一株屎肠球菌菌株及其用途
1.本发明涉及一株屎肠球菌菌株,还涉及该屎肠球菌菌株在抗炎和抗感染制剂中的用途,本发明属于生物技术领域。
背景技术:
2.屎肠球菌(enterococcus faecium)是肠球菌属的重要成员之一,大多数为任何动物的肠道黏膜表面的益生菌之一。益生性乳酸菌(lab)是人和动物肠道、呼吸道、生殖道及皮肤黏膜系统的正常菌群,许多研究成果发现乳酸菌和肠黏膜共同组成保护屏障,防止病毒、细菌等的入侵,人类越来越清晰地认识到益生菌对人类和动物生命健康的强大作用。近年来最令人兴奋的科学进展之一是认识到共生微生物(我们的微生物群)在人类生理营养、维生素合成、药物代谢、预防感染和疾病康复方面发挥着重要作用。
3.近年来,益生菌的免疫调节作用受到人们的广泛关注,免疫系统已进化为维持宿主-微生物群的共生关系,另一方面,为了维持宿主免疫稳态,肠道菌群往往发挥着一种不容忽视的免疫调节功能,脑-肠轴是肠道微生物群、肠道屏障和免疫系统之间的双向连接,在人体免疫、代谢、结构和神经系统等方面发挥着一些基本功能。肠道菌群对个体的生理和心理健康也有重要影响。研究表明,一些乳酸菌菌株能够刺激机体免疫应答,例如lactococcus lactis、lactobacillus paracasei等能够促进iga生成、增强nk细胞活性以及通过激活巨噬细胞或树突状细胞来增强特异性免疫反应;有的益生菌可刺激机体释放不同的细胞因子,来影响黏膜免疫反应和t细胞分化,从而对宿主进行免疫调节;有的益生菌能够激活机体树突状细胞,并辅助t细胞极化,使ifn-γ和il-12分泌升高,从而使幼稚t细胞或记忆t细胞转化为th1,进而使机体提高免疫力实现抗感染;除此之外,有的益生菌在增殖过程中还会分泌一些代谢产物,某些代谢产物具有抗炎作用,可减弱th17的极化,有利于treg 1型辅助性t细胞的分化。有的益生菌可通过其mamp(如脂磷壁酸、肽聚糖、s层蛋白和核酸)与宿主肠粘膜表达的pprs(toll样受体、nod样受体,c型凝集素受体)结合来调节宿主的免疫系统。
4.益生菌的抗炎、抗感染作用近年来备受关注,也已被广泛研究和应用,以往的研究主要集中在利用益生菌缓解炎症相关疾病,如过敏症状、炎症性肠病和自身免疫性疾病等方面。有研究表明,口服罗氏乳杆菌和干酪乳杆菌后,通过活化骨髓来源的树突状细胞上的受体来上调treg细胞进行免疫调节和免疫耐受,同时上调免疫负调控细胞因子il-10,最终显著下调促炎级联反应,减少了过敏反应对机体的损害;还有研究表明,一种益生菌制剂(l.rhamnoses,b.lactis,以及b.longum对炎症因子的分泌具有调节能力,能诱导人巨噬细胞系thp1产生抗炎因子il-10,显著下调促炎因子il-1β与il-6的表达;lactobacillus rhamnoses 4b15以及lactobacillus gasseri 4m13能够抑制lps刺激巨噬细胞产生一氧化氮,并且也能够降低炎性因子il-1β、il-6;在一项小鼠感染甲型流感的实验中,一株加式乳杆菌可降低小鼠肺部促炎细胞因子il-6的浸润,并显著降低了病毒滴度。益生菌的抗炎作用机制也有多方面的报道,例如由其代谢产物介导抗炎作用(例如短链脂肪酸(scfa),特别
是丙酸、乙酸和丁酸),scfa可由双歧杆菌、乳酸杆菌和多种共生细菌产生,它们通过与肠上皮细胞上的特定受体结合来发挥抗炎作用,抑制中性粒细胞和巨噬细胞产生促炎细胞因子等。有研究探讨了世界各地广泛使用的益生菌干酪乳杆菌菌株shirota(lcs)对急性肝损伤(ali)的影响及其潜在机制,发现干酪乳杆菌shirota通过重塑肠道微生物群来减轻过度炎症和代谢紊乱,从而预防急性肝损伤;口服瑞士乳杆菌la401和gasseri乳杆菌gasserila806组合可减轻小鼠的食道和胃肠道念珠菌感染引起的肠道炎症。此外,益生菌还可以减轻肠道炎症,降低肠道和肿瘤中主要致炎细胞因子il-6、tnf-α、il-17、il-23和il-1β的水平。
5.总之,许多益生菌的抗炎作用是通过免疫调节介导的,特别是通过促炎和抗炎细胞因子的平衡控制,越来越清楚的是,许多因素参与免疫调节平衡,特别与遗传背景、宿主免疫反应和微生物多样性状态等组合调节有关;多项研究通过不同研究模型(细胞系、结肠炎动物模型、临床研究)评估了益生菌在疾病治疗、预防和缓解炎症方面的作用。有研究数据表明,各种候选益生菌的免疫调节作用存在差异,免疫调节作用和方式千差万别,例如不同的菌株对相同的炎症具有明显的差别,这些结果表明,益生菌的免疫调节特征必须进行具体、复杂的检测和表征后才能应用。并且大多数已应用的益生菌株的确切抗炎机制尚未完全阐明。
6.尽管最初的炎症反应对清除感染是至关重要的,是启动免疫防御和免疫应答所必需的,然而,强烈的炎症反应会导致细胞因子风暴(cytokine storm),最终会导致组织和器官损伤而死亡。感染人类的冠状病毒,包括非典冠状病毒、中东呼吸综合征冠状病毒和新型冠状病毒,感染初期会引发强烈的炎症反应,其中在新型冠状病毒(covid-19)感染的重症患者中能观察到细胞因子风暴和促炎细胞因子级联反应,进而导致肺部的严重损伤。
