油菜BnNRT2.3-like基因及其表达载体在促进植物氮利用中的应用
油菜bnnrt2.3-like基因及其表达载体在促进植物氮利用中的应用
技术领域
1.本发明属于植物基因工程技术领域,具体涉及甘蓝型油菜中的硝酸盐转运体基因bnnrt2.3-like的分离克隆、表达载体构建以及其在提高植物氮利用效率中的应用。
背景技术:
2.油菜对氮素的需求量较大,如果缺氮,油菜生长就会受到影响,出现植株矮小、老叶黄化、甚至减产。目前油菜的高产过于依赖氮肥的大量施用,但过量施用氮肥,不仅会造成资源浪费,还会降低油菜氮素利用率,降低油菜抗病虫害的能力,故提高油菜的氮素利用效率具有重要的现实意义。
3.高等植物的主要氮源是硝酸盐(no
3-)和铵盐(nh
4+
),通常情况下在水淹土壤中生长的植物更喜欢nh
4+
,陆生植物都比较偏好no
3-,故一般认为油菜是喜硝作物。硝酸盐在根中被吸收后可以通过木质部运输到地上部分的组织和器官,然后在植物细胞内经硝酸盐还原酶(nr)、亚硝酸盐还原酶(nir)、谷氨酰胺合成酶(gs)等多种酶共同参与下被同化成氨基酸及其他含氮化合物。硝酸盐不仅作为营养物质调控植物生长和发育,也作为信号物质调控基因的转录从而影响种子萌发、植物根系生长、气孔活动等。研究表明,增硝可以提高植物根系细胞分裂素的浓度,促进植株的生长发育,同时影响其他营养物质在体内的运输,促进植物根系的生长。
4.植物根系从土壤中吸收硝态氮、植物体内硝态氮的运输和细胞内硝态氮再分配等都需要硝酸盐转运蛋白去执行,编码硝酸盐转运蛋白的主要有nrt1家族负责低亲和(lats)硝酸盐转运系统和nrt2家族负责高亲和(hats)硝酸盐转运系统,nrt2在植物中以基因家族形式存在。植物nrt2家族成员数目普遍较少,如水稻中有5个,拟南芥中有7个,油菜中有17个。其中研究较多的是水稻和拟南芥中的nrt2基因。osnrt2.1和osnrt2.2基因位于水稻2号染色体上,聚为一个亚类;osnrt2.3和osnrt2.4基因位于水稻1号染色体上,也聚为一个亚类。osnrt2.1和osnrt2.2基因编码相同的氨基酸,都受硝酸盐诱导。osnrt2.1的表达量随硝酸盐吸收量的上升而上升,表明osnrt2.1是水稻吸收硝酸盐的一个关键基因。用铵盐处理水稻植株,发现osnrt2.2表达未受到抑制,表明osnrt2.2在淹水土壤中仍能转录表达,是能提高水稻氮素利用效率的重要基因。osnrt2.3存在osnrt2.3a和osnrt2.3b两个转录本,唐仲等研究发现,osnrt2.3a定位在细胞质膜上,主要在根部木质部薄壁细胞中表达,在低氮条件下,osnrt2.3a会影响木质部硝酸盐的装载进而影响植株的生长,推测osnrt2.3a在低氮水平下能促进硝态氮从根到茎长距离的转运;范晓荣等系统研究了osnrt2.3b转运蛋白,发现osnrt2.3b基因也定位在细胞质膜上,主要在韧皮部中表达,能够通过感应外界ph值变化来调控硝酸盐转运活性,过表达osnrt2.3b基因能促进水稻对n的吸收,显著提高水稻的产量和氮利用效率。wei等发现osnrt2.4是一个双亲和性硝酸盐转运蛋白,能够调节水稻根部发育,影响水稻中硝酸盐的分布。拟南芥中,filleur等从tdna突变体中分离到了anrt2.1基因。atnrt2.1和atnrt2.2基因均在拟南芥1号染色体上,负责拟南芥中硝酸盐的吸收;只
有植物氮饥饿时,atnrt2.4和atnrt2.5基因才会参与硝酸盐的吸收;atnrt2.5是参与高亲和硝酸盐转运的主要蛋白之一,当拟南芥长时间缺氮时atnrt2.5被高度诱导。atnrt2.1、atnrt2.4和atnrt2.5基因之间的空间表达模式不同,atnrt2.1主要在主根较成熟的部位中表达,atnrt2.4主要在主根较幼嫩的部分和侧根末端表达,atnrt2.5主要在主根和侧根的根须区域表达。此外atnrt2.4和atnrt2.5还在茎中韧皮部表达,影响茎中韧皮部硝酸盐的含量。atnrt2.7主要在籽粒中表达,且是nrt2基因家族中唯一定位在液泡膜上的基因,主要负责液泡中确酸盐的转运,调节种子硝酸盐含量,控制种子休眠。在甘蓝型油菜中,tang等详细鉴定了17个nrt2基因家族成员,其中大多数nrt2基因成员被n饥饿上调,与拟南芥nrt2的表达模式很相似。bnnrt2.1s、bnnrt2.2a和bnnrt2.4a均特异性表达于根组织,而bnnrt2.7a和bnnrt2.7b主要表达于地上部,bnnrt2.5s在茎和根组织中均有表达。由此可以发现,拟南芥和水稻中已克隆的nrt2基因的功能研究较为全面,而油菜中的nrt2基因虽然有所鉴定,但其中大部分基因的功能尚未进行系统的研究。
