蜡梅CpCBL8基因及其编码的蛋白与应用
蜡梅cpcbl8基因及其编码的蛋白与应用
技术领域
1.本发明属于植物分子生物学领域,具体涉及一个蜡梅cpcbl8基因、其编码的蛋白和应用。
背景技术:
2.蜡梅(chimonanthus praecox)为蜡梅科蜡梅属落叶灌木,因其芳香馥郁,色如蜜蜡而得名,并同山茶、水仙及白梅一起被称为“雪中四友”。蜡梅是我国传统名贵花木,已有一千多年的栽培历史,亦是少有的冬季开花的园林植物,具有极高的观赏价值和经济价值。
3.蜡梅具有彩化和香化环境的作用。独特的物候期使得蜡梅在寒冷的冬季绽放,为寒冬增添色彩和馨香,是营造园林景观不可缺少的园林植物。蜡梅因其宜人的香气和独特的开花时间,在日本、欧美等地被广泛用作造景植物,具有很高的观赏价值,同时,它对多种有毒有害气体,特别是氯气和二氧化硫,具有很强的吸附作用,是美化和净化城市环境的重要树种。
4.蜡梅花蕾内花被片近轴端的蜜腺中可以散发出挥发性萜类化合物和苯类化合物,这类化合物具有浓郁的香味,因此,在蜡梅花朵中提取出的精油被广泛用作香水及各种调味品的主要成分,同时,蜡梅花精油还富含多种生物活性成分,如酚类和黄酮类化合物,因具有明显的消炎、抗氧化及防衰老等功能而被广泛用作化妆品原料;蜡梅花朵还具有重要的药用价值,其提取物是天然杀菌剂的潜在来源,同时还应用于制药工程,主要用于治疗咳嗽、风湿和麻疹;值得一提的是,在蜡梅果实和叶片中分离提取得到的倍半萜类化合物对癌细胞具有细胞毒性,从而起到抑制癌细胞生长的作用,为药物研发提供了新的方向和思路。
5.1998年,陈龙清等率先采用rapd(random amplified polymorphic dna, rapd)分子标记技术,从dna水平上分析了蜡梅天然居群的遗传多样性及遗传分化,为蜡梅的种质资源保存、引种驯化、遗传改良及新品种培育提供了重要的科学依据;2007年,赵凯歌等首次采用issr(inter-simple sequence repeat,issr)和rapd 分子标记技术报道了蜡梅栽培区的划分及不同栽培区蜡梅的遗传变异,为优化育种取样策略提供了重要数据支持,也为蜡梅种质资源的改良、鉴定及保存提供了重要理论基础;2009年,华中农业大学利用aflp(amplified fragment lengthpolymorphism,aflp)和issr分子标记技术构建了首个蜡梅种内遗传连锁图谱,该遗传图谱作为qtl(quantitative trait locus,qtl)定位的框架,为蜡梅分子育种提供了重要的遗传信息;近年来,高通量测序技术和基因挖掘技术的飞速发展,促进了解析基因表达差异、新基因发掘及基因功能研究的进步,同时,推动了分子育种技术的发展。2012年,眭顺照等构建了西南大学第一个蜡梅cdna(complementarydna,cdna)文库,并利用est(expressed sequence tag,est)技术鉴定出了95 个与环境胁迫及植物防御相关的基因,19个与花发育相关的基因,为蜡梅功能基因的研究提供了重要的基因资源;随后,刘道凤等构建了蜡梅花发育不同时期的转录组数据库,李志能等构建了低温打破蜡梅花芽休眠并使其膨大开放的转录组数据库,这些数据库为蜡梅花发育分子机制的研究提供了重要的数据资源。2019年和 2020年,南京林业大学和菏泽学院先后完成了素心
蜡梅(c.praecox var.concolor)和磬口蜡梅(c.praecox var.grandiflorus)的叶绿体基因组测序,为蜡梅的遗传育种、群体遗传学及dna条形码的研究提供了重要的数据参考和选择;2020年和2021年,尚均忠等和沈植国等分别完成了蜡梅
‘
h29’(c.praecox
‘
h29’)和红花蜡梅