7.长期以来,急性炎症风暴的治疗主要是通过限制由激活的免疫系统引起的附带损害来维持关键器官功能,例如使用糖皮质激素和地塞米松,通过抑制促炎细胞因子的合成和驱动淋巴细胞死亡,起到阻止炎症风暴的作用。益生菌能维持肠道微生物动态平衡,维护肠道微生物中的优势共生菌群,可以帮助机体调节由病原体感染引发的炎症反应与机体正常免疫功能之间的平衡,从而使机体的整个免疫系统受益,近年来,益生菌在缓解急性炎症反应方面也有报道,补充益生菌显著减少了重症率和死亡率,增加了针对严重急性呼吸综合征冠状病毒2(sars-cov2)的特异性igm和igg。然而,这些报道均是将益生菌作为治疗过程中的辅助因子发挥了重要抗炎作用,如果将益生菌单独使用来进行治疗的情况下,是否对感染导致的急性炎症风暴具有较好的抑制作用,是否可以大大缓解临床急性炎症风暴导致的急性死亡,目前还缺乏这方面的研究报道。
技术实现要素:
8.本发明的目的在于提供一种屎肠球菌新菌株以及所述菌株在抗炎及抗感染,尤其是在抗急性炎症中的用途。
9.为了达到上述目的,本发明采用了以下技术手段:
10.本发明菌株分离自黑龙江省五常县凤凰山野生健康鸡盲肠粘膜上,经mrs接种培养、分离纯化后分离获得的一株乳酸菌株,经鉴定其属于屎肠球菌(enterococcus faecium),命名为c171,分类命名为屎肠球菌(enterococcus faecium),保藏在中国微生物
菌种保藏管理委员会普通微生物中心,地址在北京市朝阳区北辰西路1号院中科院微生物研究所,其微生物保藏号是cgmcc no.24533,保藏日期为2022年3月15日。该菌株的微生物学特征是:
①
革兰氏染色典型阳性,菌体为中等偏大,形态规则的球杆菌;需氧或兼性厌氧菌,在固体和液体mrs培养基上生长良好;
②
具有耐酸特性,可在ph 4.0的mrs上生长良好;
③
具有耐胆盐特性,可在含50%的鸡胆汁mrs的培养液中存活并生长;
④
具有独特的抗炎、抗感染特性;
⑤
该菌培养最适温度为37℃,mrs培养基上菌落为乳白色,边缘整齐,表面光滑湿润,隆起,不透明的中等大小的菌落,有一种酸香的味道。
11.因此,进一步的,本发明还提出了所述的屎肠球菌菌株在制备抗炎和抗感染制剂中的用途。
12.其中,优选的,所述的抗炎和抗感染,是指抵抗急性炎症风暴,包括抵抗细菌或病毒感染导致的急性炎症和死亡的作用。
13.其中,优选的,所述的急性炎症是由超强毒力病毒感染导致的。
14.更进一步的,本发明还提出了所述的屎肠球菌菌株在制备调节动物肠道菌群或促进动物生长的药物中的用途。
15.相较于现有技术,本发明的有益效果是:
16.本发明从野生健康鸡盲肠粘膜上分离纯化得到了一株乳酸菌株,经鉴定其属于屎肠球菌(enterococcus faecium),研究表明该菌株具有独特的抗炎和抗感染双重特性,同时还具有耐酸、耐胆盐等特性,仅口服该菌株屎肠球菌c171株可抑制由超强毒性病毒感染引发急性炎症反应而导致的死亡,降低动物死亡率80%以上。本发明公开的一株屎肠球菌新菌株可用作益生菌株用于食品、饲料和药物的添加剂,添加于食品、饲料或药物实现其对动物的抗炎和抗感染及保健作用等,有强抗炎作用又有抗感染作用的天然益生菌c171株有望作为各种急性炎症的辅助性治疗制剂,尤其是在动物遭受急性感染时,保护效果更佳。
附图说明
17.图1为本发明分离得到的一株屎肠球菌新菌株的菌株在固体培养基上的生长特性;
18.图2为肠球菌新菌株enterococcus faecium c171在x-gal-mrs培养基、肠球菌显色培养基、mc培养基固体培养基上的培养特性;
19.图3为屎肠球菌c171株与巨噬细胞炎症模型相互作用;
20.图4为屎肠球菌c171对鸡原代巨噬细胞的抗炎和抗感染特性;
21.图5为屎肠球菌c171株的抗炎作用是通过调控nlrp3炎性小体通路,抑制caspase-1的活性来实现抗炎的试验结果;
22.图6为屎肠球菌c171株在体外的抗病毒增殖特性的结果;
23.图7为口服屎肠球菌c171株的可显著保护鸡超强毒力病毒感染引发的死亡,保护率达80%以上的动物实验结果;
24.图8为口服屎肠球菌c171株的显著抑制了超强毒力病毒vvibdv引发鸡的急性炎症的结果;
25.图9为口服屎肠球菌c171株的可显著提高鸡在超强毒力病毒感染时的抗感染作用的结果;
26.图10为屎肠球菌c171株安全性评价,毒力因子(a)、耐药基因(b)、鸡胚安全性(c)检测结果;
27.图11为屎肠球菌c171株能够耐受胃肠道胆盐(a)及胃酸(b)环境的结果;
28.图12为口服屎肠球菌c171株对肠道菌群属水平top30菌群分析,a柱状图,b热图的分析结果;
29.图13为口服屎肠球菌c171株对肠道菌群种水平top30菌群分析,a柱状图,b热图的分析结果;