技术实现要素:
5.有鉴于此,本发明从甘蓝型油菜中分离克隆出了一种能够促进植物尤其是油菜吸收利用no
3-的高亲和硝酸盐转运蛋白基因bnnrt2.3-like,其在植物应答特别是油菜对低氮胁迫的信号转导途径中发挥重要的作用,过量表达该基因可提高油菜的耐低氮能力,故其在植物氮素营养高效分子育种中具有较大的经济价值。
6.本发明的技术方案具体如下:
7.发明人通过对已报道的拟南芥高亲和硝酸盐转运蛋白提高植物氮效率功能进行分析,发现atnrt2.3在植物氮素利用中有重要作用,但在油菜中的同源性基因功能尚无报道。
8.本发明通过大量序列比对分析,从甘蓝型油菜westar品种中发现了一个能促进植物氮吸收利用的油菜bnnrt2.3-like基因,其核苷酸序列如seq id no.1所示,编码的蛋白质的氨基酸序列如seq id no.2所示。
9.本发明基于bnnrt2.3-like基因序列,进一步构建了重组表达载体。
10.在某一具体的实施例中,上述重组表达载体中的包含启动子double camv 35s。该表达载体的构建方法为:利用序列如seq id no.5~6所示的引物对从pcambia1300s载体中扩增得到double camv 35s片段,然后连接到pcambia2301载体的ecori/hidiii酶切位点得到重组质粒;采用序列如seq id no.7~8所示的引物对扩增bnnrt2.3-like基因,所得产物与所述重组质粒各自酶切、回收后连接即得bnnrt2.3-like过量表达载体,记为pcambia2301-1300-d35s-bnnrt2.3-like-nos。
11.本发明还提供了一种提高甘蓝型油菜氮利用的方法,具体为:将bnnrt2.3-like基因的序列连接入表达载体中,得到bnnrt2.3-like过量表达载体;将bnnrt2.3-like过量表达载体转入宿主菌并转化甘蓝型油菜,筛选得到转基因植株并通过qpcr鉴定其表达水平。
12.优选地,宿主菌为农杆菌gv3101。
13.优选地,上述技术方案中转化甘蓝型油菜的方法为:将含有bnnrt2.3-like过量表达载体的农杆菌gv3101的菌液侵染甘蓝型油菜的下胚轴。
14.优选地,所述qpcr鉴定时所用引物序列如seq id no.13~14所示。
15.通过对阳性转基因苗中bnnrt2.3-like基因的转录水平进行验证,发现bnnrt2.3-like基因的表达量显著提高。进一步实验证明,bnnrt2.3-like基因过表达株系比野生型植株显著耐低氮胁迫,其主根长、地上部干重、谷氨酰胺合成酶活性、叶绿素和含量和叶片硝酸盐含量都显著高于野生型植株;而且在正常水肥管理的盆栽和大田中,bnnrt2.3-like过表达植株的叶面积显著高于野生型植株。故重组表达载体能够用于促进油菜氮利用。
16.另外,bnnrt2.3-like基因的过表达增强了油菜对核盘菌的抗性,故该基因及其重组表达载体有望用于提高油菜抗菌核病或用于抗菌核病育种。
17.本发明的有益效果为:
18.(1)本发明克隆的bnnrt2.3-like基因未被报道,丰富了油菜中对这类硝酸盐转运蛋白nrt2基因的需求,也为其它作物氮高效育种提供了新的遗传资源,对提高其它作物对氮素的利用效率具有指导和借鉴作用。
19.(2)本发明构建的过表达载体pcambia2301-1300-d35s-bnnrt2.3-like-nos转化油菜后获得的bnnrt2.3-like过量表达株系,从bnnrt2.3-like的功能获得方面进行研究,为nrt2.3-like的硝酸盐转运功能研究提供了原材料,具有重要意义。