‘
鸿运’(c.praecox
‘
hongyun’)的全基因组测序及染色体水平的组装,补全了蜡梅分子生物学研究缺少基因组序列的不足,为解析蜡梅遗传背景、改良蜡梅观赏性状及提高蜡梅抗逆性提供了全新的基因储备和数据资源,同时也翻开了蜡梅分子生物学研究的新篇章。
6.现有的对cbl家族的研究大多是和逆境胁迫相关,而关于cbl家族成员对花发育调控的研究鲜有报道。本课题组前期对蜡梅进行了转录组测序,构建了低温打破蜡梅花芽休眠并使其膨大开放的转录组数据库。在分析该数据库中所有cbl家族成员在蜡梅花发育不同时期的表达模式时,发现cbl所有转录本中有1个关键基因,它的表达量在花发育不同阶段均具有显著差异,因此猜测该基因可能参与花期调控,同时,在蜡梅花芽需冷量积累的不同阶段,它的表达量也出现了显著变化,推测该基因可能还受低温诱导,参与响应低温胁迫。
7.为了验证这个基因的功能,首先在基因组水平上对蜡梅cbl家族成员进行筛选与鉴定,获得了蜡梅全部的cbl家族成员序列,然后根据基因鉴定结果,对转录组数据库中的cbl的转录本id进行了重命名,即cpcbl8,最后对cpcbl8进行了重点研究,以期解析它在蜡梅生长发育过程中的作用及分子机制,这为蜡梅遗传改良、新品种培育及花期调控提供重要的方法和策略,有利于更好地将蜡梅资源应用到园林造景中。
技术实现要素:
8.本发明的目的是提供蜡梅cpcbl8及其编码的蛋白与应用。
9.首先,本发明提供蜡梅cpcbl8蛋白,其为:
10.1)由seq id no.2所示的氨基酸组成的蛋白质;或
11.2)在seq id no.2所示的氨基酸序列中经取代、缺失或添加一个或几个氨基酸且具有同等活性的由1)衍生的蛋白质。
12.本发明还提供编码所述的蜡梅cpcbl8蛋白的基因。
13.优选的,所述基因的序列如seq id no.1所示。
14.本发明还提供含有所述基因的载体,宿主细胞和工程菌。
15.本发明还提供所述基因在延迟植物开花、提高植物耐旱性、提高植物耐盐性和/ 或提高植物对低温的敏感性中的用途。
16.在本发明一个实施方案中,将所述基因转入植物基因组中,并在转基因植物中超量表达,并在转基因植物中超量表达,使所述植物延迟开花、提高耐旱性、提高耐盐性和/或提高对低温的敏感性。
17.本发明还提供一种使植物延迟开花的方法,其为将含有所述基因的载体转入所述植物基因组中,并在转基因植株中超量表达。
18.本发明还提供一种提高植物耐旱性、耐盐性和/或对低温的敏感性的方法,其为将含有所述基因的载体转入所述植物基因组中,并在转基因植株中超量表达。
19.本发明的有益效果如下:
20.本发明从蜡梅中克隆到了cpcbl8的全长。表达特性分析发现,cpcbl8在蜡梅的叶
中表达量最高,其次是芽,在花中的表达量最低;在花的不同部位,中/内花被片中cpcbl8的表达量显著高于外花被片、雄蕊和雌蕊。亚细胞定位实验表明cpcbl8 定位在质膜和细胞核。与wt相比,35s::cpcbl8/拟南芥花期显著推迟,莲座叶数量显著增多,叶面积显著增大,花序显著增高,并且cpcbl8表达量越高,表型变化越显著。测定花发育相关内源基因发现,与wt相比,过表达株系soc1、ap1和lfy 表达量显著下调。盐和干旱胁迫实验表明,与wt相比,过表达株系萎蔫程度更低,超氧化物歧化酶活性、相对含水量、叶绿素及脯氨酸含量更高,但气孔导度更小,丙二醛含量和活性氧积累更少,电解质渗漏率更低,而低温胁迫下各项生理指标测定结果完全相反。
21.综上,cpcbl8是盐和干旱胁迫响应的正向调节因子,是提高蜡梅耐盐和耐旱能力的潜在基因。蜡梅cpcbl8异源表达拟南芥可增强过表达拟南芥的耐盐和耐旱能力,但使其对低温胁迫不耐受,并具有抑制花发育的作用。
附图说明
