一种水稻油体蛋白基因OsOle6及其编码蛋白与应用
一种水稻油体蛋白基因osole6及其编码蛋白与应用
技术领域
1.本发明涉及植物基因工程技术领域,具体涉及一种控制水稻种子脂肪酸含量的基因序列,以及所述基因序列的应用。
背景技术:
2.淀粉和脂类是植物储藏器官中的两类重要能量储藏物质,其对于植物的生长发育和繁殖具有重要意义。根据植物储藏物的偏好,通常可以将植物的种子分为淀粉类和油料类两种。由于脂质不仅是能量分子,其还作为很多营养成分如维生素等溶剂而广泛存在于各类植物的种子种。在淀粉类植物种子中,尽管其含量相对较小,但是由于其能够与淀粉以及维生素等形成复合结构,而对种子的品质表现起到重要作用。从营养角度看,植物脂质是人类必须的营养物质,不仅能为人体提供热能和必需脂肪酸,还能促进脂溶性维生素吸收,同时,脂质中的不饱和脂肪酸由于具有抗氧化特性而对人体健康具有多方面的改善效果(suh et al.,plant cell report,2015,34:517-8)。
3.水稻是典型的以淀粉为主要储藏物的粮食作物,其可食用部分主要为胚乳,胚乳的主要成分是淀粉,其次是蛋白质和脂质。随着国民生活水平的提高,对稻米品质的要求也越来越高。众多的研究表明,通过对稻米胚乳淀粉组成的遗传调控可以有效提高稻米的食味品质,如适当降低稻米的直链淀粉含量能够显著提高稻米的食味品质(张昌泉等,遗传,2021,43:425-441)。但是当稻米直链淀粉含量较低时,稻米外观品质往往会收到影响,表现为透明度下降,稻米胚乳变暗。此外,研究表明,通过下调胚乳中蛋白质含量也能有效改良稻米食味品质,但是,在当前高氮肥栽培条件下这种方式实施难度较大(yang et al.,nature communications,2019,10:1949)。脂质作为稻米中的第三含量组分,其尽管占比很低,但是由于其能够与淀粉形成复合体而对淀粉的理化特性和稻米品质有重要影响(wang et al.,comprehensive reviews in food science and food safety,19:1056-1079)。因此,通过对稻米中的脂质含量和组成进行遗传调控来改良稻米品质成为可能。
4.植物种子中的油脂合成可以分为三个阶段:第一阶段是在质体中以蔗糖作为主要碳源合成脂肪酸,第二阶段是在内质网中合成三酰甘油,第三阶段是三酰甘油被包裹在单层磷脂分子膜内形成油体,并以出芽的方式进入细胞质。其中位于油体膜上的蛋白质被称为油体蛋白(oleosin),其能够防止油体之间相互融合而有利于油体的分散和稳定(xu et al.,engineering,2018,4:203-225)。在油料植物中的研究表明油体蛋白对植物的生长发育、抗逆性和营养器官脂质的积累都有重要影响。拟南芥中的研究表明油体蛋白能够与甾醇互作协调调油体的组装并影响脂质的含量(yu et al.,nature communication,2021,12:5598)。在水稻中过量表达来自大豆中的油体蛋白基因能够显著增加种子的脂肪酸含量(liu et al.,theoretical and applied genetics,2013,126:2289-2297)。这些结果表明在水稻中通过对油体蛋白基因的遗传调控有可能对稻米的脂肪酸含量进行调控并进而用于稻米品质改良应用。
5.水稻中存在多个油体蛋白基因家族成员,有关各成员的功能和效应研究还极少,
其对稻米品质的效应并不清楚。因此,克隆和评价油体蛋白基因对稻米脂质和品质的影响,将丰富水稻品质基因资源,并为优质水稻新种质的创建奠定基础。
技术实现要素:
6.为解决现有稻米品质改良存在的问题,本发明的目的在于提供水稻油体蛋白基因osole6及其编码蛋白在调控稻米脂肪酸含量和品质中的应用。
7.在水稻基因数据库(https://rapdb.dna.affrc.go.jp/)中基于油体蛋白(oleosin)信息查询获得油体蛋白基因序列信息,进一步通过基因表达数据库(https://ricexpro.dna.affrc.go.jp/)对各基因的表达特性进行分析,鉴定到一个水稻胚乳特异高表达油体蛋白基因osole6(os04g0546500)。
