特异调控禾谷镰刀菌子囊孢子成熟的基因FgTBC1及其应用
特异调控禾谷镰刀菌子囊孢子成熟的基因fgtbc1及其应用
技术领域
1.本发明属于微生物学领域,具体涉及一个特异调控禾谷镰刀菌子囊孢子成熟的基因fgtbc1及其应用。
背景技术:
2.禾谷镰刀菌是一种同宗配合的植物病原丝状子囊真菌,主要侵染小麦、大麦、玉米等禾谷类经济作物,引起小麦赤霉病和玉米茎腐病,降低粮食作物的产量及品质,并通过有性生殖在小麦麦穗、秸秆处产生大量子囊壳(stack r, leonard k, bushnell w. 2003 history of fusarium head blight with emphasis on north america. fusarium head blight of wheat and barley: 1-34.)。子囊壳是禾谷镰刀菌在田间越冬的重要形式,在小麦扬花期阶段,子囊壳喷射产生成熟的子囊孢子,随风、雨水或昆虫传播至小麦上,作为初级侵染源侵染小麦穗部,引起小麦赤霉病(schmale dg, arntsen qa, bergstrom gc. 2005. the forcible discharge distance of ascospores of gibberelia zeae. can j plant pathol 27: 376
–
382; trail f, gaffoor i, vogel s (2005) ejection mechanics and trajectory of the ascospores of gibberella zeae (anamorph fuarium graminearum). fungal genet biol 42: 528-533.)。
3.上述文献表明,子囊孢子在禾谷镰刀菌侵染循环中起着至关重要的作用。因此,鉴定禾谷镰刀菌中特异调控有性生殖过程中子囊孢子发育相关基因及其突变体,不仅能够揭示有性生殖过程子囊孢子的形成机理,而且能为小麦赤霉病的绿色生态防治和传播提供科学的策略。ph-1菌株是禾谷镰刀菌第一个全基因组测序的菌株,被广泛应用于禾谷镰刀菌遗传转化与基因功能研究当中。
4.在前期研究当中,我们对禾谷镰刀菌野生型ph-1菌株中12个含有tbc(tre-2/bub2/cdc16)结构域的基因进行基因敲除,发现了一个不影响生长及子囊壳产生,却特异阻断子囊壳中子囊孢子的成熟及喷射的突变体,我们将其命名为δfgtbc1,对应的基因命名为fgtbc1。因此,本发明公开了一个特异调控禾谷镰刀菌子囊孢子成熟的基因fgtbc1基因及其敲除突变体和应用。
技术实现要素:
5.本发明的目的在于提供一个特异调控禾谷镰刀菌子囊孢子成熟的基因fgtbc1及其应用,fgtbc1基因不影响禾谷镰刀菌正常的营养菌丝生长、分生孢子形成及子囊壳产生,但特异调控了禾谷镰刀菌有性生殖过程中子囊孢子的成熟和喷射,因此fgtbc1基因对禾谷镰刀菌的传播和绿色生态防治具有重要的意义,也是研究丝状子囊真菌有性生殖过程作用机制的重要基因资源。
6.为实现上述目的,本发明采用如下技术方案:一个特异调控禾谷镰刀菌子囊孢子成熟的基因fgtbc1,所述基因fgtbc1的核苷酸序列如seq id no.1所示。
7.进一步的,上述fgtbc1基因编码的蛋白的氨基酸序列如seq id no.2所示。
