玉米双调控模块在调控植物生长和抗病平衡中的应用
1.本发明属于农业生物技术领域,具体涉及一种玉米双调控模块在调控植物 生长和抗病平衡中的应用,更具体是涉及一种tbf1基因启动子和uorf元件在 调控植物生长和抗病平衡中的应用。
背景技术:
2.玉米作为我国主要的粮食和饲料作物之一,在农业生产中占有重要地位。 近年来,伴随着气候变化和品种更替,玉米病害的发生呈现出持续加重的趋势, 严重威胁我国的粮食安全。因此,品种的抗病性改良,成为目前玉米育种工作 的重要目标。进入21世纪,植物抗病的分子机制研究有了突破性进展,建立了 植物免疫系统的基本框架,为玉米抗病分子改良奠定了良好的理论基础。然而, 生产实践和实验研究发现,植物生长和抗病之间存在紧密的拮抗关系,导入抗 病基因是提高玉米抗病性的有效手段,但是抗病基因过量表达时常常引起不可 预期的生长缺陷,比如抗病基因在改良作物抗性的同时,往往会抑制植物的生 长,进而影响作物产量。因此,生长和抗病的平衡问题,成为制约抗病基因在 玉米育种应用中的瓶颈。
3.在长期的进化和适应过程中,植物形成了复杂的调控网络和分子机制,用 来平衡自身的生长发育和抗病性。其中,拟南芥tbf1基因作为关键的开关分子, 在植物的生长和抗病转换过程中发挥重要作用。
4.作为生长和抗病转换的关键开关分子,tbf1的表达受到上游开放阅读框 (uorf,upstream open reading frame)在翻译水平的精确调控。研究表明, tbf1基因起始密码子上游区域含有2个富含苯丙氨酸密码子的uorf
tbf1
。当正 常生长时,适量的苯丙氨酸能够满足正常的代谢需求,此时,未磷酸化的翻译 起始因子eif2α会结合在uorf
tbf1
处,抑制tbf1的翻译起始;而在病原菌入 侵时,植物对苯丙氨酸的需求增加,导致大量空载苯丙氨酸转运核糖体(trna
phe
) 的积累和随后eif2α的磷酸化修饰促进其结合到tbf1的翻译起始位点,激活 tbf1表达的同时促进植物转换到抗病模式。因此,uorf
tbf1
的存在使tbf1的 表达在正常生长的拟南芥中被严格抑制,而在病原菌入侵时可以快速启动,从 而发挥其生长和抗病转换的开关作用(pajerowska-mukhtar,k.m.,wang,w.,tada, y.,et al.(2012)the hsf-like transcription factor tbf1 is a major molecular switchfor plant growth-to-defense transition.curr biol 22,103-112)。
5.基于tbf1基因转录和翻译水平上受到植物双重调控的分子机制,董欣年教 授和王石平教授团队在拟南芥和水稻中,成功克服了抗病基因表达引起的生长 缺陷(xu,g.,yuan,m.,ai,c.,et al.(2017)uorf-mediated translation allowsengineered plant disease resistance without fitness costs.nature 545,491-494.)。然 而,这种机制在其他作物中的保守性,尤其是在玉米对抗其特有病害中的作用 目前仍不清楚。
6.综上,现有技术存在的问题是,玉米抗病基因在发挥抗病性作用的同时可 对产量造成影响。虽然在拟南芥和水稻中证明tbf1基因调控模块能平衡植物的 生长和抗病性,但
玉米中tbf1基因的功能尚不清楚,尚未有通过利用玉米tbf1 启动子和uorf双调控模块调控玉米生长和抗病平衡的报道。
技术实现要素:
7.为了解决上述技术问题,本发明提供了一种玉米双调控模块在调控植物生 长和抗病平衡的应用。
8.本发明的目的是提供一种玉米双调控模块在调控植物生长和抗病平衡中的 应用,所述玉米双调控模块包括zmtbf1a基因的启动子和uorf序列,即 pzmtbf1a:uorf
zmtbf1a
(调控zmtbf1a基因表达),其核苷酸序列参见seq idno.1,其中的1-1565位为启动子,倒数第190-315位zmtbf1a基因的uorf; 所述玉米双调控模块还包括zmtbf1b基因的启动子和uorf序列,即 pzmtbf1b:uorf