30.图14为口服屎肠球菌c171株对肠道菌群的小肠α多样性指数稀释曲线分析;小肠α多样性相关箱线图;小肠β多样性指数分析;
31.图15为口服屎肠球菌c171株对肠道菌群的小肠微生物多元变量统计分析。lefse分析。
具体实施方式
32.下面结合具体实施例来进一步描述本发明,本发明的优点和特点将会随着描述而更为清楚。但实施例仅是范例性的,并不对本发明的范围构成任何限制。本领域技术人员应该理解的是,在不偏离本发明的精神和范围下可以对本发明技术方案的细节和形式进行修改或替换,但这些修改和替换均落入本发明的保护范围内。
33.实施例1本发明菌株的分离及鉴定
34.1、菌株的分离和鉴定
35.该菌株分离自黑龙江省五常县凤凰山野生健康鸡盲肠粘膜上,在mrs培养基上生长良好,经划线分离获得的一株乳酸菌株。其微生物学特征是:革兰氏染色典型阳性,菌体为中等偏大,形态规则的乳球菌;需氧或兼性厌氧菌,在固体和液体mrs培养基上生长良好,该菌培养最适温度为37℃,mrs培养基上菌落为乳白色,边缘整齐,表面光滑湿润,隆起,不透明的中等大小的菌落,有一种酸香的味道。
36.菌株菌落特征如图1所示。
37.本发明的一株屎肠球菌新菌株,命名为c171,分类命名为屎肠球菌(enterococcus faecium)。其微生物保藏号是cgmcc no.24533,保藏日期为2022年3月15日。
38.2、菌株的耐酸试验:
39.将分离获得的菌株enterococcus faecium c171按mrs液体培养基体积的10%的接种量分别接种于ph值为3.0、4.0、5.0、6.5的mrs液体培养基中,每个酸度梯度设置3个平行对照,37℃静置培养,分别取培养30min、60min、90min、120min的菌液样品测定od
600
nm值。试验表明本发明菌株具有较强的抗酸能力,可在ph 4.0的mrs上生长。
40.3、菌株的耐鸡胆汁试验:
41.将分离菌株enterococcus faecium c171按mrs液体培养基体积的10%的接种量接种于含新鲜鸡胆汁含量分别为0%、10%、20%、30%、40%、50%、60%、70%、80%、90%的mrs培养基中,每个浓度设置3个平行试验,37℃静置培养,取培养30min、60min、90min、120min的菌液样品测定od
600
nm值,结果表明该菌株能在含30%的鸡胆汁培养基中很好地生长,具有较强的抗鸡胆汁能力。
42.实施例2本发明菌株的固体培养基上的培养特性
43.1、屎肠球菌c171株在普通mrs培养基上的菌落特征:
44.实验方法:将enterococcus faecium c171在mrs固体培养基划线培养,对c171的菌落形态进行观察。
45.实验结果:如图1,屎肠球菌c171菌落形态为边缘整齐,表面光滑湿润,隆起,不透明的中等大小的菌落,乳白色,有一种酸香的味道。
46.2、屎肠球菌c171株在含x-gal-mrs培养基、肠球菌显色培养基、mc培养基上的菌落特征:
47.实验方法:将enterococcus faecium c171分别划线在x-gal-mrs培养基、肠球菌显色培养基、mc培养基上,37℃培养48h,观察菌落特征。
48.实验结果:如图2,屎肠球菌c171株在x-gal-mrs上厌氧培养时,菌落形态与mrs培养基上相似(图2a);在x-gal-mrs上有氧培养时,菌落形态与mrs培养基上相似,颜色为蓝色(图2b)(说明屎肠球菌c171株可以产生β-葡萄糖苷酶,分解x-gal底物,使菌落为蓝色);屎肠球菌c171株在肠球菌显色培养基上,有氧培养后,菌落为典型的棕褐色(图2c);屎肠球菌c171株在mc培养基上,有氧培养后,菌落四周出现透明环(图2d),说明屎肠球菌c171株产生酸类代谢产物,溶解碳酸钙形成的。
49.实施例3屎肠球菌c171株与巨噬细胞炎症模型相互作用,呈现强烈的抗炎和抗感染的特性
50.实验方法:鸡巨噬细胞hd11s(1
×
106cells/ml)接种于12孔板中,细胞贴壁后加入50moi c171,37℃ 5%co2孵育24h,利用lps和nigericin使细胞发生急性炎症反应,建立急性炎症模型,即每孔加入工作浓度为1μg/ml的lps,37℃ 5%co2作用3h,再加入工作浓度为1μm的nigericin作用1h后,收集细胞沉淀用sybr green
ꢀⅰ
荧光定量pcr相对定量,利用表1的引物来分析细胞中il-1β和il-10mrna的转录水平。以lps和nigericin处理的细胞作为阳性对照,以未作任何处理的细胞为空白对照。
51.检测c171激活巨噬细胞nf-κb和irf信号通路的特性。j774-dual
tm
(2.8
×
105个/ml)接种于96孔板中,加入50moi的c171,以1μg/ml的lps和10ng/ml的pam3csk作为阳性对照,在37℃ 5%co2刺激24h后,收集上清液。然后在96孔板中每孔加入170μl quanti-blue