8.一种禾谷镰刀菌fgtbc1基因敲除突变体δfgtbc1,所述禾谷镰刀菌fgtbc1基因敲除突变体δfgtbc1的制备步骤如下:利用引物fgtbc1-af/fgtbc1-ar、fgtbc1-bf/fgtbc1-br分别从野生型禾谷镰刀菌菌株ph-1基因组dna中扩增目的片段a和b;利用引物hyg-f/hy-r,yg-f/hyg-r分别从质粒pcb1003中扩增目的片段h1和h2;利用soe-pcr方法,将片段a和h1连接起来、将片段h2和片段b连接起来,成为长片段ah和hb;将ah和hb同时加入至野生型禾谷镰刀菌菌株ph-1的原生质体中,运用同源重组的方法对目的基因fgtbc1进行敲除,即得到所述的禾谷镰刀菌fgtbc1基因敲除突变体δfgtbc1;以上所用引物序列如下:fgtbc1-af:5
’‑
gaagtagcggagcgggttc-3’,fgtbc1-ar:5
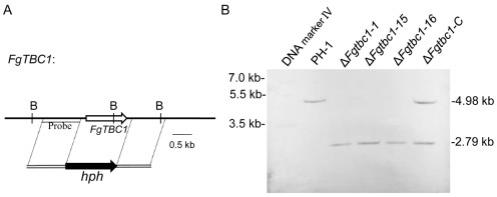
’‑
ttgacctccactagctccagccaagcc-cgccgaagtggctggttat-3’,fgtbc1-bf:5
’‑
gaatagagtagatgccgaccgcgggtt-ggcattctcctttgtcttc-3’,fgtbc1-br:5
’‑
tcttagctgggcacttgtat-3’,yg-f:5
’‑
gatgtaggagggcgtggatatgtcct-3’,hy-r:5
’‑
gtattgaccgattccttgcggtccgaa-3’,hyg-f:5
’‑
ggcttggctggagctagtggaggtcaa-3’,hyg-r:5
’‑
aacccgcggtcggcatctactctattc-3’。
9.上述一个特异调控禾谷镰刀菌子囊孢子成熟的基因fgtbc1在防控禾谷镰刀菌中的应用。
10.上述一个特异调控禾谷镰刀菌子囊孢子成熟的基因fgtbc1在研究禾谷镰刀菌致病机理中的应用。
11.上述一种禾谷镰刀菌fgtbc1基因敲除突变体δfgtbc1在研究禾谷镰刀菌致病机理中的应用。
12.相对于野生型禾谷镰刀菌ph-1菌株,上述的δfgtbc1突变体具有如下特征:(1)相对于野生型禾谷镰刀菌ph-1菌株,δfgtbc1突变体营养菌丝生长不受影响。
13.(2)相对于野生型禾谷镰刀菌ph-1菌株,δfgtbc1突变体分生孢子产生不受影响。
14.(3)相对于野生型禾谷镰刀菌ph-1菌株,δfgtbc1突变体子囊壳产生不受影响。
15.(4)相对于野生型禾谷镰刀菌ph-1菌株,δfgtbc1突变体子囊孢子成熟及喷射过程受影响。
16.本发明的优点在于:本发明经过了大量的实验,将fgtbc1基因从禾谷镰刀菌中成功敲除后,所得到的禾谷镰刀菌敲除突变体δfgtbc1的营养菌丝生长、分生孢子形成及产量均不受影响。有性生殖实验结果显示,该敲除突变体δfgtbc1在胡萝卜诱导培养基上能形成正常大小的子囊壳,子囊壳压片结果显示突变体δfgtbc1子囊孢子的成熟过程受到阻碍,无法产生正常的子囊孢子,进一步子囊壳喷射实验也显示突变体δfgtbc1无法喷射子囊孢子。上述结果说明fgtbc1基因特异调控禾谷镰刀菌有性生殖过程中子囊孢子的形成及发育。
附图说明
17.图1. 在禾谷镰刀菌中敲除fgtbc1基因的同源重组策略及其酶切位点图谱示意图(1a),southern杂交图谱(1b)。用bamhⅰ(b)酶切基因组dna,突变体δfgtbc1得到2.79 kb大小的条带,野生型ph-1得到4.98 kb大小的条带,回补菌株δfgtbc1-c同时出现上述两条带。
18.图2. 野生型ph-1、fgtbc1基因敲除突变体(δfgtbc1)及回补菌株δfgtbc1-c的菌落形态(2a)及菌落直径(2b)。
19.图3. fgtbc1基因敲除不影响分生孢子的产生。
20.图4. fgtbc1基因敲除不影响有性生殖过程子囊壳的形成。
