一种紫心甘薯花色素苷合成调控因子IbPPA及其应用
本发明涉及植物遗传与变异的分子机制领域,具体涉及一种紫心甘薯花色素苷合成调控因子ibppa及其应用。
背景技术:
1、目前,对于植物的花、果、茎、叶等地上部分花色素苷合成以及环境信号因子影响的分子机制有了一定的认识,尤其是光照对花色素苷合成的调控机制及信号传导过程已比较清楚。对于非依光型或无法直接接受光照的植物根、块根或块茎中花色素苷合成调控机制及信号传递过程却不甚明了。
2、调控花色素苷生物合成的转录因子主要有三类:myb、bhlh、wd40。其中wd40重复蛋白(wd40 repeat proteins,wdr)是一类具有β螺旋桨蛋白组结构的蛋白,其核心区域由40-60个氨基酸残基组成;该区域n端具有甘氨酸-组氨酸二肽(gly-his,gh)的重复序列,c端具有色氨酸-天冬氨酸二肽(trp-asp,wd)的重复序列,这种基序在同样的蛋白中一般可串联4-16次(simon et al.,1991;van and ludwig,2003)。重复的wd40基序可介导蛋白质之间的相互作用,在蛋白质互作时起固定作用(mishra et al.,2012),而利用酵母双杂交实验发现,与wd40蛋白相互作用的配体蛋白主要是myb和bhlh型转录因子(sompornpailin etal.,2002)。wd40蛋白参与了植物中的多种生理过程,主要包括植物非生物胁迫、生长发育和类黄酮的合成(walker et al.,1999;huang et al.,2008;miller et al.,2016)。
3、研究表明,矮牵牛的wd40重复蛋白由an11编码,包含有5个wd40重复基序,作用于an2的上游,调控矮牵牛花色素苷的合成(de vetten et al.,1997)。在拟南芥中,与矮牵牛an11转录因子高度同源的ttg1(transparent testa glabra 1),可与gl3相互作用,并与pap1形成三元复合体共同调控拟南芥花色素苷合成途径中结构基因的时空表达(brueggemann et al.,2010)。玉米的pac1编码wd40蛋白,pac1突变体中种子的糊粉层没有花色素苷的积累(carey et al.,2004)。
4、紫心甘薯作为花色素苷合成和积累部位特殊及具有重要开发价值的植物资源,用其进行非依光型及植物地下部分花色素苷合成的调控机制研究,在理论上可以丰富和深化植物花色素苷生物合成分子调控的基础理论;在应用上可以为紫心甘薯高花色素苷品种选育提供新的遗传标记,为其分子育种筛选出合适的操作元件或改造靶标,同时还可望为提高紫心甘薯块根中色素含量的栽培措施提供新的思路和线索。