8.本发明的目的是通过以下技术方案来实现的,一种水稻油体蛋白基因osole6,其特征在于,所述油体蛋白基因osole6的核苷酸序列如seq id no.1所示,或者所述油体蛋白基因osole6的核苷酸序列与seq id no.1所示序列至少90%同源。seq id no.1序列如下:gtcagtccacttctcactagctcgtagacagtgctgcacgtgggttagctacttagctctttctctgcattgctggcttaattttgcagctgcaggtgtgctcgatccccatggctgatcagcacagaggagtgatcggcggcggcggctacggtgaccgtggtggccaggagcagcaggagaagcagcccttcatgatgacggctctgaagacggtgaccgcggcgacggccgggggctcgatgctggtgctgtccgggctgatcctggccggcaccgtcatcgcgctcacggtggccaccccggtgctggtcatcttcagccccgtgctggtgccggcggccatcgcgctggcgctcatggcggccgggttcgtcacctcgggcgggctcggcgtggccgcgctctccgtgttctcgtggatgtacaagtacctcaccgggaagcacccgccgggcgccgaccagctggaccacgccaaggcgaggctcgcgtccaaggcccgcgacatcaaggaggcggcgcagcaccgcatcgaccaggcgcaggcgtcttagggcggcctgccgcggcgaggcgccgcttctgctagctagctcgatcgggcggcgcacgggacggcgccggccggcgcgcgtgcgttcgccgtggccacgcccacgctggctactcctcttatctgctgagctctgtaaatgcatgcgtacatgtttaattaagcttggtcgcgtagtactctcctatgtaatatgtcccctagattatctaagaggcgatcgtgcatgtcgggcgcctggtacgtgtggtctggtggatgtcgtgtttatggtcatgatgaataaaatgcagtgttccttcg。
9.考虑到密码子的简并性以及不同物种密码子的偏爱性,可以根据需要使用适合特定物种表达的密码子。因而,本发明的水稻油体蛋白基因osole6还包括由seq id no.1所示核苷酸序列经取代、缺失和/或增加一个或几个核苷酸,得到编码上述蛋白的核苷酸序列,或者与seq id no.1所示序列至少90%同源性。
10.具体的,所述osole6编码的氨基酸序列如seq id no.2所示,或者所述osole6编码的氨基酸序列与seq id no.2所示序列至少90%同源。seq id no.2序列如下:metalaaspglnhisargglyvalileglyglyglyglytyrglyaspargglyglyglngluglnglnglulysglnprophemetmetthralaleulysthrvalthralaalathralaglyglysermetleuvalleuserglyleuileleualaglythrvalilealaleuthrvalalathrprovalleuvalilepheserprovalleuvalproalaalailealaleualaleumetalaalaglyphevalthrserglyglyleuglyvalalaalaleuservalphesertrpmettyrlystyrleuthrglylyshisproproglyalaaspglnleuasphisalalysalaargleualaserlysalaargaspilelysglualaalaglnhisargileaspglnalaglnalaser。
11.可根据本发明公开的氨基酸序列(seq id no.2),在不影响其活性的前提下,取代、缺失和/或增加一个或几个氨基酸,得到所述蛋白的突变序列或者与seq id no.2所示
序列至少90%同源性。
12.一种水稻油体蛋白基因osole6在培育不同脂肪酸含量水稻品系中的应用。
