatggtgttatgtacgcaredox-sat-fatcaagaaaaacatcccccgtggtcccttccaaagactctcactgcaggaatctctggcredox-sat-raggaatgtcaggagattgaatgactgatgacgcaagttctgctagggttgttccgttagcs-dpas-faacaacttgccaaaacattctgacttctttcatgtgtcactgcaggaatctctggcacgcs-dpas-raaacgcactgaaaatcggcactacttacataaaacccagcgcaaattaaagccttcgaggo-fgagccggaacaaaaaatttgcttcaccacccattacaagacagatctaacatccaaagago-rtcaaacttgtcacttttgacggtgacgtaacattgtacgagcgcaaattaaagccttcgcpr-fagaaattaaatttggcaattcctctaacatgccatgggatcatgcaggaatctctggcacpr-rtttatgatatgagaagcaaacaagatttatagacagattagcatatggtgttatgtacgpas-ftgggaagttttttatgagacgatgatttcgaaaggttctatttttttcagaccatatgapas-rgaagccctcttggagctgtgttatttactattgaggaagctagggttgttccgttagca
74.表3 can模块构建所用的质粒
75.质粒名称基因型int31-std-gsampr;p2-sgd-p1-gsint26-redox1-redox1-satampr;p3-redox1-p1-redox2-p2-satint37-cs-dpasampr;p3-dpas-p1-csint21-goampr;p1-go
int35-cprampr;p3-cprint36-pasampr;p1-pas
76.注:p1为启动子paox1,p2为启动子paox2突变体(-255bp,t
→
c),p3为启动子pfld1。
77.(b)can模块的优化
78.为了优化can模块,对其中的关键蛋白前二酮乙酸酯合成酶pas(genbank号为mh136588.1)的可溶表达水平进行优化(表4),发现与mbp(mbp为麦芽糖结合蛋白,其genbank登录号为3q26_a,此外前二酮乙酸酯合成酶pas的n端与麦芽糖结合蛋白mbp的c端通过3个ggggs序列连接,命名为mbp3-pas,其编码基因序列如seq id no.2所示)融合表达对产量的提升效果最为明显,长春质碱的产量提升了一倍左右。然后用引物对can模块中的每个外源基因增加了拷贝数(表2),得到一系列的菌株。对这些菌株进行发酵,测试其长春质碱的产量(图2)。结果表明,dihydroprecondylocarpine合成酶dpas(编码基因的genbank号为ku865331.1)、长春胺合成酶cs(编码基因的genbank号为mf770512.1)、异胡豆苷-b-d-葡萄糖苷酶sgd rvsgd(编码基因的genbank号为aj302044.1)、mbp3-pas(编码基因序列如seq id no.2所示)、异胡豆苷-β-d-葡萄糖苷酶sgd(基因序列如seq id no.1所示)、缝籽木榛合成酶gs(编码基因的genbank号为mf770507.1)都是该模块的限速步骤。将限速步骤增加一个拷贝到基因组上,得到菌株can7。菌株can7的长春质碱产量为1.1mg/l,为初始菌株can4a的10倍左右。
79.表4含有can模块菌株的优化和产量
80.菌株命名基因修饰前体/碳源长春质碱产量(μg/l)can4a整合can模块异胡豆苷/甲醇125can4embp3-pas融合表达异胡豆苷/甲醇276can5acan4e增加sgd和gs拷贝数异胡豆苷/甲醇459can6can5a增加mbp3-pas拷贝数异胡豆苷/甲醇695can7can6增加dpas、cs和rvsgd拷贝数异胡豆苷/甲醇1125
81.2、str模块的构建及优化
82.(a)str模块的构建
83.str模块包含荆芥内酯到异胡豆苷的生物合成途径。在菌株can7的基础上,利用40bp的同源臂引入str模块只需2轮转化(表5)。同样的,以构建好的菌株作为donor的模板,也可作为增加拷贝数的模板(表6)。得到的菌株can9进行发酵检测,前体替换为50mg/l的荆芥内酯,长春质碱的产量为1.2mg/l(表7)。
84.表5基因组整合str模块基因所用的引物
