一种表达低聚糖脱支酶的基因工程菌及其应用
1.本发明涉及一种表达低聚糖脱支酶的基因工程菌及其应用,属于基因工程和酶工程技术领域。
背景技术:
2.淀粉是一种以α-d-葡萄糖为单体构成的天然高分子聚合物,分为直链淀粉和支链淀粉。直链淀粉是由葡萄糖单元通过α-1,4糖苷键构成的线性聚合物,支链淀粉以葡萄糖单元通过α-1,4糖苷键构成其主链,再在分支点处以α-1,6糖苷键连接不同数目葡萄糖单元构成其侧链,是一个具有复杂树枝形分支结构的大分子多糖。在淀粉加工过程中,一般需要多种淀粉酶对淀粉进行催化水解。水解α-1,4糖苷键的酶包括α-淀粉酶、糖化酶、β-淀粉酶,然而这些酶不能或只能缓慢作用于α-1,6糖苷键,限制了淀粉的水解速率。淀粉脱支酶能够专一、高效地水解支链淀粉中α-1,6糖苷键,利用淀粉脱支酶与其他糖化酶协同作用,从而加速后续酶的反应,缩短反应时间,达到提高原料利用率及生产效率的目的。淀粉脱支酶目前已经被用于葡萄糖浆、高麦芽糖浆、环糊精、分支环糊精、焙烤、啤酒酿造和抗性淀粉等淀粉加工领域。
3.根据其底物特异性的不同,淀粉脱支酶可分为普鲁兰酶、异淀粉酶、糊精脱支酶和低聚糖脱支酶。普鲁兰酶对低分子量的糊精水解活力高,能够高效地水解普鲁兰多糖,对高分子量糊精以及分支密集的糖原水解活力低,最小作用底物单元为麦芽糖基麦芽糖。异淀粉酶适宜作用于较高分子量糊精、支链淀粉以及糖原,对小分子量的分支糊精水解活力较低,且不能水解普鲁兰多糖,底物的最小单位为麦芽三糖基麦芽四糖。糊精脱支酶对中等分子量大小的麦芽糊精水解活力高,低聚糖脱支酶对更小链段糊精或低聚糖水解活力高。
4.在淀粉制糖工业中,淀粉经过调浆液化后加入糖化酶从而产生不同的淀粉糖,在糖化过程中协同添加脱支酶可以提高转化率,然而现阶段工艺还存在一定不足。如在制备葡萄糖浆时添加的葡萄糖淀粉酶会催化葡萄糖发生聚合反应,生成异麦芽糖、潘糖等低聚异麦芽糖,在制备麦芽糖浆时添加的β-淀粉酶不能作用于α-1,6糖苷键而产生大量β-极限糊精和其它寡糖。普鲁兰酶、异淀粉酶等脱支酶对此类小分子糊精及低聚糖的水解能力较弱,利用低聚糖脱支酶作用于此类较小的糊精及低聚糖,从而提高淀粉利用率,提高生产效率,并且减少副产物产生。
5.随着新型淀粉加工产品及工艺的不断创新发展,现代淀粉加工工业对淀粉脱支酶的性能提出了新的要求,包括不同的作用底物、不同的最适温度、高效性、稳定性等。因此,有必要开发新型的淀粉脱支酶,填补目前常用脱支酶底物特异性方面的空白,以适应更广泛的工业化生产的需求。
技术实现要素:
6.为了解决上述存在的技术问题,本发明通过将来源于paenibacillus sp.p22中编码低聚糖脱支酶的基因导入基因工程菌中实现低聚糖脱支酶的异源表达。
7.本发明提供了一种编码低聚糖脱支酶的基因,其基因核苷酸序列如seq id no.2所示。
8.本发明还提供了一种携带上述低聚糖脱支酶基因的重组载体。
9.本发明还提供了一种携带上述低聚糖脱支酶的基因,或上述重组载体的重组细胞。
10.本发明提供了一种重组大肠杆菌,所述重组大肠杆菌表达了来源于paenibacillus sp.p22的低聚糖脱支酶,所述低聚糖脱支酶的氨基酸序列如seq id no.1所示。
11.在本发明的一种实施方式中,编码所述低聚糖脱支酶的核苷酸序列如seq id no.2所示。
12.在本发明的一种实施方式中,所述重组大肠杆菌以e.coli bl21(de3)为表达宿主。