20.(3)本发明通过对bnnrt2.3-like过量表达株系在氮高效方面的一系列研究,充分证明了bnnrt2.3-like参与到了植物对氮素的转运和利用中,并扮演着重要的角色。通过调控bnnrt2.3-like基因的表达能够调节植物体内氮素的积累,提氮同化相关酶的活性和油菜对菌核病的抗性。
21.(4)bnnrt2.3-like的开发和利用将有助于解决生产上油菜低氮素利用率和低菌核病抗性问题,有助于降低氮肥过量使用带来的系列问题。bnnrt2.3-like基因的克隆有助于培育有一定抗病性的高氮素利用效率油菜品种,也为bnnrt2.3-like在主要油料作物(油菜)氮高效方面的应用提供了理论与实践依据。
附图说明
22.图1为测序后的油菜bnnrt2.3-like基因与已知的拟南芥atnrt2.3-like基因编码的氨基酸序列比对结果图;
23.图2为实施例2中植物转化载体pcambia2301-1300-d35s-bnnrt2.3-like-nos的构建过程示意图;
24.图3为实施例3中农杆菌介导的甘蓝型油菜下胚轴遗传转化体系示意图;
25.图4为实施例3中油菜bnnrt2.3-like过表达转基因株系阳性苗的鉴定和表达量分析图;
26.图5为实施例3中田间wt与bnnrt2.3-like过表达植株收获后表型及氮素积累对比图;
27.图6为实施例3中bnnrt2.3-like过表达转基因株系对菌核病的抗性鉴定;
28.图7为实施例3中野生型和bnnrt2.3-like(oe)的植株苗期盆栽中正常管理的植株叶片对比图;
29.图8为实施例4中野生型和bnnrt2.3-like(oe)的植株在水培低氮胁迫处理7d后油菜植株表型对比图;
30.图9为实施例4中低氮胁迫处理7d后的野生型和bnnrt2.3-like(oe)的植株生物
by quality analysis tools.nucleic acids research,1997,25:4876-82)程序进行氨基酸序列的比对发现,这个目的条带与拟南芥的nrt2.3在氨基酸水平上的同源性为93%,经与ncbi数据库比对的结果一致(图1,图中bn即甘蓝型油菜(brassica napus l.),at为拟南芥(arabidopsis thaliana))。
40.实施例2植物过表达载体的构建
41.根据ncbi公布的camv 35s序列(gene id:aj007626)信息,使用primer premier 5.0软件设计如下的引物:
[0042]5’‑
gaattcttaattaagagctcgcatgcc-3’(seq id no:5);
[0043]
和5
’‑
ggtaccgtccccgtgttctctccaa-3’(seq id no:6)。
[0044]
利用上述引物通过pcr方法从pcambia1300s载体(购自北京鼎国昌盛生物技术有限责任公司)中扩增得到double camv 35s片段,然后将其连接到pcambia2301载体(来自华中农业大学王晶老师)的ecori/hidiii酶切位点,最终得到终载体pcambia2301-1300s(11634bp)。
[0045]
在bnnrt2.3-like ta克隆扩增引物两端分别加上salⅰ/bamhⅰ酶切位点及对应的保护碱基,其中所述引物具体如下所示:
[0046]
oe-bnnrt2.3-like-f:5
’‑
tctcgagctttcgcgagctcatggcttctaatgaagaa-3’(seq id no:7);
[0047]
oe-bnnrt2.3-like-r:5
’‑
aggtcgactctagaggatccttacggaaacgtgaaaca-3’(seq id no:8)。
[0048]