21.图5. fgtbc1基因调控禾谷镰刀菌有性生殖过程中子囊孢子的成熟。
22.图6. fgtbc1基因调控禾谷镰刀菌有性生殖过程中子囊孢子的喷射。
23.图7. fgtbc1基因在禾谷镰刀菌有性发育阶段高表达。
具体实施方式
24.以下所述仅为本技术的优选实施案例,并不用于限制本技术,对于本领域的技术人员来说,本技术可以有各种变化和更改。凡在本技术的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本技术的保护范围之内。下述实施例中的实验方法,如无特殊说明,均为常规方法。下述实施例中所用的试验材料,如无特殊说明,均为常规生化试剂。
25.下述实施例中所使用的部分培养基的组成及配置方法:cm完全培养基:蔗糖10 g,酵母提取物6 g,酸性水解酪蛋白6 g,琼脂粉20 g,ddh2o定容至1 l,121℃,高压灭菌20 min备用。
26.cmc分生孢子产生培养基:酵母提取物1 g,nh4no
3 1 g/,kh2po
4 1 g,mgso47h2o 0.5 g,羧甲基纤维素钠15 g,ddh2o定容至1 l,121℃,高压灭菌20 min备用。
27.胡萝卜诱导培养基:去皮胡萝卜200 g,加适量水,榨汁机搅碎,琼脂粉20 g,ddh2o定容至1 l,121℃,高压灭菌20 min备用。
28.为进一步阐述一个特异调控禾谷镰刀菌子囊孢子成熟的基因fgtbc1及其应用,举例如下。
29.如本专利所描述,禾谷镰刀菌fgtbc1基因用斜体fgtbc1表示,蛋白用fgtbc1表示,fgtbc1基因敲除突变体用δfgtbc1表示,回补菌株用δfgtbc1-c表示,禾谷镰刀菌的野生型菌株用ph-1表示。
30.本发明经过长期大量的筛选,鉴定到一个特异调控禾谷镰刀菌有性生殖过程子囊孢子成熟的敲除突变体δfgtbc1,对应的基因为fgtbc1,其核苷酸序列为seq id no. 1,编码的蛋白质氨基酸序列为seq id no. 2。
31.实施例1:禾谷镰刀菌中fgtbc1基因的敲除利用引物fgtbc1-af/fgtbc1-ar,fgtbc1-bf/fgtbc1-br分别从野生型禾谷镰刀菌菌株ph-1基因组dna中扩增目的片段a和b;利用引物hyg-f/hy-r,yg-f/hyg-r分别从质粒pcb1003中扩增目的片段h1和h2。接着利用soe-pcr方法,将片段a和h1,h2和片段b连接起来,成为长片段ah和hb,将ah和hb同时加入至野生型禾谷镰刀菌菌株ph-1的原生质体中,运
用同源重组的方法对目的基因fgtbc1进行敲除(基本原理如图1a所示),从而获取fgtbc1敲除突变体δfgtbc1。southern杂交验证结果如图1b所示,通过bamh i酶切基因组dna,突变体δfgtbc1得到2.79 kb大小的条带,野生型ph-1得到4.98 kb大小的条带,回补菌株δfgtbc1-c同时出现上述两条带,符合预测结果。
32.所用到的引物序列为:fgtbc1-af:5
’‑
gaagtagcggagcgggttc-3’,fgtbc1-ar:5
’‑
ttgacctccactagctccagccaagcc-cgccgaagtggctggttat-3’,fgtbc1-bf:5
’‑
gaatagagtagatgccgaccgcgggtt-ggcattctcctttgtcttc-3’,fgtbc1-br:5
’‑
tcttagctgggcacttgtat-3’,yg-f:5
’‑
gatgtaggagggcgtggatatgtcct-3’,hy-r:5
’‑