13.在本发明的一种实施方式中,所述重组大肠杆菌以pet-28a(+)为表达载体。
14.本发明还提供了上述重组大肠杆菌或氨基酸序列如seq id no.1所示的低聚糖脱支酶在水解麦芽糊精中的应用。
15.在本发明的一种实施方式中,编码所述低聚糖脱支酶的核苷酸序列如seq id no.2所示。
16.本发明还提供了一种水解麦芽糊精的方法,所述方法为:将上述重组大肠杆菌或其发酵制备得到的重组低聚糖脱支酶添加至含有麦芽糊精的反应体系中进行水解;或将氨基酸序列如seq id no.1所示的低聚糖脱支酶添加至含有麦芽糊精的反应体系中进行水解。
17.在本发明的一种实施方式中,所述重组低聚糖脱支酶是将重组大肠杆菌接种至种子培养基中,制备得到种子液,将种子液按照2%~4%(v/v)的接种量接种至发酵培养基中,在25~30℃下培养48~96h,制备得到重组低聚糖脱支酶粗酶液。
18.在本发明的一种实施方式中,所述反应体系的反应条件为:在45~55℃,ph 5.5~6.0的条件下反应24~36h。
19.在本发明的一种实施方式中,所述反应体系的反应条件为:在50℃,ph 6.0的条件下反应24h。
20.本发明还提供了上述重组大肠杆菌或氨基酸序列如seq id no.1所示的低聚糖脱支酶在制备水解麦芽糊精的产品中的应用。
21.在本发明的一种实施方式中,编码所述低聚糖脱支酶的核苷酸序列如seq id no.2所示。
22.在本发明的一种实施方式中,利用发酵所得的重组低聚糖脱支酶粗酶液或经镍柱纯化后得到的纯酶于50℃水解麦芽糊精进行脱支反应。
23.在本发明的一种实施方式中,所述底物的浓度至少为10g/l。
24.在本发明的一种实施方式中,所述低聚糖脱支酶在反应体系中的加酶量至少为50u/g。
25.本发明还提供了上述基因,或上述重组载体,或上述重组大肠杆菌,或上述方法在水解麦芽糊精中的应用。
26.有益效果
27.(1)本发明提供了一种具有特定碱基序列的低聚糖脱支酶,并且成功的采用大肠杆菌对低聚糖脱支酶进行异源表达,采用本发明的方法,所表达的低聚糖脱支酶粗酶液的催化活力可达258.83u/ml。
28.(2)本发明的低聚糖脱支酶最适温度为50℃,在50℃保温60min后活性仍能保持在50%以上,且在45℃保温时酶活力出现激活现象,能够满足淀粉加工生产中对不同反应温度的需求。
29.(3)本发明提供的低聚糖脱支酶是一种新来源的低聚糖脱支酶,该来源在此前尚未报道过,具有较高比酶活,底物特异性强。
30.(4)本发明的低聚糖脱支酶能够脱支麦芽糊精中dp《6的短链段,填补了目前常用脱支酶底物特异性方面的空白,在淀粉糖、抗性淀粉、啤酒生产及乙醇燃料等工业领域中具有较高的潜在应用前景。
附图说明
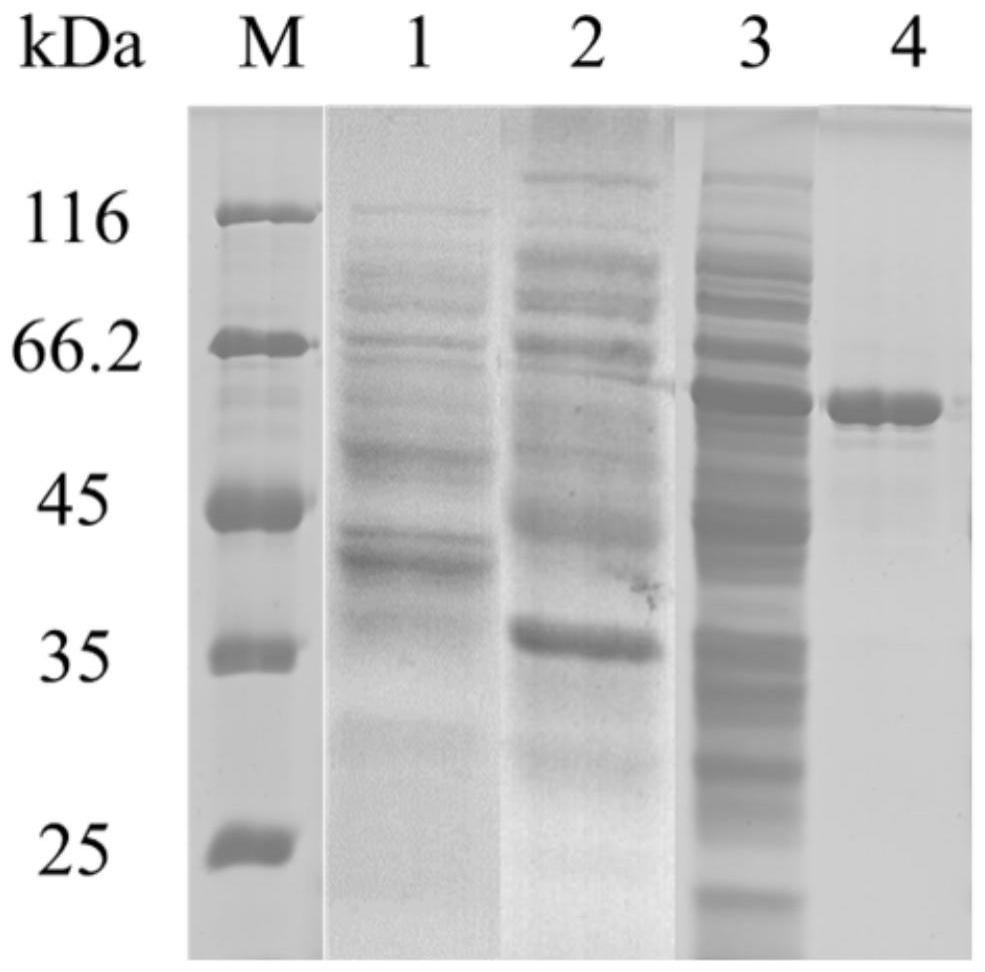
31.图1:重组低聚糖脱支酶的sds-page分析;其中,m:蛋白标准分子量;泳道1:e.coli bl21(de3)/pet-20b(+)-oga;泳道2:e.coli bl21(de3)/pet-22b(+)-oga;泳道3:e.coli bl21(de3)/pet-28a(+)-oga;泳道4:纯酶。
32.图2:不同温度下的低聚糖脱支酶的相对酶活数据。
33.图3:重组低聚糖脱支酶的在不同温度下的稳定性。
