一种水稻柱头外露率主效QTL-qTSE8、检测引物、试剂盒和应用的制作方法
一种水稻柱头外露率主效qtl-qtse8、检测引物、试剂盒和应用
技术领域
1.本发明涉及分子生物学技术领域,具体涉及一种水稻柱头外露率主效qtl-qtse8、检测引物、试剂盒和应用。
背景技术:
2.水稻作为最重要的粮食作物之一,养活了全球一半以上的人口,水稻产量的丰欠将直接影响到粮食安全和社会稳定,提高单位面积的粮食产量一直是育种者最重要的目标。杂交水稻的商业化推广极大地促进了粮食产量的增加,其种子生产能力的提高发挥了重要的作用。不育系的异交性能是影响杂交水稻制种产量的关键因素,而柱头外露率则是提高不育系异交性能最为重要的性状。
3.随着基因组学和分子标记的发展,不同的研究者基于连锁分析,利用野生稻和栽培稻或者籼粳杂交构建的作图群体,或者基于关联分析,利用核心种质的自然群体进行了相关的qtl定位研究,定位到超过40个控制柱头外露率的qtl位点在水稻12条染色体上均有分布,定位结果不尽相同,且只有少量qtl解释超过10%的表型变异,表明柱头外露率是受微效多基因控制的数量性状。利用分子标记辅助选择改良水稻柱头外露率性状是一个有效的手段。挖掘稳定的、可靠的控制柱头外露率的qtl,对提高柱头外露率的改良效率起到重要作用,有利于杂交制种产量提高和成本降低,促进杂交水稻的商业化推广应用。
技术实现要素:
4.本发明的目的在于提供一种水稻柱头外露率主效qtl-qtse8、检测引物、试剂盒和应用。利用连锁标记构建含有本发明主效qtl-qtse8的近等基因系柱头外露率显著提高,为分子标记辅助选择改良水稻柱头外露率,提高不育系异交结实率和制种产量提供了参考。
5.本发明提供了一种水稻柱头外露率主效qtl-qtse8,所述主效qtl-qtse8被定位在第8染色体上,位于连锁标记rm38和rm25之间。
6.本发明还提供了一组检测水稻柱头外露率主效qtl-qtse8的引物,所述引物包括rm38的引物和rm25的引物;所述rm38的引物的核苷酸序列如seq id no.1和seq id no.2所示;所述rm25的引物的核苷酸序列如seq id no.3和seq id no.4所示。
7.本发明还提供了一种检测水稻柱头外露率主效qtl-qtse8的试剂盒,所述试剂盒包括上述技术方案所述引物和反应液。
8.本发明还提供了检测上述技术方案所述qtl-qtse8的物质在培育或检测高柱头外露率的水稻品种中的应用。
9.本发明提供了一种水稻柱头外露率主效qtl-qtse8。本发明利用重组自交系的连锁分析和自然群体的关联分析,在第8染色体上重复鉴定到一个稳定影响柱头外露率的主效qtl位点,为进一步精细定位和候选基因克隆提供了可靠依据,利用连锁标记构建含有该主效qtl位点的近等基因系柱头外露率显著提高,为分子标记辅助选择改良水稻柱头外露
率,提高不育系异交结实率和制种产量提供了参考。本发明通过联合连锁与关联分析策略定位到的可靠的主效qtl-qtse8,结果相互验证,置信区间较小,可作为精细定位的重要候选区段。
附图说明
10.图1为本发明提供的重组自交系群体单边柱头外露率定位结果;
11.图2为本发明提供的重组自交系群体双边柱头外露率定位结果;
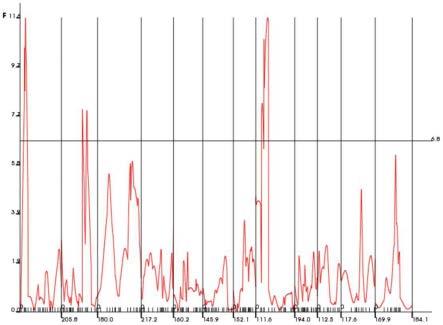
12.图3为本发明提供的重组自交系群体总柱头外露率定位结果;
13.图4为本发明提供的柱头外露率关联分析曼哈顿图。
具体实施方式
14.本发明提供了一种水稻柱头外露率主效qtl-qtse8,所述主效qtl-qtse8被定位在第8染色体上,位于连锁标记rm38和rm25之间,所述rm38的引物的核苷酸序列如seq id no.1(acgagctctcgatcagccta)和seq id no.2(tcggtctccatgtcccac)所示;所述rm25的引物的核苷酸序列如seq id no.3(ggaaagaatgatcttttcatgg)和seq id no.4(ctaccatcaaaaccaatgttc)所示。
15.本发明实施例利用1套重组自交系群体,由珍汕97b和irat109构建,包含163份株系,对两个试验地点的柱头外露率进行了三年的表型调查(包括单边柱头外露率、双边柱头外露率和总柱头外露率)。应用前期构建的遗传图谱,将三年的数据作为环境因子,采取混合线性模型的复合区间作图法进行联合qtl分析。对于单边柱头外露率性状,共检测到3个加性qtl(qsse1、qsse2和qsse8),分别位于1、2和8号染色体上,解释的表型变异为10.3%、4.4%和9.0%;对于双边柱头外露率性状,共检测到2个加性qtl(qdse1和qdse8),分别位于1和8号染色体上,解释的表型变异为4.0%和4.3%;对于总柱头外露率性状,共检测到3个加性qtl(qtse1、qtse2和qtse8),分别位于1、2和8号染色体上,解释的表型变异为9.5%、4.5%和10.0%。