以bnnrt2.3-like的ta阳性克隆菌落质粒为模板进行pcr扩增,得到的pcr产物的长度为1744bp,进行salⅰ/bamhⅰ双酶切,同时对pcambia2301-1300s载体也进行双酶切。酶切反应体系为:基因片段或载体10μg,salⅰ2.5μl,bamhⅰ2.5μl,10*m buffer 15μl,ddh20加至总体积150μl,混匀后将其置于37℃静置8~12h。分别回收双酶切基因片段和pcambia2301-1300s质粒片段,用t4连接酶将bnnrt2.3-like双酶切基因片段连接并构建到pcambia2301-1300s载体的salⅰ/bamhⅰ酶切位点上,从而获得甘蓝型油菜bnnrt2.3-like转基因过量表达载体pcambia2301-1300s-d35s-bnnrt2.3-like-nos,在其t-dna区内含有抗卡那霉素的基因序列,bnnrt2.3-like过量表达的启动子为double camv35s。
[0049]
上述过表达载体的构建过程以及各载体结构具体如图2所示,图中lb为t-dna左边界,35s为花椰菜病毒35s启动子,mcs为多克隆位点,nos为终止子,rb为t-dna右边界。
[0050]
实施例3过表达bnnrt2.3-like转基因油菜植株的获得
[0051]
本实施例通过常规的下胚轴侵染法将重组质粒(即过量表达载体)pcambia2301-1300s-d35s-bnnrt2.3-like-nos转入农杆菌gv3101,筛选并分化成苗,具体步骤如下:
[0052]
(1)播种:用75%的酒精清洗甘蓝型油菜westar种子1min,再用0.1%的升汞溶液清洗种子5min,最后用无菌水清洗种子5次。用灭过菌的镊子把种子放入m0固体培养基中,将接种的种子在24℃下暗培养5天。其中,m0固体培养基具体为:1962年配方的ms无机盐和微量元素4.404g/l,调ph至5.8-5.9,加琼脂7g/l,按照常规高压蒸汽灭菌法进行灭菌。
[0053]
(2)农杆菌的活化:播种3天后,取-80℃保存的农杆菌菌种gv3101在lb固体培养基上划线,于28℃培养6h;挑单菌落于5ml lb液体培养基中,28℃,200rpm振荡培养20-24h,使农杆菌生长至对数期;取500μl培养好的菌液于50ml的lb液体培养基扩大培养约12h。其中,
所述的lb固体培养基含有抗生素利福平(50mg/ml)、庆大霉素(50mg/ml)和卡那霉素(50mg/ml)。
[0054]
(3)侵染菌液制备:把活化好的农杆菌液倒入2个50ml无菌离心管中,3500rpm离心15min,去上清,置于冰上,用1ml的dm液体培养基(清洗菌液,离心去上清,加入2-3ml的dm液体培养基重悬菌体,使侵染菌液od
600
值约为0.6。其中,dm液体培养基的制备方法为:ms无机盐(murashige and skoog,1962年)和微量元素(murashige and skoog,1962年)4.404g/l,蔗糖30g/l,调培养基的ph至5.8~5.9,在121℃灭菌20min;灭菌完成在无菌操作台内加入2,4-d 1mg/l,as 100mmol/l,kt 0.3mg/l。
[0055]
(4)侵染外植体:将步骤(3)中的侵染菌液置于冰上,同时用解剖刀切好步骤(1)中暗培养甘蓝型油菜幼苗的下胚轴(每段下胚轴的长度为0.8-1.0cm为宜),用无菌镊子把切好的下胚轴转入装有侵染菌液的平皿,视菌液浓度侵染15-30min(每隔3min摇晃一下)。
[0056]
(5)共培养:将侵染完成的外植体转至装有滤纸的平皿(应提前灭菌处理)中,吸干外植体表面可见的侵染液,然后转入m1固体培养基中,24℃暗培养40~48h。其中,m1固体培养基具体为:murashige and skoog,1962年配方的ms无机盐以及微量元素4.404g/l,甘露醇18g/l,蔗糖30g/l,2,4-d 2mg/l,kt 0.3mg/l,调培养基的ph至5.8~5.9,加琼脂7g/l,灭菌后加入100mmol/l as。
[0057]
(6)筛选:将共培养完成的外植体转至m2固体培养基上,进行20天的筛选培养,条件为:24℃,16h光照/8h暗培养。其中,m2固体培养基具体为:murashige and skoog,1962年配方的ms无机盐以及微量元素4.404g/l,甘露醇18g/l,蔗糖30g/l,2,4-d 2mg/l,kt0.3mg/l,调培养基的ph至5.8~5.9,加琼脂7g/l。