34.图4:重组低聚糖脱支酶水解de 2麦芽糊精的高效阴离子交换色谱(hpaec-pad)分析。
具体实施方式
35.下述实施例中所涉及的培养基如下:
36.lb液体培养基:1%(w/v)胰蛋白胨,0.5%(w/v)酵母提取物,1%(w/v)氯化钠,ph 7.0。
37.lb固体培养基:在lb液体培养基的基础上添加1.5(w/v)琼脂。
38.tb液体培养基:1.2%(w/v)胰蛋白胨,2.4%(w/v)酵母提取物,0.4%(w/v)甘油,17mm kh2po4,72mm k2hpo4,ph 6.0。
39.下述实施例中所涉及的检测方法如下:
40.重组低聚糖脱支酶酶活测定方法如下:吸取800μl底物对硝基苯α-d-葡萄糖苷(pnpg,10mm)于离心管中,加入150μl磷酸缓冲液(500mm,ph 6.0)混匀后50℃保温5min,加入50μl稀释后的待测酶液反应5min后加入1ml na2co3溶液(1m)终止反应,于410nm下测定吸光值。以高温灭活的酶液作为空白对照。
41.酶活定义:在上述条件下,每分钟水解生成1μmol的对硝基苯酚(pnp)为1个酶活力单位。
42.聚合度的检测方法:取反应后上清液稀释后过0.22μm水系滤膜,采用ics-5000高效阴离子交换色谱-脉冲安培检测器(hpaec-pad)测定低聚糖脱支酶处理后产物的聚合度分布情况。流动相由250mm(w/v)氢氧化钠和1m(w/v)乙酸钠组成,流速为0.5ml/min,柱温保
持在35℃。
43.实施例1:重组大肠杆菌分泌表达系统的构建
44.(1)以ncbi上公开的paenibacillus sp.p22基因组dna为模板,采用pcr法扩增出两端分别含nco i和xhol i限制性酶切位点的目的基因seq id no.2(低聚糖脱支酶)。
45.其中所涉及的pcr扩增程序为:taq buffer(mg
2+
plus)10μl,dntp mixture(各2.5mm)4μl,正向引物(10μm)1μl,反向引物(10μm)1μl,模板dna 1μl,taq dna polymerase(1.25u/μl)1μl,加入双蒸水至50μl。pcr扩增条件为:98℃预变性3min;再进行30个循环(98℃10s,60℃15s,68℃2min);最后68℃保温10min。
46.(2)将pcr扩增出的低聚糖脱支酶基因(seq id no.2)插入到pmd 18-t simple质粒中得到克隆载体pmd 18-t simple/oga,载体经双酶切后回收含有粘性末端的目的基因片段,并插入到经相同内切酶处理过的pet-28a(+)质粒中,得到重组表达载体pet-28a(+)-oga。
47.按照上述步骤(1)~(2),区别在于,将质粒pet-28a(+)分别调整为:pet-22b(+)、pet-20b(+),分别得到重组表达载体pet-22b(+)-oga、pet-20b(+)-oga。
48.(3)分别将步骤(2)制备得到的重组表达载体转化至大肠杆菌e.coli jm109中,涂布具有卡那霉素的lb平板,挑取转化子进行测序、菌落pcr验证,分别提取得到含有低聚糖脱支酶基因的重组质粒。
49.分别将重组质粒转化至e.coli bl21(de3)中,得到重组基因工程菌:e.coli bl21(de3)/pet-28a(+)-oga,e.coli bl21(de3)/pet-22b(+)-oga,e.coli bl21(de3)/pet-20b(+)-oga。