22.图1所示为cpcbl8最大orf序列克隆凝胶电泳图。m为dna分子量标准 al2000,1、2、3和4为不同单克隆。
23.图2所示为pcambia-1300mcherry-cpcbl8双酶切验证图。m为dna分子量标准al2000,1、2和5分别为未酶切的pcambia-1300mcherry质粒、酶切过的 pcambia-1300mcherry质粒以及酶切过的cl-cpcbl8质粒;3和4为酶切后的pcambia-1300mcherry-cpcbl8重组载体。
24.图3所示为pcbl8亚细胞定位实验结果。a、b、c分别为阳性对照、阴性对照及cpcbl8实验组;merge、bright、nls和mcherry分别为混合场、明场、核定位信号及荧光信号。比例尺为50μm。
25.图4所示为cpcbl8在蜡梅不同器官、组织的相对表达量。
26.图5所示为pcambia-2301g-cpcbl8表达载体双酶切验证。m为dna分子量标准al2000,1为未酶切的pcambia-232301g载体,2、3和4分别为酶切后的 pcambia-232301g、pcambia-2301g-cpcbl8及tr-cpcbl8。
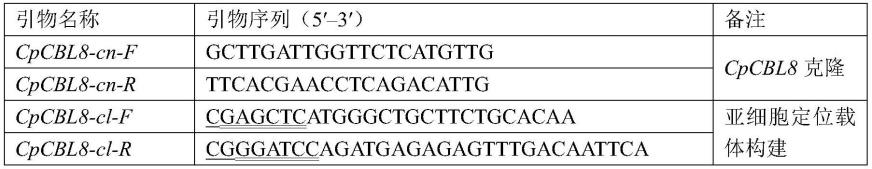
27.图6所示为t1代cpcbl8过表达株系检测。m为dna分子标准量al2000,n 为阴性对照,t为阳性对照,2~33及wt为不同拟南芥株系。
28.图7所示为不同拟南芥过表达株系cpcbl8表达水平。数据为平均值
±
标准误差,单因素方差分析及邓肯多重比较,不同字母表示显著差异且p《0.05。
29.图8所示为35s::cpcbl8/col-0和wt株系表型。比例尺为3厘米。
30.图9所示为不同浓度盐胁迫下35s::cpcbl8/拟南芥株系发芽率(a)及根长(b 和c)。a和b为平均值
±
标准误差,多重t检验,wt为对照组,*p《0.05,**p ≤0.01;c图比例尺为0.5厘米。
31.图10所示为盐胁迫下35s::cpcbl8/拟南芥株系表型。比例尺为3厘米。
32.图11所示为不同浓度甘露醇胁迫下35s::cpcbl8/拟南芥株系发芽率(a)及根长(b和c)。a和b为平均值
±
标准误差,多重t检验,wt为对照组,*p《0.05, **p≤0.01;c图比例尺为0.5厘米。
33.图12所示为干旱胁迫下35s::cpcbl8/拟南芥株系表型。比例尺为3厘米。
34.图13所示为冷胁迫下35s::cpcbl8/拟南芥株系表型。比例尺为3厘米。
35.图14所示为冷胁迫后35s::cpcbl8/拟南芥株表型叶绿素荧光参数。a图比例尺为3厘米,e图比例尺为1厘米。
36.图15所示为35s::cpcbl8/拟南芥株系离体叶片自然失水率。(a)及不同胁迫处理后的气孔导度(b和c)。图a和b数据为平均值
±
标准误差,多重t检验, wt为对照组,图a颜色相同的星号和折线代表同一株系。*p《0.05,**p≤0.01,图c比例尺为10微米。
37.图16所示为35s::cpcbl8/拟南芥过表达株系胁迫处理后的生理指标。图a~f 数据为平均值
±
标准误差,多重t检验,wt为对照组,*p《0.05,**p≤0.01;图 g和h比例尺为1厘米。
38.图17所示为35s::cpcbl8/拟南芥株系内源基因相对表达量。数据为平均值
±
标准误差,多重t检验,wt为对照组,*p《0.05,**p≤0.01。
具体实施方式
39.以下实施例用于说明本发明,但不用来限制本发明的范围。
40.实施例1蜡梅cpcbl8基因的克隆
41.以西南大学校园内长势良好且无病虫害的
‘
磬口’蜡梅叶片为材料,进行总 rna的提取,以提取的rna为模板进行cdna第一链的合成。