zmtbf1b
(调控zmtbf1b基因表达),其核苷酸序列参见seq idno.2,其中1-1576位为启动子,倒数第155-280位为zmtbf1b基因的uorf。 所述tbf1基因包括zmtbf1a和zmtbf1b,zmtbf1a核苷酸序列参见seq idno.3,其中第1-234位、倒数1-663位为外显子,zmtbf1b核苷酸序列参见seqid no.4,其中1-228、倒数1-669为外显子。
9.优选的,上述玉米双调控模块在调控植物生长和抗病平衡中的应用,所述 玉米双调控模块在正常生长条件下抑制相应基因表达,在病害条件下显著诱导 相应基因表达,保证其各自调控的基因只在病害条件下表达,进而平衡植物的 生长和抗病,使植物既具有抗病性,又具有良好的生长特性。
10.优选的,上述玉米双调控模块在调控植物生长和抗病平衡中的应用,所述 植物为玉米。
11.优选的,上述玉米双调控模块在调控植物生长和抗病平衡中的应用,所述 玉米双调控模块(tbf1基因启动子和uorf)用于调控玉米性状,使玉米表现 为抗病稳产;所述抗病是指抗病虫害。
12.优选的,上述玉米双调控模块在调控植物生长和抗病平衡中的应用,所述 抗病虫害是由多堆柄锈菌、新月弯孢菌、禾谷镰刀菌、串珠镰刀菌、凸脐蠕孢 菌、禾生炭疽菌、伊朗花叶病毒、玉米青虫、粘虫或蚜虫引起的。
13.优选的,上述玉米双调控模块在调控植物生长和抗病平衡中的应用,从玉 米dna中扩增uorf
zmtbf1
元件和对应的zmtbf1启动子序列,利用无缝克隆的 方法分别克隆到植物表达载体pcambia3301中。
14.优选的,上述玉米双调控模块在调控植物生长和抗病平衡中的应用,所述 扩增uorf
zmtbf1
元件和对应的zmtbf1启动子序列的引物参见表5。
15.优选的,玉米双调控模块在调控植物生长和抗病平衡中的应用,利用ecori 和ncoi限制性内切酶线性化pcambia3301载体,利用无缝克隆技术将扩增的 玉米双调控模块序列连接到该植物表达载体中;所述玉米双调控模块包括 pzmtbf1a:uorf
zmtbf1a
和pzmtbf1b:uorf
zmtbf1b
。
16.优选的,玉米双调控模块在调控植物生长和抗病平衡中的应用,利用ncoi 和pmli对所构建载体进行双酶切,酶切位点之间用抗病基因(抗病基因是有所 述玉米双调控模块对应调控表达的)的序列替换载体中的gus序列,从而制备 出含玉米双调控模块以及抗病基因的植物表达载体。
17.与现有技术相比,本发明具有以下有益效果:
18.1、本发明鉴定了玉米中tbf1的两个同源基因(zmtbf1a和zmtbf1b), 发现其上游均存在保守的,且能够调控基因翻译起始的uorf
zmtbf1
元件。进一 步的研究结果表明,zmtbf1受到多种玉米病原菌和虫害的诱导,证明其启动子 具有病原诱导的特性;而且该基因的过表达能够激活超敏反应和促进免疫应答 相关基因zmbip2的表达。表明zmtbf1参与玉米的抗病反应,并且具有调控植 物生长和抗病转换的潜力。在此基础上,本发明克隆了zmtbf1a和zmtbf1b 基因上游的启动子和uorf调控元件,组成适用于玉米的病原诱导型基因调控 模块;构建了两个基于pcambia3301的植物表达载体,用于调控抗病基因在 玉米中的特异诱导表达,进而培育高抗和稳产的转基因玉米新品种。
19.2、本发明设计的两个病原诱导型调控模块可以平衡抗病基因在玉米应用中 对生长和抗病的影响,进而培育产量高、抗病性强的转基因玉米新品种。未来, 还可以利用基因编辑技术,将本发明设计的调控模块导入玉米抗病基因的上游, 实现无转基因标记的玉米高产抗病新品种。
附图说明
20.图1为拟南芥tbf1的蛋白序列在玉米基因组中的blast结果;
21.图2为拟南芥hsf家族基因和玉米中tbf1候选同源基因蛋白序列的进化 分析;进化树采用最大似然法构建;
22.图3为拟南芥和玉米tbf1基因蛋白、启动子和5
’‑
utr序列的比较;
23.a,拟南芥和玉米tbf1蛋白的氨基酸序列比对;b,拟南芥和玉米tbf1 基因的启动子序列比对;c,拟南芥和玉米tbf1基因的5
’‑
utr序列比对;