tm
),再加入30μl j774-dual
tm
细胞作用上清后置于37℃ 5%co24 h,测定650nm处的吸光度,来反映nf-κb诱导的seap水平。j774-dual
tm
cells可同时检测irf信号通路:j774-dual
tm
(2.8
×
105cells/ml)接种于96孔板中,加入50moi的c171,以1μg/ml的lps和1μg/ml的2
’3’‑
cgamp作为阳性对照,在37℃ 5%co2刺激24h后,收集上清液。在不透光的96孔板中加入20μl j774-dual
tm
细胞作用上清,用微孔板化学发光检测仪(lb 960)检测荧光素酶的表达。
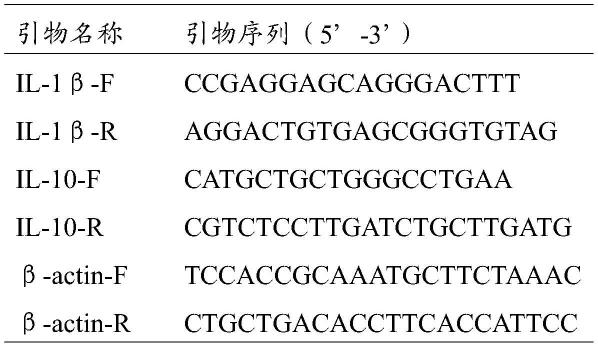
52.表1引物序列
[0053][0054]
实验结果:如图3a-d所示,为屎肠球菌c171对巨噬细胞抗炎和抗感染作用的结果。c171能显著下调急性炎症模型下hd11产生的炎症细胞因子il-1β,与l+n对照组相比差异极显著(p《0.0001;图3a)。c171能有效刺激hd11产生抗炎细胞因子il-10,与l+n对照组相比差异极显著(p《0.0001;图3b)。c171可以有效激活巨噬细胞j774-dual
tm
细胞的nf-κb信号通路,空白对照相比差异极显著(p《0.0001;图3c);c171可以有效激活巨噬细胞j774-dual
tm
细胞中的干扰素调节因子(irf)信号通路,与空白对照组相比差异极显著(p《0.0001;图3d)。结果说明,屎肠球菌c171具有强烈抗炎作用,同时具有抗感染作用。
[0055]
实施例4屎肠球菌c171对鸡原代巨噬细胞的抗炎和抗感染特性
[0056]
实验方法:
[0057]
一、首先分离鸡原代骨髓巨噬细胞,分离步骤如下:
①
安乐死处死3周龄spf鸡只,在无菌操作台中取鸡股骨;
②
将股骨两端剪断,用吸入pbs的注射器将骨髓冲出;
③
将骨髓收集至40μm滤网中,用10ml注射器塞研磨,然后用pbs冲下滤网上的骨髓;
④
混合物轻轻加至等体积histopaque-1083分离液上,2,000r/min离心25min,加速度调成最小;
⑤
吸取中间层细胞,pbs洗2遍;
⑥
将细胞计数后,接种至12孔板中,6h后换液,洗去不贴壁或半贴壁的细胞;
⑦
培养6d(每2d换一次培养液)后进行后续实验。
[0058]
二、使用cell counting kit-8,验证c171对细胞增殖活性的影响,以判断c171对细胞的毒性。鸡原代骨髓巨噬细胞按1
×
106cells/ml的浓度接种100μl细胞到96孔板中,加入50moi的c171悬液,孵育24h后,加入10μlcck-8溶液。在37℃孵育2h后,测定450nm处的吸光度。
[0059]
三、探究c171对鸡原代骨髓细胞是否具有抗炎作用。细胞接种于12孔后加入50moi益生菌悬液,37℃ 5%co2孵育24h,利用lps和nigericin使细胞发生急性炎症反应,建立急性炎症模型,即每孔加入工作浓度为1μg/ml的lps,37℃ 5%co2作用3h,再加入工作浓度为1μm的nigericin作用1h后,收集细胞沉淀用syby green
ꢀⅰ
荧光定量pcr根据表2的引物,来分析细胞中il-1β和il-10mrna的转录水平。以lps和nigericin处理的细胞作为阳性对照,以未作任何处理的细胞为空白对照。
[0060]
表2引物序列
[0061][0062][0063]
实验结果:图4为c171在原代巨噬细胞上的抗炎和抗感染作用的结果。c171对鸡原代骨髓巨噬细胞无毒性,并且c171能够刺激鸡原代骨髓细胞增值,c171与空白对照相比差异极显著(p《0.0001)。c171能显著下调急性炎症模型下鸡原代骨髓巨噬细胞产生的细胞因子il-1β(p《0.01),上调细胞因子il-10(p《0.001)和ifn-γ(p《0.0001)。结果说明:c171对鸡原代骨髓巨噬细胞同时具有抗炎和抗感染作用。
[0064]
实施例5屎肠球菌c171株的抗炎作用是通过调控nlrp3炎性小体通路,抑制caspase-1的活性来实现的。
[0065]
实验方法:
[0066]
一、使用cell counting kit-8,验证c171对hek293t细胞增殖活性的影响,以判断c171对细胞的毒性。hek293t细胞按1
×
106cells/ml的浓度接种100μl细胞到96孔板中,加入50moi的c171悬液,孵育24h后,加入10μl cck-8溶液。在37℃孵育2h后,测定450nm处的吸光度。
[0067]
二、检测c171的抗炎机制。将hek293t细胞以5
×