42.利用primer premier 5软件,以低温打破蜡梅花芽休眠并使其膨大开放的转录组数据库中筛选出的cpcbl8序列为模板,进行特异性扩增引物设计,pcr产物要包含目的基因的最大orf(open reading frame,orf)序列。设计好的引物送华大基因公司合成,具体引物序列见表1。
43.表1引物列表
[0044][0045]
以cpcbl8-cn-f和cpcbl8-cn-r为引物进行cpcbl8全长的pcr扩增,对 cpcbl8的pcr扩增产物进行凝胶电泳检测,结果表明各产物长度和预测的pcr产物长度基本一致。将目的基因连接到pmd
tm
19-载体后进行菌落pcr及测序,菌落pcr结果和基因克隆pcr结果一致,结合测序结果可知克隆得到的cpcbl8长893bp (图1),包含完整的663bp cds,序列如seq id no.1所示,与转录组数据库中的序列一致,编码蛋白的氨基酸序列如seq id no.2所示,本步骤重组质粒命名为 cn-cpcbl8。
[0046]
利用t4连接法和同源重组的方法获得了pcambia-1300mcherry-cpcbl8重组载体,对重组载体进行pcr扩增检测,条带长度正确;对相应的样品菌液进行测序,测序结果反馈无突变、缺失或插入位点;对重组载体进行双酶切验证,结果表明, pcambia-1300mcherry-cpcbl8可被限制性核酸内切酶切割成双条带,且条带长度正确(图2)。
[0047]
亚细胞定位实验结果表明,在对照组,携带pcambia-1300mcherry空载的农杆菌侵染洋葱表皮细胞后,可以在洋葱表皮细胞的各个位置发现荧光信号(图3a),而不携带载体
cpcbl8重组质粒进行双酶切,获得线性化的pcambia-2301g及带酶切位点的目的基因。利用琼脂糖凝胶电泳及胶回收的方法获得相应的片段,其中 pcambia-2301g表达载体酶切产物回收长片段,tr-cpcbl8重组质粒酶切产物回收短片段。利用t4连接法将tr-cpcbl8短片段与pcambia-2301g长片段连接,获得重组载体。将连接产物转化至大肠杆菌感受态细胞中,然后将转化产物涂布在lb固体培养基平板(含50mg/l kan)上;将培养基平板倒置放在恒温培养箱中,设置条件37℃,培养11~12h;挑取培养后的数个单克隆菌落至1000μl lb液体培养基 (含50mg/l kan)中,然后利用恒温振荡器扩繁,设置条件37℃、180rpm,培养 11~12h;培养完成后利用pcr扩增及琼脂糖凝胶电泳的方法检测阳性菌株。将经凝胶电泳检测条带长度正确的阳性菌株送擎科生物公司(重庆)测序,对测序正确的单菌落进行扩繁及重组质粒提取,对重组质粒进行双酶切验证(图5)。将验证无误的载体命名为pcambia-2301g-cpcbl8,获得cpcbl8遗传转化重组载体。
[0060]
通过电转化的方法将重组载体pcambia-2301g-cpcbl8转到农杆菌感受态细胞gv3101(psoup)中,利用浸花法转化拟南芥。利用ms培养基(含kan 50mg/l) 对过表达株系进行筛选,获得了t1代过表达株系,以t1代植株叶片为材料进行阳性植株检测,结果如图6,其中阴性对照为同体积超纯水,阳性对照为 pcambia-2301g-cpcbl8重组载体。由电泳图可以看出阴性对照及wt株系无条带,过表达株系条带和阳性对照条带长度一致,4、10、17、26、27及28号株系所在泳道无条带,对pcr检测阳性的株系继续进行筛选,以获得纯合过表达株系。
[0061]