16.本发明还利用1套水稻核心种质群体中的138份籼稻亚群,对两个试验地点的柱头外露率进行了两年的表型调查(包括单边柱头外露率、双边柱头外露率和总柱头外露率)。应用基因组重测序获得的约102万个位点的snp基因型数据,在基因组上覆盖的密度为平均每2.7kb有一个snp。结合snp基因型和表型信息,采取混合线性模型对柱头外露率性状进行全基因组关联分析。对于单边柱头外露率性状,两年在第8染色体上都定位到同一个显著关联位点,2016年海南还在第3染色体上检测到一个显著关联位点;对于双边柱头外露率性状,2016年上海分别在第3、5、7和10染色体上检测到4个显著关联位点,2017年海南一共定位到12个显著关联位点,分布在第1、2、3、4、8、10、11和12染色体上;对于总柱头外露率性状,两年在第3染色体、第8染色体上都定位到同一个显著关联位点,2017年海南,在第9和11染色体上检测到两个显著关联位点。
17.比较两种定位分析结果,发现关联分析在第8染色体上检测到的与柱头外露率显著相关的snp位于rm38~rm25区间内,即通过连锁定位在第8染色体上的检测到的qtl-qtse8区间内,且该区间还未有报道与柱头外露率性状相关,是一个新的位点。利用两种分析方法,在不同环境条件下在第8染色体上重复检测到一个稳定影响柱头外露率的新qtl位
点,为后续研究进行精细定位和候选基因分析提供了可靠依据。
18.本发明还提供了一组检测水稻柱头外露率主效qtl-qtse8的引物,所述引物包括rm38的引物和rm25的引物;所述rm38的引物的核苷酸序列如seq id no.1和seq id no.2所示;所述rm25的引物的核苷酸序列如seq id no.3和seq id no.4所示。
19.本发明还提供了一种检测水稻柱头外露率主效qtl-qtse8的试剂盒,所述试剂盒包括上述技术方案所述引物和反应液。
20.本发明还提供了检测上述技术方案所述qtl-qtse8的物质在培育或检测高柱头外露率的水稻品种中的应用。本发明以珍汕97b为供体亲本,以irat109为受体亲本和轮回亲本,利用qtse8位点连锁标记rm38和rm25每一代选择杂合位点单株与轮回亲本irat109回交,回交4代在自交,在bc4f2群体中,选择rm38和rm25标记检测带型与珍汕97b一致,农艺性状与irat109相似的单株自交,连续自交5代,获得含有qtse8位点的近等基因系,其总柱头外露率与轮回亲本irat109相比显著提高,证明该qtl位点可以提高水稻柱头外露率,对分子标记辅助选择改良水稻柱头外露率具有重要意义。
21.下面结合具体实施例对本发明所述的一种水稻柱头外露率主效qtl-qtse8、检测引物、试剂盒和应用做进一步详细的介绍,本发明的技术方案包括但不限于以下实施例。
22.实施例1
23.1.珍汕97b/irat109重组自交系柱头外露率qtl定位
24.1.1重组自交系群体柱头外露率表现
25.重组自交系群体三次表型调查分别在2016年夏上海、2017年春海南和2018年春海南完成。调查的性状包括单边柱头外露率、双边柱头外露率和总柱头外露率。双亲的柱头外露率表型差异不大,但ril群体变异幅度较大,在海南的柱头外露率均值高于在上海的均值,柱头外露率表现为连续分布,且均观察到有超亲分离现象(表1)。
26.表1珍汕97b/irat109 ril群体及亲本柱头外露率表现
27.[0028][0029]
对2018年海南调查的表型数据相关性分析发现,单边柱头外露率、双边柱头外露率和总柱头外露率呈极显著正相关,其中单边柱头外露率与总柱头外露率的相关系数高达0.987,其次是双边柱头外露率与总柱头外露率的相关系数为0.794,表明单边柱头外露率对总柱头外露率的贡献比较高(表2)。
[0030]
表2 ril群体柱头外露率性状相关分析(2018年海南)
[0031][0032]
**p=0.01
[0033]
采用qtlnetwork 2.0软件利用混合线性模型的复合区间作图(mcim)进行联合qtl分析,环境因子作为变量(表3)。
[0034]
对于单边柱头外露率性状(图1,重组自交系群体单边柱头外露率定位结果),经过置换检验,在5%的显著性水平上,得到加性qtl的f统计量阈值为6.7。共检测到3个加性qtl,分别位于1、2和8号染色体上,解释的表型变异为10.3%、4.4%和9.0%,其中第2染色的qtl的加性效应来自亲本irat109,其余qtl的加性效应来自亲本珍汕97b。