104/孔的浓度接种于48孔板中,当细胞密度达到70%时使用1μl的x-tremegene hpdnatransfection reagent瞬时转染100ng igluc,10ng pcaggs-caspase-1,10ng pcaggs-asc,12.5ng pcaggs-nlrp3,转染空质粒和单独转染igluc作为对照组.12h后加入moi为25、50、100的益生菌c171,再继续共培养12h后,收集细胞上清,3000rpm离心5min,去除细胞碎片。根据制造商的说明,用piercetm gaussia luciferase flashassay kit测定荧光素酶活性。细胞沉淀用于进行western blot相关蛋白(nlrp3、asc、caspase-1p20、pro-il-1β)的检测。
[0068]
实验结果:如图5所示,c171对hek 293t细胞无毒性(图5a),并且能够增加细胞的活性,且呈明显的剂量依赖性。由荧光素酶结果显示,c171能显著抑制il-1β的分泌,并且具有剂量依赖性,随着c171的增加,抑制作用越明显,与阳性对照组差异极显著(p《0.0001;图5b)。western blotting结果显示,c171不影响nlrp3、asc、pro-il-1β的表达,但能够降低caspase-1p20的表达,且呈剂量依赖性,灰度分析可知,25moi时,c171组是阳性对照的0.60倍(图5c);50moi时,是阳性对照的0.47倍(图5c);moi为100时,是阳性对照的0.29倍(图5c)。综合以上结果可知,c171通过抑制caspase-1的活性从而抑制il-1β的分泌,进而调控
nlrp3炎性小体通路。
[0069]
实施例6屎肠球菌c171株在体外的抗病毒增殖特性
[0070]
实验方法:dt40(1
×
106个/ml)以无血清的rpm 1640接种于12孔板中,37℃培养30min,细胞贴壁后,感染1moi vvibdv,同时加入moi为25、50、100的益生菌c171,三者作用24h后,收集200μl细胞悬液,使用premix ex taq采用绝对分析法定量测定ibdv vp5拷贝数(表3引物),细胞沉淀检测细胞因子il-1β,il-10,ifn-γ(表2引物),并通过westernblot检测vp2蛋白。
[0071]
表3引物序列
[0072][0073]
实验结果:如图6所示,c171不影响dt40的细胞活性,对dt40细胞安全无毒(图6a)。c171在体外有明显的抗病毒作用,与阳性对照相比c171能够明显降低vvibdv的病毒拷贝数,并且有明显的剂量依赖性,随着c171的剂量增加,抑制作用越明显。在益生菌与dt40的moi为100时,抑制作用最为显著(p《0.0001;图6b)。western blotting结果显示,c171能明显降低vvibdv vp2的蛋白表达,并且有明显的剂量依赖性。灰度分析可知,随着c171的剂量增加,vp2蛋白的表达逐渐降低,在益生菌与dt40的moi为100时,vp2的表达量最低,是阳性对照的0.43倍(图6c)。il-1β结果显示,c171能显著下调vvibdv感染模型下dt40细胞产生的细胞因子il-1β,并且具有计量依赖性,当moi为50和100时,与阳性对照组相比差异最显著(p《0.0001;图6d)。il-10结果表明,c171能有效刺激dt40细胞产生细胞因子il-10,当moi为100时,与阳性对照组相比差异极显著(p《0.0001;图6e)。ifn-γ结果显示,c171能有效刺激dt40细胞因子ifn-γ,并且具有剂量依赖性,当moi为100时,刺激作用最明显,与阳性对照组相比差异极显著(p《0.0001;图6f)。综合以上结果表明,c171在体外能降低病毒复制,降低炎性因子的表达,通过分泌ifn-γ来发挥抗感染作用。
[0074]
实施例7口服屎肠球菌c171株的可显著保护鸡超强毒力病毒感染引发的死亡,保护率达80%以上。
[0075]
实验方法:
[0076]
一、动物实验方案:spf鸡购买于中国农业科学院哈尔滨兽医研究所实验动物中心,饲养于负压过滤空气隔离器中。40只18日龄spf鸡随机分为三组:空白对照组(15只),口服c171组(15只),攻毒对照组(10只)。口服c171组:用pbs重悬c171后,每只鸡每日1ml,2
×
109cuf/ml,直接灌入口中,连续口服7天。七天后,口服c171组与空白对照组每组各杀5只
鸡。除空白对照组外,剩余的20只21日龄spf鸡,用中国vvibdv参考株hlj0504攻毒(vvibdv-hlj0504),攻毒方式为滴鼻点眼,攻毒剂量为5ld50/只,每只鸡200ul,之后每天观察发病情况。其中口服c171组,在攻毒后,再次连续口服7天c171。每天观察鸡发病情况,攻毒七天后,所有鸡全部剖杀,并进行剖检,计算存活率。
[0077]
二、测定各组鸡的法氏囊和体重,计算各组鸡的法氏囊指数(bbix),按照以下公式进行计算:bbix=试验组鸡的囊重比/空白对照组鸡的平均囊重比;囊重比=(法氏囊重/体重)