85.引物名称引物序列(5’到3’)str-lamt-fttgaagtgaaaaaaagaaatgcacgacttatgcaaattagcatttttttcagaccatatstr-lamt-rgtcaaagtacagccctgccagaacacttattgccatctgagcgcaaattaaagccttcgsls-7dlh-factaacccacattttcctcctagcacgtgaatcttttaaacatttttttcagaccatatsls-7dlh-rataatctaactagtttgccgcgaccaccatacaatatttagcatagcaaattaaagcct7dlgt-io-fagatgaagcaaaaatgactgtaatccttatacggctgattcagatctaacatccaaaga7dlgt-io-rgagtgtacatttaatcatgtgttttaaaactgatgatgaagcatatggtgttatgtacg
tdc-cypadh-fatctccacatatcaaggaattatcataaccagtctcttttcagatctaacatccaaagatdc-cypadh-rgtcatcttccttgctttcgtcttattcgatgctactaattgctagggttgttccgttag
86.表6 str模块构建所用的质粒
87.质粒名称基因型int20-str-lamtampr;p2-str-p1-lamtint2-sls-7dlhampr;p2-sls-p1-7dlhint22-7dlgt-ioampr;p1-7dlgt-p3-ioint25-tdc-cypadhampr;p1-tdc-p2-cypadh
88.(b)str模块的优化
89.为了提高str模块对前体的转化效率,利用引物对模块str中涉及的外源基因进行了增加拷贝数(表5),其中,7-脱氧马钱子酸羟化酶7dlh的编码基因的genbank号为kf302067.1,琉乙二醛氧化酶io的编码基因的genbank号为kf591593.1,由此得到了一系列菌株。将这些菌株进行发酵,检测其长春质碱的产量(图3)。可以发现,str模块的主要限速步骤为异胡豆苷合酶str(核苷酸序列如seq id no.3所示)和开环马钱子苷合成酶sls(编码基因的genbank号为l10081.1)。其余基因增加拷贝数,对产量基本没有提升效果。
90.将测定所得的限速步骤增加一个拷贝,得到菌株can11。菌株can11的长春质碱产量为2.7mg/l。
91.表7各菌种发酵生产长春质碱的产量
92.菌株命名基因修饰前体/碳源长春质碱产量(μg/l)can9can7整合str模块荆芥内酯/甲醇1201can11can9增加str和sls拷贝数荆芥内酯/甲醇2667
93.3、nep模块的构建及优化过程如下:
94.(a)nep模块的构建
95.nep模块包含甲醇到荆芥内酯的生物合成途径。利用40bp的同源臂,需要4轮转化可以将nep模块通过相应的表达盒整合到毕赤酵母基因组(表8)。在菌株can11的基础上引入nep模块,并且敲除香叶醇的歧化途径(老黄酶oye3a和老黄酶oye3b,老黄酶oye3a的编码基因的genbank号为xm_002492555.1,老黄酶oye3b的编码基因的genbank号为xm_002492556.1)。根据已有的文献报道,在酵母中重构荆芥内酯合成途径时,香叶醇的歧化途径会严重降低其下一步产物8-羟基香叶醇的产量。为了增强nep模块的荆芥内酯产量,引入组成型启动子驱动的表达盒,产量得到了一定的提升(表9,图4),其中,法尼基焦磷酸合成酶突变体erg20
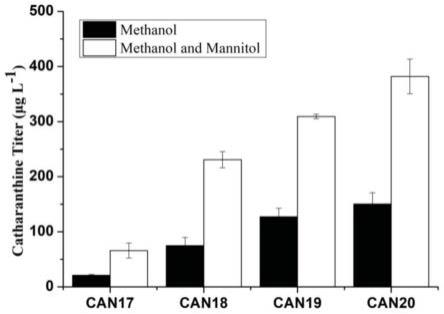
ww
的编码基因的genbank号为853272,香叶醇氧化酶g8h的核苷酸序列如seq id no.4所示,8-羟基香叶醇还原酶gor的核苷酸序列如seq id no.5所示,琉蚁二醛合酶isy的编码基因的genbank号为jx974564,乳胶类蛋白mlpla的编码基因的genbank号为qke59462.1,碳源响应转录因子mxr1的编码基因的genbank号为xm_002493877.1。构建得到的菌株can17在以甲醇为唯一碳源时,产量为20μg/l(表10)。
96.表8基因组整合nep模块基因所用的引物
97.引物名称引物序列(5’到3’)idi-aggpps2-ftcagaatgctgaagtaggcccttacacgtacattagtacaaagtgggttacctttatctidi-aggpps2-rtcagctatgtcgtggaccaacgaaatcttgatacattagcttacagacccaaagcagct
g8h-ges-erg20
ww-fcctcaccaagagtcagatcagcgtaaggaccagttggctcagcacttggttcttcagtcg8h-ges-erg20