50.实施例2:重组低聚糖脱支酶的表达、分离、纯化
51.具体步骤如下:
52.1、重组低聚糖脱支酶的表达
53.(1)分别将实施例1制备得到的单菌落e.coli bl21(de3)/pet-28a(+)-oga,e.coli bl21(de3)/pet-22b(+)-oga,e.coli bl21(de3)/pet-20b(+)-oga接种于50ml含有20μg/ml卡那霉素的lb液体培养基中,37℃、200rpm条件下摇瓶培养8~12h,分别制备得到种子液;
54.(2)分别将上述制备得到的种子液按2%~4%(v/v)的接种量转接至50ml含有20μg/ml卡那霉素的tb液体培养基中,当od
600
=0.6时加入终浓度为0.01mm的iptg,25~30℃、200rpm下摇瓶培养48~96h,分别制备得到发酵液;
55.(3)分别将发酵液在4℃、10000rpm条件下离心20min,丢弃上清液,用50mm、ph6.0的醋酸钠缓冲液重悬菌体,在200w、开1s、关2s程序下,置于冰上超声破碎15min,最后在4℃、10000rpm条件下离心20min,取上清液,即为胞内粗酶液。
56.分别检测e.coli bl21(de3)/pet-28a(+)-oga,e.coli bl21(de3)/pet-22b(+)-oga,e.coli bl21(de3)/pet-20b(+)-oga制备得到的胞内粗酶液的酶活数据,并进行sds-page蛋白电泳验证,结果如表1和图1所示。
57.表1:不同重组菌株制备得到的重组低聚糖脱支酶的酶活数据
[0058][0059]
结果显示,采用pet-22b(+)、pet-20b(+)为表达载体时,没有检测到酶活,且蛋白电泳显示重组低聚糖脱支酶不能得到表达,只有采用pet-28a(+)为表达载体时,检测到所述重组低聚糖脱支酶得到了表达,经检测,其粗酶活为258.83u/ml。
[0060]
2、重组低聚糖脱支酶的纯化
[0061]
采用镍柱对步骤1中重组菌株e.coli bl21(de3)/pet-28a(+)-oga制备得到的重组低聚糖脱支酶粗酶液进行纯化,平衡液(a液,ph 7.5)为500mm nacl+50mm tris-hcl+20mm咪唑,洗脱液(b液,ph 7.5)为500mm nacl+50mm tris-hcl+500mm咪唑。发酵粗酶液经0.45μm水系膜过滤后,用5~6个柱体积的a液平衡镍柱;上样,流速为2ml/min;之后先用a液平衡离子柱,再采用60%的b液洗下目的蛋白,根据洗脱峰收集对应洗脱液,置于透析袋中,在4℃条件下采用50mm、ph 6.0的醋酸钠缓冲液进行过夜透析,随后通过sds-page蛋白电泳(结果如图1所示)及酶活测定的方式进行鉴定。经检测,纯酶的酶活为407.65u/mg。
[0062]
实施例3:重组低聚糖脱支酶的最适温度及热稳定性
[0063]
分别检测实施例2制备得到的经纯化后的重组低聚糖脱支酶最适温度及热稳定性,具体步骤如下:
[0064]
(1)重组低聚糖脱支酶的最适温度测定方法如下:按照酶活测定方法,分别测定酶液在30℃~80℃下的酶活力,以酶活最高者为100%,结果如表2和图2所示。
[0065]
表2:不同温度下的相对酶活数据