[0035]
对于双边柱头外露率性状(图2,重组自交系群体双边柱头外露率定位结果),经过置换检验,在5%的显著性水平上,得到加性qtl的f统计量阈值为5.9。共检测到2个加性qtl,分别位于1和8号染色体上,解释的表型变异为4.0%和4.3%,加性效应均来自亲本珍汕97b。
[0036]
对于总柱头外露率性状(图3,重组自交系群体总柱头外露率定位结果),经过置换检验,在5%的显著性水平上,得到加性qtl的f统计量阈值为6.8。共检测到3个加性qtl,分别位于1、2和8号染色体上,解释的表型变异为9.5%、4.5%和10.0%,其中第2染色的qtl的加性效应来自亲本irat109,其余qtl的加性效应来自亲本珍汕97b。
[0037]
表3重组自交系柱头外露率qtl定位结果
[0038][0039][0040]
注:a:加性效应,正值表示增效基因来自珍汕97b;ae1,ae2和ae3:分别是2016上
海,2017海南和2018海南试验中加性效应与环境因素的互作效应。
[0041]
2.核心种质群体柱头外露率关联分析
[0042]
2.1核心种质群体柱头外露率表现
[0043]
从270份由中国水稻微核心种质资源和抗旱核心种质资源中选出的138份籼稻亚群作为本试验的关联分析群体,两次表型调查分别于2016年夏季在上海和2017年春季在海南完成。群体柱头外露率变幅较大,总柱头外露率2016年在上海从0.5%到80.5%,2017年在海南也是从0.5%到72.8%,明显要大于重组自交系群体的变幅,说明柱头外露率这个表型性状在本试验所用自然群体中多样性很丰富,遗传多样性较大,适合于进行关联分析(表4)。
[0044]
表4核心种质群体柱头外露率表现
[0045][0046][0047]
2.2gwas分析
[0048]
本试验前期已经通过重测序获得了约102万个位点的snp基因型数据,在基因组上覆盖的密度为平均每2.7kb有一个snp。利用snp分子标记以及表型信息,采取混合线性模型对柱头外露率性状进行全基因组关联定位,以p《10-6
为阈值,结果如下(表5,图4,柱头外露率关联分析曼哈顿图):
[0049]
表5柱头外露率的显著关联位点
[0050]
[0051][0052]
单边柱头外露率
[0053]
两年在第8染色体上都定位到同一个显著关联位点,2016年海南还在第3染色体上检测到一个显著关联位点。
[0054]
双边柱头外露率
[0055]
2016年上海分别在第3、5、7和10染色体上检测到4个显著关联位点,2017年海南一共定位到12个显著关联位点,分布在第1、2、3、4、8、10、11和12染色体上。
[0056]
总柱头外露率
[0057]
两年在第3染色体、第8染色体上都定位到同一个显著关联位点,且这两个位点也影响着单边柱头外露率,表明总柱头外露率与单边柱头外露率显著相关。2017年海南,在第9和11染色体上检测到两个显著关联位点。
[0058]
3 gwas关联定位与连锁qtl定位结果的重合性分析
[0059]
比较核心种质群体关联分析结果和重组自交系连锁定位结果,发现关联分析在第8染色体上检测到的与柱头外露率显著相关的snp位于rm38-rm25区间内,即第8染色体上的通过连锁定位得到的qtl-qtse8。利用两种分析方法,在不同环境条件下在第8染色体上重复检测到一个稳定影响柱头外露率的qtl位点,为后续研究进行精细定位和候选基因分析提供了可靠依据。
[0060]
4利用连锁标记构建qtse8位点的近等基因系
[0061]
以珍汕97b为供体亲本,以irat109为受体亲本和轮回亲本,利用qtse8位点连锁标记rm38和rm25每一代选择杂合位点单株与轮回亲本irat109回交,回交4代在自交,在bc4f2群体中,选择rm38和rm25标记检测带型与珍汕97b一致,农艺性状与irat109相似的单株自交,连续自交5代,获得含有qtse8位点的近等基因系。轮回亲本irat109的单边柱头外露率、双边柱头外露率和总柱头外露率分别为18.6%、0.3%和18.9%,而含有qtse8位点近等基因系的表型则分别为29.4%、9.5%和38.9%,柱头外露率相关性状与轮回亲本相比得到显著提高。
[0062]
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人
员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1