24.图4为uorf
tbf1
的序列和进化分析;
25.拟南芥和玉米tbf1基因uorf的dna(a)和氨基酸(b)序列比对;c, 不同物种uorf2
tbf1
同源序列比对;d,不同物种uorf2
tbf1
同源序列的进化分 析;at,arabidopsis thaliana拟南芥,pv,phaseolus vulgaris四季豆,gm,glycine max大豆,gr,gossypium raimondii棉花,nb,nicotiana benthamiana本式烟, ca,cicer arietinum鹰嘴豆,pd,phoenix dactylifera海枣,ma,musa acuminata subsp.malaccensis香蕉,os,o.sativa水稻,zm,zea mays l.玉米;
26.图5为uorf
zmtbf1
对下游基因翻译水平的抑制作用;
27.a,pzmtbf1:uorf
zmtbf1
驱动luc报告基因的载体示意图;b,uorf
zmtbf1a
对下游基因蛋白水平的影响;c,uorf
zmtbf1a
对下游基因转录水平的影响;d, uorf
zmtbf1b
对下游基因蛋白水平的影响;e,uorf
zmtbf1b
对下游基因转录水平 的影响;图5b-e中,i、ii、iii、iv分别对应图5a中的载体;
28.图6为本发明zmtbf1基因的表达分析;
29.a,zmtbf1a基因的组织表达特性;b,zmtbf1b基因的组织表达特性; c,zmtbf1a基因上调表达的处理;d,zmtbf1b基因上调表达的处理; e,zmtbf1a基因上调表达的突变体;f,zmtbf1b基因上调表达的突变体; 图6中,f.graminearum,禾谷镰刀菌;f.verticillioides,串珠镰刀菌; mimv,伊朗花叶病毒;c.graminicola,禾生炭疽菌;e.turcium,大斑病凸脐 蠕孢菌;sa,水杨酸;s.exigua,玉米青虫;m.separata,玉米粘虫;
aphid, 蚜虫;t.urticae,二斑叶螨;
30.图7为zmtbf1受到玉米病害的快速诱导;
31.a,zmtbf1a和zmtbf1b基因受到玉米南方锈病致病菌的诱导;b, zmtbf1a和zmtbf1b基因受到玉米弯孢叶斑病致病菌的诱导;c,zmpr1基因 在玉米接种新月弯孢菌后的表达模式;d,zmtbf1a基因在玉米接种新月弯孢 菌后的表达模式;e,zmtbf1b基因在玉米接种新月弯孢菌后的表达模式。p. polysora,多堆柄锈菌;c.lunata,新月弯孢菌;
32.图8为zmtbf1对免疫应答的调控作用;
33.a,zmtbf1a和zmtbf1b基因过表达诱导超敏反应;b,zmtbf1过表达 和zmbip2启动子驱动luc报告基因的载体示意图;c,zmtbf1诱导zmbip2 启动子驱动的luc报告基因上调表达;
34.图8c中i、ii、iii、iv分别对应图8b中的载体,“+”表示共转化了两个 载体;
35.图9为含有zmtbf1a基因启动子和uorf
zmtbf1a
元件(a)以及含有zmtbf1b 基因启动子和uorf
zmtbf1b
元件(b)的植物表达载体结构图;
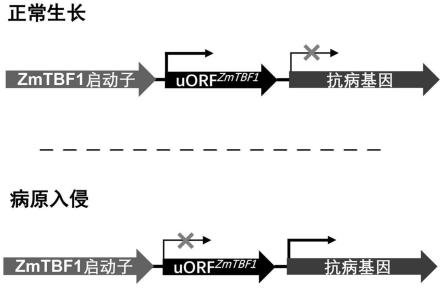
36.图10为基于zmtbf1基因设计的两个病原诱导型植物表达载体的工作模式 图。
具体实施方式
37.为了使本领域技术人员更好地理解本发明的技术方案能予以实施,下面结 合具体实施例和附图对本发明作进一步说明。
38.在本发明的描述中,如未特殊说明,所用试剂、载体均为市售,所用方法 (比如载体的构建方法、pcr扩增方法等)均采用本领域常规技术。
39.一、玉米tbf1同源基因的鉴定和序列分析
40.拟南芥tbf1/hsf4基因属于hsf热激蛋白家族。但是,功能上的研究表明, attbf1并没有参与植物的热激反应。本发明首先利用gramene(https://www. gramene.org/)网站的blast工具,检索了attbf1的蛋白序列,发现玉米中存 在较多与attbf1蛋白类似的序列,但是一致性相对不高(图1)。因此,本发 明进一步构建了这些基因和拟南芥hsf家族基因的系统进化树(图2)。结果表 明,有两个候选基因(zm00001eb314890和zm00001eb100770)同拟南芥tbf1 基因的进化关系非常近,极有可能是其功能类似的同源基因,因此分别命名为 zmtbf1a和zmtbf1b(图2)。