13.所述应用的方法如下:对水稻油体蛋白基因osole6进行编辑、敲除、修饰、抑制或过量表达,使得目标水稻品种中的osole6基因表达水平改变,进而获得不同脂肪酸含量的水稻品种。
14.进一步地,所述基因编辑过程中所用的载体为pc1300-cas9-ole6,所述pc1300-cas9-ole6包含基因osole6,载体系统为crispr/csa9,系统中包含中间载体sk-grna和最终载体pc1300-cas9。
15.优选的,所述载体pc1300-cas9-ole6的制备方法,所述的方法如下:将引物与aar i限制性内切酶切割后的线性中间载体sk-grna混合,并用t4 dna连接酶连接,得到质粒为sk-grna-ole6,再利用限制性内切酶kpn i和bgl ii切割sk-grna-ole6c质粒,回收550bp片段并与经过kpn i和bamh i双酶切过的pc1300-cas9载体混合,利用并用t4 dna连接酶连接。
16.所述引物序列如下:
17.序列名称序列序列编号primer15'ggcaggcggcggctacggtgaccg3'seq id no.3primer25'aaaccggtcaccgtagccgccgcc3'seq id no.4。
18.进一步地,所述基因过量表达过程中所用的过量表达载体为ubi-ole6,其特征在于,所述ubi-ole6包含基因osole6,载体为植物表达载体pcambla1300,其中包括来自水稻自身组成性高表达基因osubi的启动子。
19.优选的,所述过量表达载体ubi-ole6的制备方法,所述的方法如下:将经限制性内切酶bamhi和sali切割线性化的携带有osubi启动子的pcambla1300-ubi载体与包含osole6的pcr扩增产物进行混合,用clonexpress ultra one step cloning kit进行同源重组连接。
20.所述pcr扩增引物序列如下:
[0021][0022]
对水稻中油体蛋白编码基因osole6进行编辑,敲除、修饰、抑制或过量表达,使水稻中油体蛋白编码基因osole6如seq id no.1所示序列发生突变或过量表达,进而使得所述氨基酸序列如seq id no.2所示表达水平降低、增加或活性改变,进而获得不同脂肪酸含量的水稻品种。
[0023]
提供水稻油体蛋白基因osole6在水稻品质中的应用,根据调控需要,破坏osole6基因编码蛋白的生物学功能降低水稻种子脂肪酸含量并提高稻米的食味品质,或利用水稻
胚乳特异表达启动子gt1驱动osole6基因的表达提高稻米的脂肪酸含量并提高了稻米的营养的品质。
[0024]
本发明具有以下有益效果:(1)本发明发现水稻油体蛋白基因osole6及其编码蛋白通过调控水稻脂肪酸含量来影响稻米品质表现,具体为破坏osole6基因编码蛋白的生物学功能能够显著降低水稻种子脂肪酸含量并显著提高稻米的食味品质。因此,在培育优良食味水稻新品种领域具有重要的应用价值。
[0025]
(2)本发明利用水稻胚乳特异表达启动子gt1驱动osole6基因的表达后,能够显著提高稻米的脂肪酸含量进而提高了稻米的营养的品质。因此,在培育高油含量和高营养价值的水稻新品种培育领域具有重要应用价值。
附图说明
[0026]
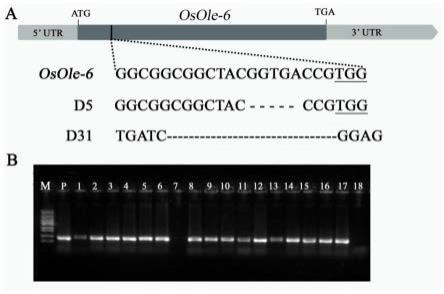
图1a为利用crispr/csa9载体系统所创建的osole6基因的两种突变类型d5和d31的具体突变信息。
[0027]
图1b为利用基因特异引物primer7和primer8进行的ubi-ole6水稻的pcr检测凝胶电泳图(注:b图中泳道从左到右分别为marker、阳性对照质粒、转基因植株(泳道1-17)和亲本(泳道18))。
[0028]
图2为osole6突变体稻米的脂肪酸含量和组分测定结果(注:a和b分别是米糠中脂肪酸含量和组分测定结果;c和d分别是精米中脂肪酸含量和组分测定结果)。
[0029]
图3为osole6突变体稻米的食味(a)和质构特性测定结果(b)。
[0030]
图4为osole6过量表达转基因稻米的脂肪酸含量(a)和脂肪酸组分(b)测定结果。
具体实施方式