gtattgaccgattccttgcggtccgaa-3’,hyg-f:5
’‑
ggcttggctggagctagtggaggtcaa-3’,hyg-r:5
’‑
aacccgcggtcggcatctactctattc-3’。
33.实施例2:禾谷镰刀菌中δfgtbc1突变体的营养生长及分生孢子测定分别将野生型ph-1、fgtbc1敲除突变体(δfgtbc1)及回补菌株(δfgtbc1-c)转接至cm固体培养基,28℃倒置培养3天,观察菌落形态并测量菌落直径。如图2a和图2b所示,ph-1、δfgtbc1及δfgtbc1-c的菌落形态及直径无明显变化,表明fgtbc1缺失不影响禾谷镰刀菌的营养生长。
34.为了分析fgtbc1对禾谷镰刀菌分生孢子产量的影响,我们将野生型ph-1、δfgtbc1及δfgtbc1-c接种至cmc液体产孢培养基中,28℃,180 rpm 振荡培养3天。取出孢子液,同等倍数稀释,血球计数板统计分生孢子数量。结果如图3所示,突变体δfgtbc1的产孢量与野生型ph-1及回补菌株δfgtbc1-c相比无显著性差异,表明fgtbc1基因的缺失不影响禾谷镰刀菌分生孢子的产量。
35.实施例3:禾谷镰刀菌中δfgtbc1的有性生殖过程分析将野生型ph-1,突变体δfgtbc1及回补菌株δfgtbc1-c接种至胡萝卜培养基中,放28℃培养箱培养5天左右,待菌丝长满后,于超净工作台中加入2.5%的吐温60,用无菌涂布棒压实菌丝。然后置于22℃,黑光灯光照培养箱,5天后观察到所有的胡萝卜培养基表面均长出黑色的子囊壳(图4)。随着培养时间的延长,野生型ph-1与fgtbc1敲除突变体生成子囊壳的数量逐渐增多,子囊壳表面出现黄色的物质cirrhi(图5a,箭头表示),而突变体δfgtbc1未观察到黄色的cirrhi(图5a)。选取培养至9天的子囊壳压片处理,进一步观察子囊孢子形态,结果如图5a所示,野生型ph-1形成典型的子囊孢子,δfgtbc1-1突变体释放出大量短棒状的畸形子囊,未观察到成熟子囊孢子(图5b)。为了更加客观地观察子囊及子囊孢子的释放情况,我们对野生型ph-1与fgtbc1敲除突变体进行子囊孢子喷射观察,结果发现fgtbc1敲除突变体子囊壳未能喷射出子囊孢子(图6)。以上结果表明fgtbc1在禾谷镰刀菌的有性生殖过程中发挥重要作用,fgtbc1敲除不影响子囊壳的产生,却特异调控子囊及子囊孢子的成熟与喷射。
36.实施例4:禾谷镰刀菌中fgtbc1在营养菌丝与子囊壳形成过程中的表达量为了探究fgtbc1在禾谷镰刀菌不同生长发育阶段的表达量变化,我们分别提取野生型ph-1菌丝(36 h)及子囊壳(5 day、9 day、13 day)的总rna,反转录成cdna。利用引物
fgtbc1-qf/fgtbc1-qr,fgactin-qf/fgactin-qr,通过实时荧光定量pcr检测fgtbc1在上述各个阶段的转录水平,以菌丝阶段为参照。
37.所用到的引物序列为:fgtbc1-qf:5
’‑
agatccgagaacaagcctactgc-3’;fgtbc1-qr:5
’‑
ttccgccttgaccaccttcatc-3’;fgactin-qf:5
’‑
acggaaacattgtcatgtctggtg-3’;fgactin-qr:5
’‑
ctctcgtcgtactcctgcttgg-3’。
38.结果如图7所示,fgtbc1在子囊壳生长发育的各个阶段均呈现不同程度的上调,表明fgtbc1在禾谷镰刀菌的有性生殖阶段高表达,发挥重要功能。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1