为选出不同cpcbl8表达量的株系,以t2代纯合35s::cpcbl8/拟南芥幼苗为样品,进行qrt-pcr,根据结果可以看出,表达量最高的三个株系分别为oe13-1、oe3-1 和oe19-2(图7),这3个株系在过表达株系阳性鉴定电泳图中分别对应条带较清晰的13、3和19(图6)。oe13-1株系的cpcbl8表达量显著高于oe3-1和oe19-2, oe3-1株系的cpcbl8表达量也显著高于oe19-2;oe31-11株系的表达量较低,但其表达量和wt株系也具有显著差异;oe8-5、oe13-2、oe15-3、oe30-1及oe31-5株系基本无表达,他们的表达量和wt株系无显著差异。通过dna检测及表达量检测相结合的方法最终选择oe13-1、oe3-1和oe31-11分别作为35s::cpcbl8/col0高、中和低表达株系,这3个株系将用于后续cpcbl8基因功能验证。
[0062]
对筛选出的t2代纯合35s::cpcbl8/拟南芥株系进行表型观察,发现oe13-1、 oe3-1及oe31-11植株的抽薹时间和莲座叶数均与wt具有显著差异(图8,表4),过表达植株抽薹时间显著推迟,莲座叶数量显著增多,同时,oe13-1和oe3-1植株的莲座叶面积及花序高度与wt也均具有显著差异,五周龄时过表达拟南芥莲座叶更大,八周龄时的过表达拟南芥花序更高。
[0063]
表4 35s::cpcbl8/col-0和wt株系表型统计
[0064]
株系表型wtoe13-1oe3-1oe31-11抽薹时间(d)28.75
±
0.22b31.30
±
0.33a30.90
±
0.39a30.70
±
0.35a莲座叶数(片/piece)12.05
±
0.21b15.25
±
0.40a14.45
±
0.44a14.45
±
0.37a莲座叶长(cm)1.87
±
0.04c2.77
±
0.05a2.49
±
0.03b1.95
±
0.04c莲座叶宽(cm)0.99
±
0.02c1.31
±
0.03a1.2
±
0.02b0.92
±
0.03c花序高度(cm)24.76
±
1.19c35.40
±
0.26a32.15
±
0.22b26.85
±
0.54c[0065]
实施例4过表达拟南芥耐盐性鉴定
[0066]
将筛选出的cpcbl8表达量差异显著的t2代纯合35s::cpcbl8/拟南芥及wt株系的种子播种在模拟不同程度盐胁迫(0、10、20、50、100mm nacl)的ms培养基上,其中表达株系播种于培养基含kan 50mg/l的组培瓶中,使植株根系有充足的空间竖直生长。12d后统计不同nacl浓度下各株系的萌发率。使用根系扫描仪扫描不同浓度nacl胁迫下各株系的根,然后根据扫描时画的标尺对根长进行测量。结果发现,cpcbl8高表达的株系耐盐能力更强。在10、20、50及100mm的nacl胁迫下, oe13-1和oe3-1株系种子的萌发率均显著大于wt株系(图9a),cpcbl8低表达的oe31-11株系种子萌发率和wt株系无显著差异。对幼苗根长进行测量,结果发现在10、20、50及100mm的nacl胁迫下,oe13-1株系的根长均显著大于wt株系 (图9b和c),在10mm的nacl胁迫下,oe3-1株系的根长也显著大于wt株系。以上结果表明,异源表达cpcbl8,过表达拟南芥的耐盐能力显著增强。
[0067]
将一部分播种在不含nacl的ms培养基平板上的各株系幼苗(14d龄)移栽到装有基质的方盆中,每个方盆移栽25株左右,常规培养3w后,进行盐胁迫处理。每盆浇灌150mm nacl溶液100ml,每个处理重复三次,胁迫期间进行拍照记录。
[0068]