×
100%,bbix小于0.70的法氏囊被认为是萎缩。
[0078]
三、elisa检测口服后1周后血清中ifn-γ的含量。
[0079]
四、通过荧光定量的方法检测法氏囊中ibdv的拷贝数。
[0080]
实验结果:
[0081]
如图7所示,对鸡进行口服c171,然后用vvibdv-hlj0504感染spf鸡(图7a),在动物体内检测c171抗炎、抗感染作用。血清elisa结果表明,口服c171一周后,能诱导机体产生ifn-γ,与blank group相比差异极显著(p《0.0001;图7b)。攻毒结果表明,用vv-ibdv-hlj0504攻毒一周后,ibdv group spf鸡的存活率为20%,而c171+ibdv group spf鸡的存活率为80%,与ibdv group相比增加了4倍(图7c)。剖检结果显示,c171+ibdv group与ibdv group存活鸡的bbix全部小于0.7,两者无显著差异(p》0.05;图7d),两者极显著低于blank group(p《0.0001;图7d)。我们也检测了病毒在体内的复制能力,荧光定量结果显示,c171+ibdv group法氏囊中病毒拷贝数显著低于ibdv group(p《0.01;图7e)。综合以上结果表明,c171在体内能抑制病毒复制,降低死亡率。
[0082]
实施例8口服屎肠球菌c171株的显著抑制了超强毒力病毒vvibdv引发鸡的急性炎症。
[0083]
实验方法:
[0084]
一、动物实验方案同实施例7。取每组鸡的部分法氏囊,用10%中性缓冲福尔马林固定,苏木精和伊红染色,进行进一步的组织病理学检查。
[0085]
二、通过western blot检测法氏囊、脾脏、肠道中hmgb1蛋白的表达情况。
[0086]
三、elisa检测攻毒后1周后血清中il-1β、il-10的含量。
[0087]
四、通过荧光定量的方法检测法氏囊中il-1β、il-10的转录水平。
[0088]
实验结果:
[0089]
如图8所示,为探究口服c171后能否减轻ibdv对法氏囊的损伤,鸡法氏囊病理切片观察结果显示,c171+ibdv group(图8a-a)出现了组织病理学损伤,法氏囊中滤泡萎缩,广泛性淋巴细胞坏死、减少,间质增生;ibdv group(图8a-b)组织病理学损伤较c171+ibdvgroup严重,法氏囊滤泡严重萎缩,广泛性淋巴细胞坏死、减少,间质增生,滤泡中央形成隐窝,内有坏死细胞团块;blank group(图8a-c)与c171 group(图8a-d)均正常,未发现任何临床症状和显微病变(图8a)。为了验证c171能否减轻机体的急性炎症反应,检测了血清和组织中的炎性细胞因子和法氏囊、脾脏、肠道中hmgb1的表达情况。western blotting结果显示,ibdv group的hmgb1蛋白在法氏囊、脾脏、肠道中的表达量均高于c171+ibdv group组(图8b)。检测了血清中il-1β,il-10的含量,elisa结果表明,血清中il-1β的浓度c171+ibdv group极显著低于ibdv group(p《0.001;图8c),与blank group无显著差异(p>0.05;图8c);血清中il-10的浓度c171+ibdv group极显著高于ibdv group(p《0.0001;图
8d)和blank group(p《0.0001;图8d)。检测了法氏囊中il-1β,il-10的mrna的转录水平,荧光定量结果表明,口服c171后能下调法氏囊中il-1β的mrna转录水平,c171+ibdv group极显著低于ibdv group(p《0.001;图8e),与blank group无显著差异(p>0.05;图8e);口服c171后能上调法氏囊中il-10的mrna转录水平,c171+ibdv group与ibdv group,blank group相比差异显著(p<0.05;图8f)。综合以上结果表明,c171能抑制il-1β的产生,促进il-10的表达,降低了机体炎症反应。
[0090]
实施例9口服屎肠球菌c171株的可显著提高鸡在超强毒力病毒感染时的抗感染作用。
[0091]
实验方法:
[0092]
一、动物实验方案同实施例7。从新鲜抗凝血中分离外周血单个核细胞(pbmcs),检测其t细胞亚群。随机采集三只口服c1711周和攻毒后1周的抗凝血标本,使用鸡外周血淋巴细胞分离试剂盒分离得到pbmcs。将r-phycoerythrin/cyanine 5(sprd)标记的抗鸡cd3抗体、fluorescein isothiocyanate(fitc)标记的抗鸡cd4抗体和r-phycoerythrin(pe)标记的抗鸡cd8抗体稀释在含有5%fbs的pbs里,按照说明书提供的染料与细胞的浓度比例对pbmcs进行染色,在4℃摇床上染色30min后,将pbmcs用含有5%fbs的pbs清洗三次之后,用流式细胞分析仪分析cd3+淋巴细胞、cd4+淋巴细胞和cd8+淋巴细胞的比例。