ww-rcagcgtgtcaacgctctaggatacaacacattcagcagatgccgaggcaaattaaagccthmg-ftactttaaataagatgactgtgcctgatctgaaagaaacacaaagtgggttacctttatthmg-raccaatcaatgttctatcgtttgcattgacatccctaattgcttacagacccaaagcagisy-fttggtaatcttaataaaaccttctgcttccaaaaacagtttttttttcagaccatatgaisy-rgaggttacaatgtgtattgtatcagttgatttctgggttcatagggttgttccgttagcmxr1-ftcatccgatcattgccctgttgggtagaagagcagaataatgcaggaatctctggcacgmxr1-rtttctcgtgtctctgaatcgcgttttataaaggcgatgtatatggtgttatgtacgcaamlpla-fgctaggttgagatttctcaaccctcatggatcaaatcactttttttcagaccatatgacmlpla-rgagaagaaaaaaaagagggagaggtcggagaaatatacttctagggttgttccgttagcint4-g8h-ges-erg20
ww-fgtcattcaatagggaggtagaagagaactgagggagaaaaagtgggttacctttatctcint4-g8h-ges-erg20
ww-rcaaaaatgtccgagaaattttcagcattagactatccgggaagcagctctcacttaatc
98.表9 nep模块构建所用的质粒
99.质粒名称基因型int17-idi1-aggpps2ampr;p1-idi1-p2-aggpps2int34-g8h-ges-erg20
ww
ampr;p2-g8h-p1-ges-erg20
ww
oye3a-gor-thmg1ampr;p1-gor-p2-thmg1int6-isyampr;p2-isyint33-mxr1ampr;p3-mxr1int4-g8h-ges-erg20
ww
ampr;p12-g8h-p11-ges-erg20
ww
int32-mlplaampr;p2-mlpla
100.注:p11为启动子pgap,p12为启动子ptef1;oye3a-gor-scthmg1指拥有靶向老黄酶oye3a的编码基因的质粒,在敲除老黄酶oye3a基因的同时整合外源基因。
101.(b)菌株培养条件的优化和发酵
102.当甲醇为唯一碳源时,70%以上的甲醛会通过异化途径被消耗,导致合成产物的代谢流不足;而利用葡萄糖或者甘油等碳源会抑制甲醇启动子。因此,选择非抑制性碳源对构建的菌株进行混合发酵测试。从新鲜的ypad平板上挑取单菌落进行培养,接种到含有5mlypad的试管中,在30℃、250rpm条件下培养16h。然后取2.5ml种子液接种到50ml新鲜ypad中培养24h。离心,弃去上清,菌体用无菌水洗2遍后。用ypam和其余碳源(2%的海藻糖,或2%的甘露醇,或2%的丙氨酸,或2%的山梨醇,或2%的葡萄糖)组成的培养基进行重悬,在25℃的环境中诱导培养3天。每24h补充1%甲醇和2%相应的碳源。诱导3天后,收菌,取上清进行lcms检测。结果表明甲醇和非抑制性碳源甘露醇混合使用时,产量提高了3倍以上(表11)。
103.表10各菌种发酵生产长春质碱的产量
104.菌株命名基因修饰前体/碳源长春质碱产量(μg/l)can14can11整合nep模块,敲除oye3a和oye3b无/甲醇2can15bcan14增加g8h和ges-erg20
ww
拷贝数无/甲醇5can16can15b增加mxr1和mlpla拷贝数无/甲醇15can17can16增加ges-erg20
ww
拷贝数无/甲醇20
105.表11菌株发酵的碳源优化
106.菌株命名碳源长春质碱产量(μg/l)can17甲醇20can17甲醇,海藻糖64can17甲醇,甘露醇63can17甲醇,丙氨酸46can17甲醇,山梨醇18can17甲醇,葡萄糖0
107.4、底盘细胞全局优化提高长春质碱产量
108.(a)p450酶表达水平的优化
109.p450往往是天然产物合成途径中的限速步骤。根据已有的研究,发现过表达毕赤酵母的重组酶rad52(genbank号为xm_002490995.1)可以显著提高p450的酶活。而引入细胞色素b5(cyb5)(genbank号为xm_002493758.1),可以提高向p450传递电子的效率,从而提高p450的酶活。因此,在现有的菌株内,过表达重组酶rad52和cyb5的编码基因。设计引物(详见表12),将需要增加拷贝数的基因整合到基因组上,可得到菌株can18。菌株以甲醇为唯一碳源进行发酵,长春质碱的产量可达75μg/l。目的产物的产量提高了3倍以上,说明p450酶是长春质碱合成的主要限速步骤之一。
110.(b)外源蛋白的表达