对移栽到方盆中的t2代纯合35s::cpcbl8/拟南芥进行盐胁迫处理并表型观察,处理前各株系浇足水分以保证生长正常,缓苗3天后用150mm nacl溶液浸盆处理。实验结果表明,在浇灌nacl第3天时,各株系拟南芥开始出现胁迫表型,莲座叶开始萎蔫、枯黄(图10);在第六天时,wt株系已死亡近半,而oe13-1、oe3-1和 oe31-11仅有个别植株开始死亡;在第10天时,wt株系已基本死亡,而cpcbl8高表达量的株系oe13-1仅有部分植株死亡;在第13天时,oe13-1植株存活率大于50%,oe3-1株系植株存活率近40%,oe31-11存活率近20%。以上结果进一步表明,过表达cpcbl8可增强过表达拟南芥的耐盐能力。
[0069]
实施例5过表达拟南芥耐旱性鉴定
[0070]
将筛选出的cpcbl8表达量差异显著的t2代纯合35s::cpcbl8/拟南芥及wt株系种子播种在模拟不同程度干旱(0、50、100、200、300mm甘露醇)的ms培养基上,其中表达株系播种于培养基含kan 50mg/l的组培瓶中,使植株根系有充足的空间竖直生长。12d后统计不同浓度甘露醇胁迫下各株系的萌发率(萌发的标准为幼苗长出两片完整的绿色子叶),使用根系扫描仪扫描不同甘露醇浓度下各株系的根,然后根据扫描时画的标尺对根长进行测量。结果表明,cpcbl8高表达的株系具有更强的耐旱能力。在50、100、200及300mm的甘露醇胁迫下,oe13-1种子的萌发率均显著大于wt株系,除200mm外,oe3-1的种子萌发率也显著大于wt,而在50 mm和100mm的甘露醇胁迫下,oe31-11种子萌发率也显著大于wt株系(图11 a)。对幼苗根长进行测量,结果发现在50、100、200及300mm的甘露醇胁迫下, oe13-1和oe3-1的根长均显著大于wt株系,在50mm的甘露醇胁迫下,oe31-1 的根长也显著大于wt株系(图11b和c)。以上结果表明,异源表达cpcbl8,过表达拟南芥的耐旱能力显著增强。
[0071]
将一部分播种在不含甘露醇的ms培养基平板上的各株系幼苗(14d龄)移栽到装有基质的方盆中,每个方盆移栽25株左右,常规培养3w后,进行干旱处理。处理前使每盆土壤基质充分吸水,缓苗24h后进行干旱胁迫处理,采取自然干旱的方法,约2周后进行复水,每个处理重复三次,干旱期间进行拍照记录。
[0072]
对移栽到方盆中的t2代纯合35s::cpcbl8/拟南芥进行自然干旱处理并观察表型,处理前给土壤基质浇足水分以确保各株系生长正常。实验结果表明,在停止浇水第6天时,方盆质量明显变轻,表明种植基质中的水分严重不足,此时各株系拟南芥开始出现胁迫表
型,如叶片萎蔫、叶边缘枯黄及叶基部发紫(图12);在停止浇水第 10天时,wt株系的植株体积已明显减小,而过表达株系oe13-1、oe3-1和oe31-11 仍有较多叶片正常生长;在停止浇水第13天时,wt株系已基本无鲜绿色叶片,而过表达株系oe13-1、oe3-1和oe31-11仍有一小部分鲜绿色叶片;对干旱13天的各株系进行恢复供水,在复水第3天时,发现wt株系有近20%的植株彻底死亡,剩余 80%得以恢复正常,而过表达株系oe13-1、oe3-1和oe31-11株系各植株基本全部恢复正常。以上结果进一步表明,异源表达cpcbl8可增强过表达拟南芥的耐旱能力。
[0073]
实施例6过表达拟南芥耐寒性
[0074]