41.通过进一步的蛋白序列比对发现,zmtbf1a和zmtbf1b蛋白的氨基酸序 列一致性非常高,达到83.61%,但是与拟南芥tbf1蛋白的序列存在较大差异, 一致性分别只有38.87%和37.42%(图3a)。启动子作为基因表达的重要调控区 域,对基因的功能发挥非常重要。attbf1、zmtbf1a和zmtbf1b基因的启动 子均存在较大差异,一致性在35%左右(图3b)。但是zmtbf1a和zmtbf1b 基因的5
’‑
utr区域存在一段一致性非常高的区段(图3c)。
42.二、玉米中uorf
zmtbf1
元件能够调控下游基因的翻译起始
43.通过序列分析发现,zmtbf1a和zmtbf1b在5
’‑
utr区域高度一致的序 列中含有一个与uorf2
attbf1
序列类似的开放阅读框(图4a和4b)。拟南芥中 的研究表明,tbf1的功能发挥依赖于其基因上游的两个富含苯丙氨酸的开放阅 读框(uorf1
tbf1
和uorf2
tbf1
),但玉米zmtbf1基因5
’‑
utr区域存在的uorf 元件的功能尚不清楚。在不同物种中的检索结果表明,uorf2
tbf1
是一个广泛存 在的保守开放阅读框(图4c)。目前还不清楚uorf2
tbf1
在进化过程
中,是否因 为序列的变异产生功能上的多样性。进化分析表明,该元件在玉米和水稻中的 关系较近(图4d)。
44.已有的研究表明,uorf的一个重要功能就是调控下游基因的翻译水平。因 此,本发明从玉米b73自交系的dna中扩增了两个uorf
zmtbf1
元件和对应的 zmtbf1启动子序列,利用无缝克隆的方法(采用诺唯赞生物技术公司的 clonexpress ultra one step cloning kit产品完成)分别克隆到双荧光素酶报告系 统的载体中(图5a)。同时,通过在引物中引入突变碱基(atg突变为ctg) 的方法破坏了uorf
zmtbf1
的功能(uorf
zmtbf1
图5a)。上述载体构建的引物见表 1。通过peg介导的方法将这些载体瞬时转化到玉米原生质体中,并测定荧光素 酶的活性。结果发现,两个uorf
zmtbf1
突变后下游fluc的转录水平表达量没 有显著差异,但是相对酶活即蛋白水平的表达量明显升高(图5b-e)。这些结果 证明玉米中的这两个uorf
zmtbf1
对下游基因的翻译有抑制作用。基因表达检测 的引物见表2。
45.表1 uorf
zmtbf1
功能验证的载体构建引物
[0046][0047][0048]
表2 uorf
zmtbf1
功能验证的载体构建引物
[0049][0050]
三、zmtbf1基因受到多种玉米病原菌的诱导
[0051]
拟南芥tbf1基因作为生长和抗病转换的开关因子,其转录和翻译水平都受 到了严格的调控。借助pprd(http://ipf.sustech.edu.cn/pub/plantrna/)网站,本 研究对zmtbf1a和zmtbf1b基因在公共数据库中的表达水平进行了分析。结 果表明,zmtbf1基因的表达量在玉米的根、叶和雄蕊等组织中维持在一个相当 低的水平(图6a和6b)。而在玉米病原菌(如穗腐病和茎腐病的致病菌禾谷镰 刀菌和串珠镰刀菌、大斑病的致病菌凸脐蠕孢菌和炭疽病的致病菌禾生炭疽菌 等)处理时,两个基因的表达显著升高(图6c和6d)。同拟南芥中的结果类似, zmtbf1的表达量在sa处理下也受到明显诱导(图6c和6d)。另外,伊朗花 叶病毒和一些玉米常见虫害如玉米青虫、粘虫和蚜虫等也可以诱导zmtbf1的 基因表达,提
示zmtbf1广泛参与到玉米的病虫害应答反应中,同时表明该基 因的启动子具有病原诱导表达的特性(图6c和6d)。
[0052]