技术实现思路
1、本发明要解决的技术问题在于筛选一种促进紫心甘薯ibwd40转录因子表达的上游调控因子。
2、为了解决上述技术问题,本发明首先用trizol法从甘薯块根提取rna,用smart技术逆转录合成双链的cdna,构建紫心甘薯酵母单杂交cdna文库。
3、进一步的,以紫心甘薯块根dna为模板,用takara高保真酶max dnapolymerase扩增两端带有不同酶切位点末端的ibwd40的启动子dna片段,并将启动子ibwd40构建到pabai载体中,扩增ibwd40的启动子dna片段的pcr引物对的具体序列如下所示:
4、pibwd40-f:5'-tcctagccaagaagagtggagaga-3'和
5、pibwd40-r:5'-tctcataccaccacaccctagtgg-3'。
6、构建的pabai-pibwd40诱饵载体经过自激活检测后,确定其自激活aba最低抑制浓度为300ng/ml。
7、本发明其次将诱饵菌株制成y1hgold感受态细胞,把文库质粒转入pabai-pibwd40诱感受态细胞中,通过酵母单杂交筛库对结合蛋白进行筛选,筛选得到了紫心甘薯ibwd40基因表达的上游调控因子为ibppa。
8、因此,本发明的第一个目的是提供一种紫心甘薯花色素苷合成调控因子ibppa,其氨基酸序列如seq id no.1所示。
9、本发明的第二个目的是提供一种上述的紫心甘薯花色素苷合成调控因子ibppa的编码基因,该编码基因的核苷酸序列如seq id no.2所示。
10、本发明的第三个目的是提供一种含有上述的编码基因的重组载体或重组菌。
11、本发明的第四个目的是提供一种含有上述的编码基因的表达盒。
12、本发明的第五个目的是提供一种上述的紫心甘薯花色素苷合成调控因子ibppa的扩增引物,该扩增引物的具体序列如下所示:
13、ibppa-f:5'-atggttccacctattgaaacacca-3'和
14、ibppa-r:5'-ctatcgcctcaggctttcca-3'。
15、本发明的第六个目的是提供上述的紫心甘薯花色素苷合成调控因子ibppa在促进甘薯ibwd40转录因子表达中的应用。
16、本发明的第七个目的是提供上述的紫心甘薯花色素苷合成调控因子ibppa在促进甘薯花色素苷生物合成中的应用。
17、本发明的第八个目的是提供上述的紫心甘薯花色素苷合成调控因子ibppa在甘薯高花色素苷品种选育中的应用。
18、本发明的第九个目的是提供一种促进甘薯花色素苷合成的方法,是在甘薯植株中过表达上述的调控因子ibppa。