[0031]
在本发明中所使用的术语,除非有另外说明,一般具有本领域普通技术人员通常理解的含义。
[0032]
下面结合具体的制备实施例和应用实施例,并参照数据进一步详细地描述本发明。应理解,这些实施例只是为了举例说明本发明,而非以任何方式限制本发明的范围,在不背离本发明的精神和范围的情况下,任何基于本发明技术方案的等效变换、简单替换等显而易见的改变,均落在本发明保护范围之内。
[0033]
在以下的实施例中,未详细描述的各种过程和方法是本领域中公知的常规方法。所用到的引物,均在首次出现时标明,其后所用相同引物,均以首次标明的内容相同。
[0034]
下述实施例中所用方法如无特别说明,均为常规方法。下述实施例中所用的试验材料,如无特殊说明,均为自常规生化试剂公司购买得到的。
[0035]
实施例1 crispr/cas9系统的水稻油体蛋白基因osole6编辑方法
[0036]
一、crispr/cas9载体构建
[0037]
(1)crispr引物序列如下:
[0038]
序列名称序列序列编号primer15'ggcaggcggcggctacggtgaccg3'seq id no.3primer25'aaaccggtcaccgtagccgccgcc3'seq id no.4。
[0039]
(2)载体构建及遗传转化
[0040]
本研究所使用的crispr/csa9载体系统由中国水稻研究所王克剑研究员提供,系统中包含中间载体sk-grna和最终载体pc1300-cas9,其dna骨架分别来源于pbluescript(sk+)载体和pcambla1300载体。
[0041]
具体步骤包括将上述引物primer1和primer2分别稀释到100μm浓度,各取10μl上述引物混合后100℃变性5分钟,随后室温冷却退火,形成带有粘性末端的片段。取7μl退火后的引物与用aar i限制性内切酶切割后的线性中间载体sk-grna(100ng)混合,并用t4 dna连接酶连接。随后,利用热激法将连接产物转化入大肠杆菌dh5α感受态细胞(南京诺唯赞公司)。将转化细胞涂布在含有100mg/l氨苄青霉素的lb固体培养基上进行培养,并挑选克隆进行测序,将测序正确的质粒命名为sk-grna-ole6。
[0042]
利用限制性内切酶kpn i和bgl ii切割sk-grna-ole6c质粒,回收550bp片段并与经过kpn i和bamh i双酶切过的pc1300-cas9载体混合,利用并用t4 dna连接酶连接。利用热激法将连接产物转化入大肠杆菌dh5α感受态细胞(南京诺唯赞公司;c502)。将转化细胞涂布在含有50mg/l卡那霉素的lb固体培养基上进行培养,并挑选克隆进行测序,将测序正确的质粒命名为pc1300-cas9-ole6。用电击法将pc1300-cas9-ole6转化农杆菌eha105菌株(擎科生物公司),将转化细胞涂布在含有50mg/l卡那霉素的lb固体培养基上进行培养,并挑选克隆进行测序,得到阳性菌株命名为cas9-ole6。
[0043]
将上述阳性cas9-ole6农杆菌菌株,利用农杆菌介导的水稻成熟胚转化方法(刘巧泉等,植物生理学报,1998)转化水稻中花11愈伤组织,通过潮霉素抗性筛选得到成功转入的愈伤组织细胞,再经组织培养和植物激素调节再分化为水稻幼苗,获得t0代水稻植株。
[0044]
实施例2 osole6过量表达载体构建与遗传转化方法
[0045]
一、osole6过量表达载体构建
[0046]
(1)osole6编码序列扩增引物序列如下:
[0047][0048]
(2)载体构建及遗传转化
[0049]
本研究所使用的植物表达载体为pcambla1300(购自上海联迈公司),其中包括来自水稻自身组成性高表达基因osubi的启动子。
[0050]
具体步骤包括用植物总rna提取试剂盒(dp441,天根公司)提取水稻中花11开花后15天的种子总rna,经微量核酸分析检测rna的质量后,利用逆转录试剂盒hiscript iii 1st strand cdna synthesis kit(诺唯赞公司),参照试剂盒说明,合成cdna。将上述引物primer3和primer4分别稀释到10μm浓度进行pcr扩增目的基因。
[0051]