[0058]
(7)分化培养:将筛选后的外植体转至m3固体培养基上,开始分化培养,每隔15-20天继代一次,直到分化出芽(培养条件与步骤(6)中筛选条件一致)。其中,m3固体培养基具体为:murashige and skoog,1962年配方的ms无机盐以及微量元素4.404g/l,葡萄糖10g/l,木糖0.25g/l,酵母浸出(液)膏(mes)0.6g/l,调培养基的ph至5.8-5.9,加琼脂7g/l,灭菌后加入玉米素(zt)2mg/l,iaa(吲哚乙酸)0.1mg/l,tmt试剂250mg/l,kan 25mg/l。
[0059]
(8)生根培养:等到分化出芽的幼苗上能发现明显生长点后,用解剖刀贴着愈伤组织和芽相接的位置,小心翼翼地把幼苗切下(操作过程中一定要小心不能伤害到生长点),然后把幼苗转移至m4固体培养基中生根。其中,m4固体培养基具体为:murashige and skoog,1962年配方的ms无机盐和微量元素2.202g,sucrose 10g/l,iba(吲哚丁酸)0.5mg/l,调培养基的ph至5.8~5.9,加琼脂7g/l,灭菌后加入tmt 250mg/l,kan 25mg/l。
[0060]
上述农杆菌介导的甘蓝型油菜下胚轴遗传转化过程如图3所示,a图为经75%的酒精和0.1%升汞消毒后的甘蓝型油菜westar种子播于培养基上,b图为经含有转化载体的农杆菌gv3101中侵染后的油菜下胚轴置于m1培养基中共培养,c图为共培养后的下胚轴转入m2培养基中经16h光照/8h暗培养周期选择培养15~20d,d图为选择培养后的外植体置于m3培养基中经16h光照/8h暗培养周期选分化培养15d以上直至出芽,e图为分化培养出芽后的外植体置于新鲜m4培养基中经16h光照/8h暗培养周期选择分化培养15d以上直至分化成幼苗。
[0061]
进一步通过以下过程对转基因植株进行鉴定:
[0062]
①
.采用常规ctab法对甘蓝型油菜叶片基因组dna进行提取,具体步骤为:取长度
为1~2cm的幼嫩甘蓝型油菜叶片,放置于提前预冷的研钵中,途中加2~3次液氮研磨至细微的浆状后,转移到1.5ml离心管中,加入700μl 2x ctab溶液。70℃温育30min,间隔6min轻柔摇晃一次,70℃温育30min间隔10min轻柔摇晃一次。冷却至室温,再加入700μl体积比为25:24:1的tris饱和酚:氯仿:异戊醇,反复颠倒混匀后轻轻摇动40次左右。在3100rpm,室温离心15min。吸取上清约500μl,加入等体积的24:1的氯仿:异戊醇。摇匀后在100rpm,室温离心15min。弃上清加入冰冻的-20℃无水乙醇1ml在-20℃冰浴30min后,12000rpm,室温离心10min。用75%的酒精泡洗并反复吹打沉淀3min去盐。倒掉酒精,风干后每个样品加30~50μl ddh2o溶解。抽提的甘蓝型油菜基因组dna用nanodrop微量核酸检测仪检测浓度。
[0063]
②
.阳性转基因植株检测:以提取的dna为模板,用pcr扩增载体上的kan基因进行阳性苗鉴定,其中所用的引物具体为:
[0064]
kan-f:5
’‑
actgggcacaacagacaatcg-3’(seq id no:9);
[0065]
kan-r:5
’‑
gcatcagccatgatggatacttt-3’(seq id no:10)。
[0066]
检测结果如图4中a图所示:oe-1、oe-18、oe-25、oe-36和oe-50这5个转化植株(oe即overexpression的缩写)均能够扩增出预期大小的电泳条带(289bp),而非转基因的野生型对照则没有相应的电泳条带,表明转基因油菜基因组中已经含有外源基因dna片段。
[0067]
③
.过表达bnnrt2.3-like转基因油菜的qpcr鉴定
[0068]
a.甘蓝型油菜叶片基因组rna的提取:利用super总rna提取试剂盒(购自promega公司,美国),抽提wt和
②