[0093]
二、elisa检测攻毒后1周后血清中il-12、il-2、il-4、ifn-γ的含量。
[0094]
三、从新鲜抗凝血中分离外周血单个核细胞(pbmcs),检测其增殖活性。通过concanavalina(cona)and phorbol 12-myristate 13-acetate(pma)刺激的pbmcs来检测t细胞增殖活性。采用鸡外周血单个核细胞分离试剂盒(tbd,china)来分离的鸡外周血单个核细胞。用含有10%胎牛血清和青霉素-链霉素的rpmi 1640培养基稀释pbmcs至1
×
106cells/ml。96孔板每孔加入100μlpbmcs。然后每孔加入5μg/ml cona和100ng/mlpma,在37℃的细胞培养箱中培养48h。之后加入10μl cck-8,4h后测量在450nm处的吸光度。
[0095]
实验结果:如图9所示,探究了c171在体内的抗感染机制,通过流式细胞术分析pbmcs中t细胞亚型。流式结果表明,与blank group相比,口服c171一周后,cd8+t细胞的含量升高(图9a),c171 group的cd8+/cd4 t cell ratio为0.192,blank group的cd8+/cd4 t cell ratio为0.157(图9b);攻毒vv-ibdv-hlj0504一周后,ibdv group和c171+ibdvgroup的存活鸡cd8+t细胞含量均升高(图9a),ibdv group与c171+ibdv group cd8+/cd4 t cell ratio分别为0.259和0.546(图9b)。其中c171+ibdv group存活鸡cd8+/cd4+t细胞的比值最高,显著高于ibdv group(p《0.05;图9b);c171 group与blank group相比无显著差异(p>0.05;图9b)。因cd8+t cell能够诱导机体产生高水平的ifn-γ,cd4+t cell能够诱导机体产生高水平的il-4,所以我们又检测了血清中ifn-γ,il-4的含量。elisa结果表明,血清中ifn-γ的浓度c171+ibdv group极显著高于ibdv group(p《0.0001;图9c)和blank group(p《0.0001;图9c)。血清中il-4的浓度c171+ibdv group极显著低于ibdvgroup(p《0.0001;图9d),极显著高于blank group(p《0.0001;图9d)。这说明口服c171后能诱导机体产生高水平的cd8 t cell,促进ifn-γ的分泌,提高机体抗感染能力。为探究攻毒后各组t细胞增值活性的差异,我们将攻毒后一周的pbmcs用cona+pma刺激,检测各组鸡pbmcs中t细胞增值活性。结果显示,c171+ibdv group的pbmcs增值活性显著高于ibdv group(p《0.01;图9e)和blank group(p《0.0001;图9e)。细胞因子能刺激t细胞增值,又检测了血清中t细胞生长因
子il-12,il-2,il-23的含量。elisa结果显示,血清中il-12的浓度c171+ibdvgroup极显著高于ibdv group和blank group(p《0.0001;图9f)。血清中il-2的浓度c171+ibdv group极显著高于ibdv group和blank group(p《0.0001;图9g)。血清中il-23的浓度c171+ibdv group极显著高于ibdv group和blank group(p《0.0001;图9h),白细胞介素-23(interleukin-23;il-23),是由p19和il-12的40kd亚基组成,由活化的dc分泌。p19与il-6、g-csf和il-12的35kd亚基结构相似,能促进cd45ro+记忆t细胞增殖并产生γ干扰素。综合以上结果表明,口服c171可显著提高pbmcs中cd8 t cell的比例,促进机体分泌ifn-γ,提高t细胞的增殖活性,有利于机体提高抗感染能力。
[0096]
实施例10屎肠球菌c171株安全性评价。
[0097]
实验方法:鸡胚安全性检测,将3枚10日龄鸡胚从37℃孵化箱取出,蛋胚在照蛋器上照视,用铅笔画出气室与胚胎位置,并在绒毛尿囊膜血管较少的地方作记号。用碘酒消毒气室蛋壳,再用75%的酒精脱碘,并用钢针在记号处钻一小孔。用1ml注射器吸取培养12h菌液,针头刺入孔内,经绒毛尿囊膜入尿囊腔,注入0.1ml;用石蜡封孔后于37℃孵卵器孵育11天;气室接种:操作同上,接种部位为气室,共接种3枚10日龄鸡胚。
[0098]