利用高保真dna聚合酶phanta master(诺唯赞公司),按照以下组份顺序配制pcr反应液(50μl体系):2
×
phanta master mix(25μl),ddh2o(20μl),primer3(1.5μl),primer4(1.5μl和cdna(2μl)。随后在pcr仪上进行基因扩增,反应程序为:95℃变性1min;再
95℃10sec,55℃30sec,72℃45sec,共35个循环;然后72℃延伸5min;4℃保存。
[0052]
对pcr产物进行1%琼脂糖凝胶电泳检测,并切下含有目标基因片段的凝胶,用胶回收试剂盒(dp209天根)回收目的片段。随后将回收产物与经限制性内切酶bamhi和sali切割线性化的携带有osubi启动子的pcambla1300-ubi载体进行混合,用ultra one step cloning kit(诺唯赞公司)进行同源重组连接。利用热激法将连接产物转化入大肠杆菌dh5α感受态细胞(南京诺唯赞公司)。将转化细胞涂布在含有100mg/l氨苄青霉素的lb固体培养基上进行培养,并挑选克隆进行测序,将测序正确的质粒命名为ubi-ole6。
[0053]
将上述阳性ubi-ole6农杆菌菌株,利用农杆菌介导的水稻成熟胚转化方法(刘巧泉等,植物生理学报,1998)转化水稻日本晴愈伤组织,通过潮霉素抗性筛选得到成功转入的愈伤组织细胞,再经组织培养和植物激素调节再分化为水稻幼苗,获得t0代水稻植株。
[0054]
实施例3转基因材料分析
[0055]
(1)检测引物的设计
[0056]
对于基因编辑靶位点突变的检测,在靶位点基因组序列上下游60-300bp范围内设计测序引物primer5和primer6,用于检测靶点附近突变情况。
[0057]
测序引物序列如下:
[0058]
序列名称序列序列编号primer55'gcttcccggtgaggtacttg 3'seq id no.7primer65'accctcttctccctcctcac 3'seq id no.8。
[0059]
对于ubi-ole6水稻的检测,分别ubi启动子上设计引物primer7和ole6编码序列上设计primer8进行阳性转基因植株的鉴定。
[0060]
引物序列如下:
[0061]
序列名称序列序列编号primer75'agtgctagttattgaatca 3'seq id no.9primer85'agacgcctgcgcctggtc 3'seq id no.10。
[0062]
(2)转基因植株的鉴定
[0063]
对于crispr/cas9基因编辑水稻,利用ctab法提取t0代水稻叶片dna。利用引物primer5和primer6进行靶基因区段的常规pcr扩增,并经过1%琼脂糖凝胶电泳检测后,送南京擎科生物公司进行测序。测序结果利用在线工具包crisp-ge中的在线解码工具dsdecodem(http://skl.scau.edu.cn/dsdecode/)进行突变类型分析。收获具有突变类型的t0代水稻种子并再次种植,在t1代获得在靶位点缺失5个碱基和31个碱基的两种纯合突变类型d5和d31(如图3a所示),并继续种植,在t2代获得d5和d31两种纯合株系,用于后续分析。
[0064]
对于ubi-ole6过量表达转基因水稻,在利用ctab法提取t0代植株叶片dna后,利用引物primer7和primer8进行常规pcr扩增目的片段,并经过2%琼脂糖凝胶电泳检测,筛选能够扩增出目标片段的t0代阳性植株(如图1b所示),并进一步收获种植获得t1代阳性转基因苗。
[0065]
(3)转基因水稻性状分析
[0066]
对于crispr/cas9基因编辑水稻,在纯合突变d5和d31水稻的生长发育过程中进行肉眼观察,通过比较其与亲本对照在株高、分蘖数、生育期、穗长和粒形等性状,发现突变体与亲本对照无可见明显变化,说明osole6突变未对水稻的生长发育未产生明显影响。
[0067]