rp1-d21突变体是一个r基因重组导致的类病斑突变体,表现出广谱的锈 病抗性。在公共数据库的转录组数据中,两个zmtbf1基因在rp1-d21突变体 中的表达均量显著升高(图6e和6f)。本课题组前期收集了多份类病斑突变体 (les),对其中表现出抗病性提高的材料进行了转录组测序。结果表明,zmtbf1 基因在这些les突变体中的表达量也显著增高(图6e)。以上结果暗示zmtbf1 基因的启动子受到病原入侵的调控,导致基因的表达量升高。
[0053]
为了进一步分析该基因同玉米病害的关系,本发明利用qrt-pcr的方法, 检测了zmtbf1基因在玉米南方锈病的致病菌多堆柄锈菌和弯孢叶斑病的致病 菌新月弯孢菌接种后的表达量,所用引物见表3。结果表明,两种病菌的接种 均显著诱导了zmtbf1a和zmtbf1b的表达(图7a和7b)。此外,本发明对 玉米接种新月弯孢菌后,zmtbf1基因和病程相关基因zmpr1的表达变化进行 了为期一周的监测。结果发现,zmpr1基因的表达量在接种病菌4天后开始上 调,第6天达到高峰,而zmtbf1a和zmtbf1b基因的表达量在第4天就达到 峰值,随后快速恢复到接种前的水平(图7c-e)。因此,zmtbf1基因对于病 害的响应十分灵敏。
[0054]
表3 zmtbf1和zmpr1基因的定量引物
[0055][0056]
四、zmtbf1激活超敏反应,促进免疫应答相关基因zmbip2的表达
[0057]
植物激活免疫系统后常常会引发超敏反应,导致细胞死亡以对抗病原菌的 入侵。为了验证玉米zmtbf1的功能,本发明构建了该基因组成型过表达的载 体(图8b),通过农杆菌介导的方法瞬时转化到烟草中。结果表明,zmtbf1a 和zmtbf1b的过表达均导致了明显的超敏反应表型(图8a),表明zmtbf1基 因对免疫应答反应有激活作用。已有的研究表明,拟南芥tbf1蛋白可以结合到 bip2基因启动子的tl1元件上,参与植物的免疫应答过程。序列分析发现,玉 米bip2同源基因的启动子区域也含有一个典型的tl1元件。因此,本发明构建 了zmbip2启动子驱动luc的报告系统载体,并将其与zmtbf1的过表达载体 共转化到玉米原生质体中(图8b和8c)。相关载体构建引物见表4。由于zmtbf1 的过表达会引起强烈的超敏反应,因此对报告基因luc的酶活测定在转化后8 小时即进行。结果表明,zmtbf1a和zmtbf1b均促进了zmbip2启动子驱动的 luc基因表达(图8c)。
[0058]
表4 zmtbf1过表达和zmbip2启动子报告载体构建的引物
[0059][0060]
五、zmtbf1基因启动子和uorf的克隆和植物表达载体构建
[0061]
上述实验结果表明,两个zmtbf1基因的启动子和uorf
zmtbf1
元件组成的 调控模块能够灵敏的响应玉米的植物病害(如南方锈病和弯孢叶斑病),在平衡 玉米生长和抗病方面表现出较高的应用潜力。因此,本发明将两个zmtbf1启 动子和uorf
zmtbf1
序列组成的调控模块克隆到植物表达载体pcambia3301中 (图9-10)。载体构建引物见表5。后续应用过程中,利用ncoi和pmli进行双 酶切,酶切位点之间用抗病基因的序列替换载体中的gus序列,从而构建含有 病原诱导模块的抗病基因植物表达载体。该病原诱导型调控模块的工作模式见 图10。
[0062]
表5 zmtbf1过表达和zmbip2启动子报告载体构建的引物
[0063][0064][0065]
需要说明的是,本发明中涉及数值范围时,应理解为每个数值范围的两个 端点以及两个端点之间任何一个数值均可选用,由于采用的步骤方法与实施例 相同,为了防止赘述,本发明描述了优选的实施例。尽管已描述了本发明的优 选实施例,但本领域内的技术人员一旦得知了基本创造性概念,则可对这些实 施例做出另外的变更和修改。所以,所附权利要求意欲解释为包括优选实施例 以及落入本发明范围的所有变更和修改。
[0066]
显然,本领域的技术人员可以对本发明进行各种改动和变型而不脱离本发 明的精神和范围。这样,倘若本发明的这些修改和变型属于本发明权利要求及 其等同技术的范围之内,则本发明也意图包含这些改动和变型在内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1