111.菌株can20包含了数十个拷贝的外源蛋白的高表达,容易引起未折叠蛋白反应(upr),会导致细胞减少翻译,降解未折叠蛋白,从而使得外源蛋白的实际表达量低于理论表达量,导致长春质碱的实际产量远低于理论产量。换而言之,提高外源蛋白的表达量可以有效的提高途径中甲醇的转化效率。因此,敲除液泡蛋白酶pep4(编码基因的ncbi登录号为8200047),减少蛋白的降解。过表达折叠因子内质网分子伴侣pdi1(编码基因的genbank号为xm 002494247.1)和内质网巯基氧化酶ero1(编码基因的genbank号为xm 002489600.1)来协助外源蛋白的折叠。通过敲除蛋白酶,引入折叠辅助因子,可以构建得到菌株can19。该菌株的长春质碱产量为127μg/l。
112.(c)增强辅因子供应
113.腺苷甲硫氨酸是酵母体内一种重要的代谢中间体,可以作为甲基、丙氨基的供体参与胞内各类生化反应。长春质碱的合成途径中,存在多步反应需要甲基作为供体参与(例如lamt催化的反应)。因此,提高甲基的供应量,可以有效的提高途径整体的效率,解除限速步骤。已有研究表明,酿酒酵母的腺苷甲硫氨酸合酶sam2(编码基因的genbank号为nm_001180810.3)效果要优于毕赤酵母的。因此为了增强甲基的供应,过表达酿酒酵母的腺苷甲硫氨酸合酶sam2的编码基因。将腺苷甲硫氨酸合酶sam2的编码基因表达盒整合到基因组上,可得到菌株can20。在以甲醇为唯一碳源的情况下,长春质碱的产量可达150μg/l(表14)。
114.表12底盘细胞全局优化所用到的引物
115.引物名称引物序列(5’到3’)cyb5-fgcccagcaattaccggcttccgtctagcacgtgattgctctgcaggaatctctggcacgcyb5-rgcaattcaagatttacgaggattagcgtgtgagaacttgatatggtgttatgtacgcaarad52-ftagtccagcgatgacttgggaagtccattatcgccaggtggatctaacatccaaagacg
rad52-rattgcggtcctgattcttccaatgcattgattgtttccctgcaaattaaagccttcgagpep4-q1ttgtccaagtcgaattttatgtctcagcaagataaattctcaatgcagaaattggtgcpep4-q2tgtcaaggtagctgacaagcaatcpep4-q3aacccttggtagcaccaatttctgcattgagaatttatcttgctgagacataaaattcgpep4-q4agtagacagaattcataatggtgagpdi1-fcacatccaaaaagatcttattctctccagtacaatcagcactgctactctggtcccaagpdi1-ragtccacgtttcaagatgagggtggttcttgagatcgtctctcacttaatcttctgtacero1-fgtccgccaatcgaaccacaacactattgaagaaagcaaccaaggagatttcagaggagtero1-rgatacgtttttcagtcacaaacacatacacatgcacatgtagggttgttccgttagcacsam2-ftccgccaatcgaaccacaacactattgaagaaagcaacgagatctaacatccaaagacgsam2-rgaggatgatacgtttttcagtcacaaacacatacacatgcgcaaattaaagccttcgag
116.表13底盘细胞全局优化所用的质粒
117.质粒名称基因型int18-rad52-cyb5ampr;p1-pprad52-p2-cyb52int19-pdi1-ero1ampr;p11-pdi1-p12-ero1int23-sam2ampr;p1-sam2
118.表14代谢工程对菌株产量的提升
[0119][0120]
进一步测试菌株can20在混合碳源时的产量,通过发酵得到长春质碱的产量为383μg/l(图5)。
[0121]
为了进一步提高长春质碱的产量,在1l的发酵罐中对菌株can20进行了发酵(图6)。从ypad的平板上接种新鲜的单菌落,接种于5ml ypaman液体培养基中,在30℃、250rpm条件下培养16h。然后取2.5ml种子液接种到50ml新鲜ypaman中培养24小时。将50ml菌液转移到含有0.7l ypaman的1l生物反应器中。在起始温度为30℃、转速为800rpm、空气流速为3l/min的生物反应器中进行发酵。用盐酸氨水维持发酵液的ph在5.5~6.5。在生物反应器中自动添加甘露醇,使溶解氧水平(do)保持在20%~40%。当od
600
达到50时,加入2%的甲醇诱导长春质碱生物合成途径相关基因的表达。诱导4h后,将温度调节到25℃,每3h向生物反应器中加入2%的甲醇或2%的混合液(甲醇∶甘露醇=1∶1)。每3小时取样以确定细胞的生长速度和长春质碱的产量。长春质碱的产量在发酵罐得到了显著的提升,以甲醇作为唯一碳源时的产量可达620μg/l,而混合碳源的产量可达2.57mg/l。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1