以移栽到方盆中常规培养3w后的t2代纯合35s::cpcbl8/拟南芥及wt株系为材料,每个株系取3盆,每盆移栽25株左右,进行低温胁迫处理。首先在人工气候箱中4℃冷驯化9h后置于-4℃、黑暗条件下冷冻处理9h,然后在4℃、黑暗条件下解冻9h,最后转移至常规培养条件下恢复7d,对不同处理时期的植株进行拍照记录。实验结果表明,在4℃冷驯化9h后,各株系仍生长正常,无明显表型变化(图 13);将各株系进行-4℃低温冷冻处理,9h后,oe13-1和oe3-1株系严重萎蔫,而wt和oe31-11株系仅有部分植株出现萎蔫;将冷冻处理后的各株系放到4℃黑暗条件下恢复,9h后,oe13-1和oe3-1株系无明显恢复迹象,而wt和oe31-11 株系部分植株已恢复正常;将各株系转移到温室中常规培养7天后,发现各株系已恢复正常,但oe13-1和oe3-1株系植株死亡近50%,而wt和oe31-11株系除部分植株死亡外,多数植株仍正常生长,特别是wt株系,基本无植株死亡。以上结果表明,异源表达cpcbl8,过表达拟南芥对低温更加敏感。
[0075]
对过表达株系进行-5℃处理1小时,统计各株系冻伤程度,并测定各株系叶绿素荧光参数。让-5℃处理1h后的各株系暗适应30min后,使用调制式叶绿素荧光成像系统拍照并测定各株系最大光化学效率fv/fm和电子传递量子速率φ
psⅱ,每个株系至少测定3个单株,进一步确定转基因拟南芥的耐寒性。结果表明,在未处理的对照组,各株系均生长正常,各株系最大光化学效率(fv/fm)(图14a、b和e) 和psⅱ电子传递量子效率(φpsⅱ)(图14c)无显著差异;经过冷冻处理后,各株系均出现冻伤,但wt株系冻伤程度更低,其正常植株占比显著高于oe13-1株系;同时,各株系fv/fm和φpsⅱ均出现下降,表明psⅱ受到损伤,但oe13-1 和oe-31的fv/fm和φpsⅱ均显著低于wt,表明过表达株系psⅱ受损程度更高。以上结果进一步表明异源表达cpcbl8,过表达拟南芥对低温更加敏感。
[0076]
实施例7过表达拟南芥离体叶片自然失水率及不同胁迫下的气孔导度
[0077]
为了探究cpcbl8在参与植株保水能力方面发挥的作用,对过表达株系的离体叶片进行了连续8小时的失水率测定。通过测定结果可以发现,wt株系在离体1h 时的失水率就显著高于oe13-1和oe3-1(图15a),在离体2h后,wt株系失水率显著高于oe3-1,离体3h后显著高于oe13-1。在之后的时间段中,各株系间失水率无显著差异,但在8h后,wt株系失水率已达68%,而oe13-1和oe3-1株系分别为52%和61%,失水率差异显著,同时,cpcbl8低表达株系oe31-11和wt株系在整个阶段的失水率均无显著差异。由此可以推测出异源表达cpcbl8增强了过表达拟南芥在脱水早期阶段(0~3h)的保水能力。
[0078]
气孔运动对植物抵抗逆境胁迫具有重要意义。为了验证35s::cpcbl8/拟南芥抗逆性强弱,对各株系离体叶片进行了胁迫处理,并观察气孔导度变化,统计气孔长宽比,每个株系观察气孔30个以上。由实验结果可以发现,在正常生长条件下,各株系的气孔导度无显
著差异(图15b和c);对叶片进行模拟盐(50mm nacl)和干旱(300mm甘露醇)胁迫发现,oe13-1和oe3-1的气孔导度显著小于wt;在低温 (4℃)胁迫下,oe13-1、oe3-1和oe31-11的气孔导度均显著大于wt,说明异源表达蜡梅cpcbl8,过表达拟南芥可以通过气孔运动降低蒸腾作用,从而增强耐盐和耐旱能力,但却对低温变得更加敏感。
[0079]
实施例8过表达拟南芥胁迫处理后的生理指标
[0080]
对35s::cpcbl8/拟南芥分别进行了300mm nacl、30%peg6000和4℃低温胁迫处理,然后测定多个生理指标。结果表明,在对照实验中,除oe13-1的叶绿素含量显著高于wt,丙二醛含量显著低于wt株系外,其他指标均无显著差异(图16b 和e)。在盐和干旱胁迫下,oe13-1株系的相对含水量、叶绿素和脯氨酸含量显著高于wt株系(图16a~c),电解质渗漏率和丙二醛含量也显著低于wt株系(图16 d和e),而超氧化物歧化酶的活性则显著高于wt株系(图16f),同时h2o2(图 16g)和o