中鉴定出的bnnrt2.3-like过表达阳性株系叶片的总rna。并利用ii q rt supermix for qrna(+gdna wiper)试剂盒(购自vazyme公司,中国)将rna反转录成cdna。
[0069]
b.为了确定bnnrt2.3-like在油菜中是否过量表达,采用时荧光定量pcr方法对鉴定得到的转基因植株进行分析。qpcr使用green realtime pcr master mix-plus-试剂盒(宝生物工程(大连)有限公司),采用油菜管家基因bnactin7(brassica napus actin-7)作为内标基因,其引物由南京金斯瑞生物科技有限公司合成。
[0070]
bnactin7(gene id:106418315)的qpcr引物为:
[0071]
bnactin7-f1:5
’‑
tcttcctcacgctatcctccg-3’(seq id no:11),
[0072]
bnactin7-r1:5
’‑
agccgtctccagctcttgc-3’(seq id no:12);
[0073]
bnnrt2.3like基因的qpcr引物为(该对引物是以实施例1中测序得到的核苷酸序列设计的qpcr特异性引物):
[0074]
q-bnnrt2.3-like-f:5
’‑
atgtttttgagaccgtctagcg-3’(seq id no:13),
[0075]
q-bnnrt2.3-like-r:5
’‑
ggttcgcatcagagttccag-3’(seq id no:14)。
[0076]
pcr程序:95℃预变性30sec,然后经40个循环(95℃10sec,60℃10sec,72℃26sec)。
[0077]
检测结果如图4中b图所示:oe-18、oe-25、oe-36和oe-50株系的bnnrt2.3-like表达水平均显著高于非转基因的野生型(wt)对照。表明这几个株系为独立的bnnrt2.3-like过表达转基因株系。
[0078]
④
.转基因植株的田间培养
[0079]
将鉴定得到的转基因植株和野生型在相同的田间水肥管理下进行大田培养,待植
株收获时,对bnnrt2.3-like过表达油菜的田间产量和农艺性状进行记录,并分析bnnrt2.3-like过表达对植株表型和氮素积累的影响。
[0080]
bnnrt2.3-like过表达油菜与野生型收获时的株高、全株有效角果数、角果长度、每角果粒数千粒重和单株籽粒产量的比较如下表所示:
[0081][0082]
从上表可知,与野生型相比,各bnnrt2.3-like过表达植株在株高、全株有效角果数、角果长度、每角果粒数千粒重和单株籽粒产量上均显著增加。
[0083]
图5为植株收获后表型及氮素积累对比图,其中a图为wt植株(左)与bnnrt2.3-like过表达植株(右)在成熟期收获后的表型对比,b图为wt与bnnrt2.3-like过表达植株收获后的总生物量,c图为wt与bnnrt2.3-like过表达植株收获后地上部份氮素积累量。表明在相同的田间水肥管理下,bnnrt2.3-like过表达植株的成熟期长势明显优于野生型且总生物量显著增加,但地上部氮素积累量增加但不显著。
[0084]
⑤
.bnnrt2.3-like过表达转基因油菜对菌核病的抗性鉴定
[0085]
取wt植株与bnnrt2.3-like过表达植株的苗期叶片离体接种油菜菌核病菌(sclerotinia sclerotiorum),接种36小时后观察发病表型(图6中a图)和病斑大小(图6中b图)。
[0086]
图中wt+s.sclerotiorum和bnnrt2.3-like(oe)+s.sclerotiorum分别表示用野生型和bnnrt2.3-like过表达株系接种核盘菌。从图6可知:接菌36小时后bnnrt2.3-like(oe)株系叶片的菌丝扩展面积明显小于对照,说明bnnrt2.3-like过表达后,植株对菌核病的抗性增强。
[0087]
另外,发现在正常水肥管理的盆栽和大田中,bnnrt2.3-like(oe)植株叶面积显著高于野生型植株,具体如图7所示。
[0088]