19、进一步的,为了进一步验证筛选出来的调控因子ibppa与启动子ibwd40结合,将ibppa构建到pgadt7酵母重组表达载体,选择载体中ecorⅰ和bamhⅰ作为目的片段插入的酶切位点,合成引物序列见表2。将pgadt7-ibppa酵母重组表达载体质粒与pabai-pibwd40诱饵载体共同转化y1hgold酵母中进行酵母单杂交实验,结果发现:阳性对照p53abai+ad-53转化的菌株在sd/-leu/aba培养基上能够生长,而阴性对照pibwd40-1-pabai+pgadt7空载转化的菌株不能在sd/-leu/aba培养基上生长。说明该酵母单杂交实验能够有效检测蛋白是否结合在启动子上。而pibwd40-1-pabai+ibppa-pgadt7在sd/-leu/aba培养基上能够生长(图1),说明ibppa蛋白能结合在启动子ibwd40上。
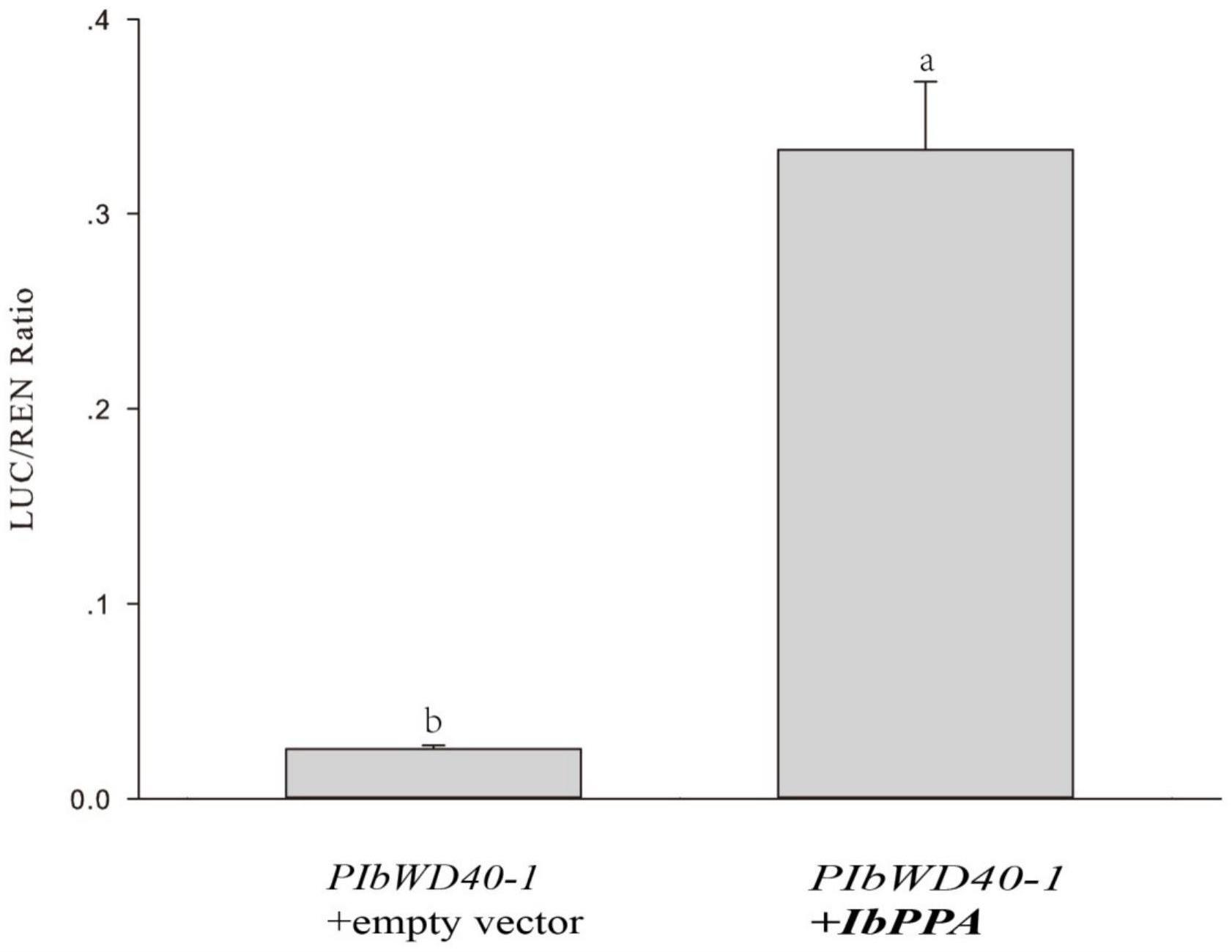
20、进一步的,为了验证启动子ibwd40与其调控因子ibppa的相互作用,本发明将ibppa构建到过表达载体pgreenii 0029 62-sk上,将pibwd40插入到载体pgreenii 0800-luc荧光素酶的前端作为报告质粒,选择pgreenii 0029 62-sk载体中sac i和xho i及pgreenii0800-luc载体中的kpn i和nco i作为插入目的片段的酶切位点,构建载体引物序列见表2。将测序成功的重组菌液提取质粒后与pibwd40+pgreenii 0800 luc重组质粒共同转入拟南芥原生质体中。结果显示ibppa能够提高ibwd40启动子的活性(图2),说明ibppa能够促进ibwd40的表达。
21、进一步的,为了明确上游调控因子ibppa的功能,本发明构建了ibppa与绿色荧光蛋白(gfp)的融合蛋白,对ibppa的作用场所进行定位。通过构建亚细胞定位表达载体,瞬时转化拟南芥原生质体后,利用激光共聚焦显微镜观察亚细胞定位情况,pcambia1300-gfp作为阳性对照。空载中的gfp蛋白能够在拟南芥原生质体的各个结构中表达,ibppa蛋白在细胞核中表达(图3),表明ibppa是一个典型的转录因子。
22、本发明的有益效果在于:通过酵母单杂交文库筛选实验,对启动子ibwd40的上游调控因子的筛选中成功获得了其上游调控因子ibppa。此发明结果在理论上可以丰富和深化植物花色素苷生物合成分子调控的基础理论;在应用上可以为紫心甘薯高花色素苷品种选育提供新的遗传标记,为其分子育种筛选出合适的操作元件或改造靶标,同时还可望为提高紫心甘薯块根中色素含量的栽培措施提供新的思路和线索。
- 还没有人留言评论。精彩留言会获得点赞!