毒力因子和耐药基因检测,取37℃静置培养6h的c171菌株200μl放入1.5ml离心管,取一个分离的菌落,12000g离心min,弃上清上。沉淀中加入200ul insta gene溶液,在56℃金属浴下培养15-30分钟。涡旋10s,放入100℃金属浴作用10min后,取出放入冰中,12000g离心5min,吸出200μl上层清夜到新离心管用于后续pcr检测。
[0099]
pcr反应体系50μl:premix taq酶(ex taq2.0 plus)25μl,上下游引物(10μmol/l)各1μl,模板1.5μl,ddh2o 21.5μl。pcr反应程序:95℃预变性5min;95℃变性1min,退火45s,72℃延伸1min30s,35个循环;72℃延伸10min。pcr产物经1.0%琼脂糖凝胶电泳检测。引物及退火温度见表4。
[0100]
表4毒力因子、耐药基因引物序列及退火温度
[0101][0102]
实验结果:如图10所示,c171组和空白对照组共6枚10日龄鸡胚均成功孵化(图10a)。以屎肠球菌c171 dna为模板,毒力因子基因和耐药基因引物进行pcr扩增,其毒力因子基因fsra有明亮条带,hyl、bopd、cylb、cyla、scm、cylm、spre、gele、asal未见目的条带(图10b);耐药基因tetm、mefa、tem、aac、ermb、vana均未见目的条带(图10c)。综上所述,屎肠球菌c171安全性较好。
[0103]
实施例11屎肠球菌c171株耐酸耐胆盐益生特性。
[0104]
实验方法:往新鲜的mrs培养液中加入1mol/lhcl调整培养液的ph值,将ph调整至3.0,4.0,5.0。往新鲜的mrs培养液中加入.0.4%,0.6%,0.8%,1.0%,2.0%,3.0%和4.0%(w/v)的牛胆盐。具体方法如下:在无菌96孔板中每孔加入180μl配制好的不同ph值的mrs培养液或含不同浓度胆盐的培养液,随后将20ul培养12h的屎肠球菌c171菌液接种于每个孔中,于37℃细菌培养箱中培养。使用酶标仪在630nm下记录每小时益生菌的生长状态。
[0105]
实验结果:屎肠球菌c171在胆盐质量分数为0.2%时增殖活性最好,随着胆盐质量分数的增加增殖活性逐渐下降,但在浓度为2.0%时仍有增殖。图11a显示益生菌c171株的耐胆盐能力较强;益生菌c171在ph值为3~6的mrs培养基中均能生长,od
630
nm值随着ph值降低而明显降低,但依旧可以生长,表明c171具有较强的耐酸性能(图11b)。综上所述,c171株能够耐受肠道胆盐的环境,并能顺利通过胃肠道的酸性环境并在胃肠道中生存。
[0106]
实施例12口服屎肠球菌c171株对spf鸡肠道菌群的影响。
[0107]
实验方法:同实施例7,口服屎肠球菌c171株一周后分别采集小肠内容物,每份样品至少3个重复,-80℃冰箱保存,干冰保温箱送上海欧易公司进行16s多样性分析。
[0108]
实验结果:根据群落结构柱状图分析及热图分析属水平top30物种菌群结果:口服c171后与对照组下相比,小肠肠道菌群结构没变,物种组成丰度改变。在前30菌属中,乳杆菌属、柠檬酸杆菌属、副拟杆菌属、栖粪杆菌属、肠球菌属及丁酸梭菌属等有益菌属丰度增多,丰度较高(图12a、12b);而伯克霍尔德氏菌属-卡巴罗勒尼亚属-帕拉伯克霍尔德属减少,魏斯氏菌属减少,泛菌属减少,sphingomonas减少,romboutsia丰度减少。空白对照组中伯克霍尔德氏菌属-卡巴罗勒尼亚属-帕拉伯克霍尔德属、romboutsia、溶杆菌属、sediminibacterium、大肠埃希氏志贺菌属、栖粪杆菌属、节线菌属暂定种丰度较高(图12a、12b)。
[0109]
根据群落结构柱状图和热图分析种水平top30物种,得出以下结果:口服c171后与对照组相比,小肠肠道菌群结构没变,物种组成丰度改变,植物乳杆菌、食窦魏斯氏菌、单形拟杆菌、耐久肠球菌、长双歧杆菌、约氏乳杆菌增加,而fungorum伯克霍尔德氏菌减少,食窦魏斯氏菌减少(图13a、13b)。空白对照组中成团泛菌、romboutsia_ilealis_g__romboutsia、假长双歧杆菌、大肠杆菌、肠乳杆菌、鼠李糖乳杆菌、罗伊氏乳杆菌、fungorum伯克霍尔德氏菌、jejuni普雷沃氏菌、螺杆菌属的某个种、普通拟杆菌、危险罗尔斯通氏菌、产酸拟杆菌、溶杆菌属的某个种、扭曲真杆菌、sediminibacterium sp.g sediminibacterium、杜雷拟杆菌丰度较高(图13a、13b)。
[0110]
口服屎肠球菌c171株对肠道菌群的小肠α多样性指数稀释曲线分析表明,小肠α多样性随稀释变为平缓,α多样性具有菌群代表性;小肠α多样性相关箱线图,口服c171多样性增加;小肠β多样性指数分析,组间存在差异(图14)。
[0111]
图15为口服屎肠球菌c171株对肠道菌群的小肠微生物多元变量lefse分析表明,该结果表明口服c171后乳杆菌、肠球菌等显著增加。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1