[0066][0067]
(2)重组低聚糖脱支酶的热稳定性测定方法如下:将酶液分别置于不同温度(45℃
~60℃)下保温,每隔一定时间将酶液取出,迅速冷却后,测定酶的残余酶活力,以未保温酶液的活力为100%,结果如表3和图3所示。
[0068]
表3:不同温度下保温不同时间的相对酶活数据
[0069][0070][0071]
结果显示,重组低聚糖脱支酶的最适温度和热稳定性如图2和图3所示。该酶在50℃时达到最佳活力,当温度进一步升高大于50℃时,酶的活力发生急剧下降,在70℃时酶已基本失活。该酶在45℃保温时酶活力稳定,并且随着时间延长酶活力有稍许增加。50℃孵育60min后,其活性仍保持在50%以上。
[0072]
实施例4:重组低聚糖脱支酶的应用
[0073]
本实施例中采用低聚糖脱支酶水解麦芽糊精;具体步骤如下:
[0074]
(1)配制浓度为1%(w/w)的de 2麦芽糊精溶液100g,调节ph至6.0。
[0075]
(2)将浓度为1%(w/w)的de 2麦芽糊精溶液置于50℃水浴摇床中预热10min,随后按照50u/g的加酶量添加实施例2制备得到的低聚糖脱支酶纯酶,在50℃水浴摇床中反应24h,随后转移至沸水中灭酶20min,以终止反应;其中,用等量的醋酸钠缓冲液替换酶液,作为空白对照。
[0076]
(3)待灭酶处理后,将反应液在10000rpm条件下离心10min,取上清液过0.22μm的水系膜去除杂质后,即可用离子色谱分析反应液中不同聚合度的分布情况。为探究低聚糖脱支酶在水解麦芽糊精中的底物特异性,测定反应前后体系中不同dp值寡糖链的峰面积,用峰面积表示其含量,从而表征低聚糖脱支酶水解底物α-1,6糖苷键的情况。色谱分析结果如表4和图4所示。
[0077]
表4:不同聚合度的分布情况
[0078][0079]
结果显示,以de 2麦芽糊精为底物,经过低聚糖脱支酶水解后,体系中dp《6的链段含量均上升,说明低聚糖脱支酶作用于麦芽糊精的α-1,6糖苷键,使得短支链的分支点被水解,生成了一系列dp《6的寡糖链。因此,低聚糖脱支酶在水解麦芽糊精中倾向于水解dp《6的分支点。
[0080]
实施例5:重组低聚糖脱支酶的应用
[0081]
具体实施方式同实施例4,区别在于,将步骤(1)中底物分别替换成浓度为1%(w/w)的de 4麦芽糊精溶液100g、浓度为1%(w/w)的de 6麦芽糊精溶液100g。
[0082]
按照实施例4的方法,测定反应前后体系中不同聚合度的含量变化。结果如表5所示。
[0083]
表5:不同聚合度的分布情况
[0084][0085]
结果显示:低聚糖脱支酶在水解不同de值的麦芽糊精时,都能够特异性水解麦芽糊精中的短分支,生成一系列dp《6的寡糖链。
[0086]
此外,随着麦芽糊精de值的增加,低聚糖脱支酶水解生成dp《6的寡糖链含量随之上升,表明低聚糖脱支酶对de值更高的麦芽糊精亲和力更强。
[0087]
综上,低聚糖脱支酶对麦芽糊精具有一定水解作用,能够水解底物中的α-1,6糖苷键,且底物特异性高,特异性水解dp《6的分支点。其对短支链的特异性填补了目前常用脱支酶底物特异性方面的空白,在淀粉糖、抗性淀粉、啤酒生产及乙醇燃料等工业领域中具有较高的应用潜力。
[0088]
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1