2-(图16h)积累也明显比wt株系少;在盐胁迫下,oe3-1株系除叶绿素和脯氨酸含量与wt株系无显著差异外,其他生理指标测定结果均表明其比wt具有更强的耐盐能力;在30%peg6000模拟干旱胁迫下,除丙二醛含量与wt株系无显著差异外,其他生理指标均表明其比wt株系具有更强的耐旱能力;值得注意的是,虽然在盐和干旱胁迫下,oe3-1株系的以上三个指标与wt株系无显著差异,但其叶绿素和脯氨酸含量的平均值高于wt,丙二醛含量平均值低于wt;以上结果表明 oe13-1和oe3-1比wt具有更强的耐盐和耐旱能力。另一方面,低温胁迫后oe13-1 的生理指标与盐和干旱胁迫后的测定结果完全相反,表明oe13-1在低温胁迫下损伤程度更高,同时,低温胁迫后oe3-1的丙二醛含量显著高于wt,脯氨酸含量显著低于wt,相对含水量、叶绿素含量及sod活性的平均值也低于wt,表明oe3-1损伤程度更高。以上实验结果进一步证明了异源表达cpcbl8可增强过表达拟南芥的耐盐和耐旱能力,但使其对低温胁迫更加敏感。
[0081]
实施例9过表达拟南芥表型及耐寒性变化的分子机制
[0082]
为了探究cpcbl8在植物花期调控及低温胁迫响应分子机制中扮演的角色,对与花期调控及低温胁迫应答相关的拟南芥内源基因进行了表达分析。选择actin2作为内参基因,ft(flowering locus t)、soc1(suppressor of overexpression of co1)、 lfy(leafy)和ap1(apetala 1)作为花发育调控通路相关基因,cbf1/2/3(c-repeatbinding factor 1/2/3)、cor47/15a/15b(cold regulated 47/15a/15b)、crpk1 (cold-responsive protein kinase 1)及14-3-3λ(也称g-box regulating factor 6,grf6) 作为低温响应通路相关基因,进行qrt-pcr实验,以探究cpcbl8在过表达拟南芥中发挥功能的分子机制。qrt-pcr所需引物由ncbi在线引物设计工具设计,送擎科生物公司合成,具体引物序列见表5。
[0083]
表5引物序列
[0084][0085][0086]
鉴于本研究是基于低温打破蜡梅花芽休眠并使其膨大开放的转录组数据展开的,主要涉及低温诱导及花发育,因此重点研究了花发育及低温胁迫响应的相关分子机制。测定过表达拟南芥花发育及耐寒性调控通路相关基因的相对表达量,qrt-pcr结果表明,在正常生长条件下,过表达株系soc1、lfy和ap1相对表达量显著低于 wt,ft的相对表达量与wt无显著差异(图17a);在未进行低温处理的对照组, oe13-1株系的cprk1、14-3-3λ和oe3-1株系的cprk1相对表达量极显著高于wt (图17b),oe13-1株系的cbf1/2/3、cor15a/15b/47和oe3-1株系的cor15a/47 相对表达量极显著低于wt,同时oe3-1株系cbf3的相对表达量显著低于wt(图 17c);低温处理后,和wt相比,过表达株系oe13-1的cbf1/3、oe3-1的cbf1、 oe13-1/3-1的cor15a/47及oe13-1的cor15b相对表达量均极显著下调,cor15b 在oe3-1中表达量显著下调,同时,以上两个过表达株系的14-3-3λ极显著上调,而 crpk1相对表达量平均值上调,但不具有显著性。
[0087]
虽然,上文中已经用一般性说明及具体实施方案对本发明作了详尽的描述,但在本发明基础上,可以对之作一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1