对于crispr/cas9基因编辑水稻,收获纯合突变d5和d31水稻的种子和亲本对照,利用砻谷机脱壳获得糙米,随后利用精米机出精米,收集每个样品的米糠和精米,送上海三黍生物公司进行脂肪酸组分和含量测定。具体为准确称取100mg样品,利用浓硫酸水浴法处理样品,并利用气相色谱分析agilent 7890a测定。气相色谱仪的工作条件为:fid氢火焰离子化检测器,hp-ffap 30m*0.25mm 0.25um色谱柱,进样口温度260℃,分流比20:1,检测器温度280℃,色谱柱初温150℃,程序升温10℃/min,升至210℃,并在此温度下维持8min,再程序升温20℃/min,升至230℃,并在此温度下维持6min。
[0068]
经数据分析,发现两个突变体的米糠和精米中的总脂肪酸含量较亲本对照均极显著下降(如图2a,c所示)。其中突变体米糠中的总脂肪酸含量下降幅度最大,降幅接近40%,而突变体精米中的总脂肪酸含量下降幅度相对较小,但也达到极显著差异。进一步分析了两个组织中主要脂肪酸的含量,结果表明,米糠和精米中的主要脂肪酸如棕榈酸、硬酯酸、油酸和亚油酸的含量较亲本对照均极显著下降,且米糠中各组分下降幅度相对较高(如图2b,d所示)。
[0069]
对于crispr/cas9基因编辑水稻,收获成熟的种子,脱壳、出糙和出精,获得精米。利用日本佐竹公司生产的米饭食味计(型号sta1a),进行米饭食味测定。主要操作步骤为:称取整精米30.0g于配套铝制容器中,使用配套淘米用具在清水下清洗30sec;擦干容器外层水分。在天平上补加蒸馏水至70g(精米:水=1:1.33),用同一个电饭锅蒸煮35min后,室温冷却平衡20min;称取10.0g冷却好的米饭,装入仪器配套金属环内,正反面各10s压平;在利用黑、白标准样板校准设备后,进行食味测定,每个样品重复3次,计算平均值。测定参数包含食味值、外观、硬度、粘度和平衡度(硬度粘度比)。
[0070]
对于crispr/cas9基因编辑水稻,在进行上述食味测定的同时,用ta-xt型质构仪测定米饭的质构特性。具体步骤为:每个样品随机挑选3粒完整米,按照夹角约120
°
将三粒米摆置在测样板上,测试采用tpa.p/36r探头,压缩程度为50%,测试前速率4.0mm/s,测试速率4.0mm/s,测试后速率4.0mm/s。利用系统自带软件(texture expert software program,version 5.16)分析测量参数:硬度和粘附性。每个样重测以3粒米为一组,重复测量6次,取平均值。
[0071]
经数据分析,发现两个突变体的米饭食味值极显著高于亲本对照,尤其是突变类型d31稻米的食味值最高(如图3a所示)。此外,突变体米饭的外观、黏度和平衡度均显著高于亲本对照,而硬度则显著低于亲本对照(如图3a所示)。这些指标进一步说明osole6突变后,米饭食味得到很好的改善。为进一步明确上述食味分析结果,利用米饭质构仪对米饭的质构特性做了进一步分析,结果表明突变体米饭的硬度极显著下降而粘附性极显著增加(如图3b所示),这再次表明osole6突变后,米饭的食味特性有了显著提升。
[0072]
对于ubi-ole6水稻,在t1代阳性植株进行种植过程中,通过肉眼观察,发现ubi-ole6水稻在生长发育过程中,其株高、分蘖数、生育期、穗长和粒形等性状与亲本对照无明显差异,说明osole6过量表达未对水稻的生长发育未产生明显影响。随后,挑选了t1代植株
的2个阳性单株ubi-ole6-1#和ubi-ole6-2#进行初步的脂肪酸含量和组分分析。
[0073]
对于ubi-ole6水稻,经数据分析,发现osole6过量表达植株ubi-ole6-1#和ubi-ole6-2#糙米的总脂肪酸含量较亲本对照极显著增加(如图4a所示)。从主要脂肪酸组分看,糙米中的棕榈酸、硬酯酸、油酸和亚油酸的含量均较亲本显著增加(如图4b所示)。这些结果说明通过过量表达osole6基因能够有效提高稻米的脂肪酸含量,进而增加稻米的营养品质。
[0074]
最后应说明的是:以上所述的各实施例仅用于说明本发明的技术方案,而非对其限制;尽管参照前述实施例对本发明进行了详细的说明,本领域的普通技术人员应当理解:其依然可以对前述实施例所记载的技术方案进行修改,或者对其中部分或全部技术特征进行等同替换;而这些修改或替换,并不使相应技术方案的本质脱离本发明各实施例技术方案的范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1