实施例4 bnnrt2.3-like过表达转基因油菜对低氮环境的耐受能力
[0089]
本实施例通过以下过程检测bnnrt2.3-like过表达株系在低氮环境中的生理反应:
[0090]
将供试材料的种子在阳光下晾晒2h,再用纯水浸泡1d,然后播种于纯水浸润的纱布上,每天加水以保证纱布处于润湿状态。播种约7-9d后,选择根长符合要求的幼苗移入营养液中,设置低氮胁迫(-n,0.3mm n)和正常供氮(+n,6mm n)两种处理进行营养液培养,每种处理5个生物学重复。以no
3-(ca(no
3-)2)为唯一氮源,大量元素和微量元素分别采用阿夫道宁和阿农营养液配方(常规),铁盐为0.05mm edta-fe;具体培养方法参考杨凝眉(杨凝眉,甘蓝型油菜响应缺氮胁迫的基因表达谱及共表达网络分析[d],2019)。
[0091]
图8为野生型(wt)和bnnrt2.3-like(oe)的植株在水培低氮胁迫处理7d后油菜植
株表型,发现在7d时野生型油菜下部两片叶已经开始黄化,子叶已黄化或者脱落,而bnnrt2.3-like过表达植株则仅一片有黄化趋势,表明在低氮胁迫下,bnnrt2.3-like过表达能促进主根伸长,延缓叶片黄化。
[0092]
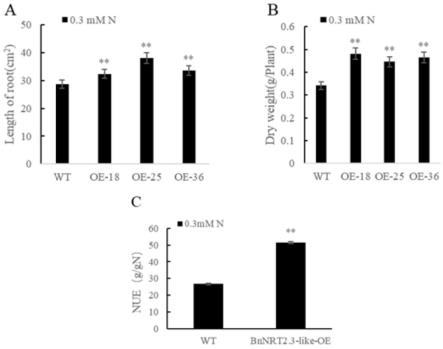
图9为低氮胁迫处理7d后的野生型(wt)和bnnrt2.3-like(oe)的植株生物量、主根长和氮素利用效率。其中,a图为低氮胁迫处理7d后wt与bnnrt2.3-like(oe)的主根长,b图为低氮胁迫处理7d后wt与bnnrt2.3-like(oe)植株的生物量,c图为低氮胁迫处理7d后wt与bnnrt2.3-like(oe)植株的氮素利用效率。从图9可知,低氮胁迫处理下bnnrt2.3-like(oe)植株的生物量和氮素利用效率显著高于wt,bnnrt2.3-like(oe)植株的主根显著长于wt,说明bnnrt2.3-like在低氮环境中能促进植物根的生长并提高植物氮素利用效率,从而促进植株硝态氮的吸收利用。
[0093]
在低氮处理7天后,取其新鲜的部位一致的功能叶片提取硝酸还原酶和谷氨酰胺合成酶并测定酶活性,同时测定叶片的硝酸盐含量和叶绿素浓度进行分析。图10为低氮胁迫处理7d后野生型(wt)和bnnrt2.3-like(oe)的植株叶片的亚硝酸还原酶(nr)活性、谷氨酰胺合成酶(gs)活性、硝态氮含量和叶绿素含量。其中,a图为低氮胁迫处理7d后wt与bnnrt2.3-like(oe)植株叶片的nr活性,b图为低氮胁迫处理7d后wt与bnnrt2.3-like(oe)植株叶片的gs活性,c图为低氮胁迫处理7d后wt与bnnrt2.3-like(oe)植株叶片的叶绿素含量,d图为低氮胁迫处理7d后wt与bnnrt2.3-like(oe)植株叶片的硝态氮含量。
[0094]
从图10可知,低氮处理7d后的bnnrt2.3-like(oe)植株叶片在硝态氮含量、nr活性、gs活性和叶绿素含量上均显著高于wt,说明在低氮环境下bnnrt2.3-like过表达能促进植物对硝态氮的同化,对低氮胁迫中的植物氮同化、提高氮素利用效率有重要作用。
[0095]
综上所述,bnnrt2.3-like过表达植株相对于野生型植株,不仅长势更优、总生物量显著增加,而且bnnrt2.3-like基因的过表达增强了油菜对核盘菌的抗性。在低氮胁迫下,bnnrt2.3-like过表达植株的主根长、地上部干重、谷氨酰胺合成酶活性、叶绿素和含量和叶片硝酸盐含量都显著高于野生型植株。由此可见,过表达bnnrt2.3-like基因不仅可以在植物吸收利用硝态氮的过程中发挥重要作用,还为提高油菜菌核病抗性提供了新的思路,这对于培育油菜氮高效品系具有重要的意义。
[0096]
以上所述是本发明的优选实施方式,不能以此来限定本发明之权利范围,应当指出,对于本技术领域的普通技术人员来说,凡在本发明的精神和原则之内所作的任何修改、等同替换和改进等,均应包含在本发明的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1