一种产琥珀酸的大肠杆菌及其构建方法和应用与流程
1.本发明属于生物工程技术领域,具体涉及一种产琥珀酸的大肠杆菌及其构建方法和应用。
背景技术:
2.琥珀酸,被美国能源部列为十二种最有潜力大宗生物基化学品中的首位,在食品、化学、医药等领域有广泛的应用。生产琥珀酸的化学方法有石蜡氧化法、氯乙酸甲酯氰化水解法和五氧化二钒催化加氢法等,但是化学法生产琥珀酸的原料来自不可再生的石油资源。
3.为了摆脱以石油基为唯一原料生产化学品,可以选择植物来源的葡萄糖作为起始原料,利用工业微生物埃希氏菌发酵生产多种化学品,生物材料、药物中间体,例如氨基酸、有机酸、维生素等。因此,研究生物法生产琥珀酸,具有较好的应用前景。
4.然而,当以埃希氏菌株进行大规模发酵时,经常碰到噬菌体感染事件,扰乱了生产秩序,造成经济损失。当大肠杆菌感染噬菌体后,通常采用紫外光照射、甲醛熏蒸、菌种轮换等措施来保证生产的持续性。但这些应对措施耗时长、要求严格、成本高,无法从根本上解决噬菌体感染的问题。因此,筛选抗噬菌体的埃希氏起始菌株作为底盘细胞,对解决大规模发酵中噬菌体感染具有重要意义。
5.目前,以大肠杆菌为底盘细胞,通过基因工程改造宿主菌生产琥珀酸成为研究热点,例如张学礼等人改造的大肠杆菌hx024菌株,琥珀酸产量可达95.9g/l,刘立明等人改造的大肠杆菌fmme-suap菌株,琥珀酸产量达到80g/l。分析已发表的文献和公开的专利,我们发现起始菌株生产琥珀酸的过程中会产生乳酸、甲酸、乙酸等副产物,目的产物琥珀酸的生产效率低,,而且起始菌株及基因工程改造后的衍生菌株不具有抗噬菌体的能力,不适于工业化大规模生产。
技术实现要素:
6.为了解决上述技术问题,本发明从自然界筛选到一株抗噬菌体的埃希氏菌,以基因工程手段改造此埃希氏菌,通过敲除产生副产物的基因,获得一株抗噬菌体及高产琥珀酸的埃希氏菌株,具有重要的工业应用价值。
7.为实现上述目的,本发明采用的技术方案如下:
8.第一方面,本发明提供了一种抗噬菌体的埃希氏菌hb1,所述埃希氏菌hb1于2022年10月13日保藏于中国典型培养物保藏中心,保藏编号为cctcc no:m 20221565,保藏地址为中国湖北武汉,武汉大学。
9.第二方面,本发明提供了一种重组埃希氏菌的构建方法,对上述埃希氏菌hb1进行以下(1)-(4)中的一种或几种基因改造:
10.(1)敲除乳酸脱氢酶基因ldha;
11.(2)敲除醇脱氢酶基因adhe;
12.(3)敲除丙酮酸甲酸裂解酶基因pflb;
13.(4)敲除磷酸乙酰转移酶基因pta和/或乙酸激酶基因acka。
14.在本发明的一种实施方式中,利用red同源重组技术进行上述基因改造。
15.在本发明的一种实施方式中,所述基因改造包括:pcr扩增具有筛选与反筛选功能的cat-sacb片段;将cat-sacb片段整合到埃希氏菌hb1基因组中,筛选具有氯霉素抗性的转化子,并用pcr验证;筛选蔗糖不敏感转化子,完成目标基因敲除。
16.第三方面,本发明提供了利用上述重组埃希氏菌的构建方法制备得到的重组埃希氏菌。
17.在本发明的一种实施方式中,一种产琥珀酸的埃希氏菌hb332,通过对埃希氏菌hb1进行以下(1)-(4)的基因改造获得:
18.(1)敲除乳酸脱氢酶基因ldha;
19.(2)敲除醇脱氢酶基因adhe;
20.(3)敲除丙酮酸甲酸裂解酶基因pflb;
21.(4)敲除乙酸激酶基因acka。
22.第四方面,本发明提供了一种产琥珀酸的埃希氏菌hb333,所述埃希氏菌hb333于2022年10月13日保藏于中国典型培养物保藏中心,保藏编号为cctcc no:m 20221566,保藏地址为中国湖北武汉,武汉大学。
23.第五方面,本发明提供了上述埃希氏菌hb1或重组埃希氏菌或埃希氏菌hb333在生产琥珀酸中的应用。
24.在本发明的一种实施方式中,所述生产琥珀酸的方法包括:在发酵培养基上发酵培养埃希氏菌hb1或重组埃希氏菌或埃希氏菌hb333。
25.在本发明的一种实施方式中,所述发酵培养的方法为:将活化后的埃希氏菌hb1或重组埃希氏菌或埃希氏菌hb333以3-6%的接种量接种至发酵培养基中,30-40℃下进行厌氧发酵,发酵过程控制ph为7.0。
26.在本发明的一种实施方式中,发酵过程控制ph在7.0,所用中和剂为2.4mol/l碳酸钾和1.2mol/l氢氧化钾混合液(2.4m k2co3溶解在1.2m koh中)。
27.在本发明的一种实施方式中,所述发酵培养基为:含100g/l葡萄糖和100mm碳酸氢钾的无机盐培养基;
28.所述无机盐培养基为:nh4cl 1g
·
l-1
、kh2po
4 0.5g
·
l-1
、k2hpo
4 1.5g
·
l-1
、mgso40.2g
·
l-1
、nacl 1g
·
l-1
、微量元素溶液2ml/l,ph7.0;
29.所述微量元素溶液为:cocl2·
6h2o 0.1g
·
l-1
、mncl2·
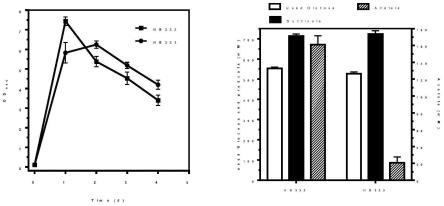
4h2o 0.425g
·
l-1
、zncl20.05g
·
l-1
、nicl2·
6h2o 0.01g
·
l-1
、cuso4·
5h2o 0.015g
·
l-1
、na2moo4·
2h2o 0.01g
·
l-1
、na2seo4·
2h2o 0.01g
·
l-1
。
30.在本发明的一种实施方式中,所述埃希氏菌hb1或重组埃希氏菌或埃希氏菌hb333在种子培养基中活化,具体步骤包括:将埃希氏菌hb1、埃希氏菌hb332或埃希氏菌hb333单克隆接种到种子培养基中,37℃,180r
·
min-1
振荡培养培养12h,获得种子液,用于发酵培养基接种;
31.其中,种子培养基为含50g/l葡萄糖和100mm碳酸氢钾的无机盐培养基;
32.所述无机盐培养基为:nh4cl 1g
·
l-1
、kh2po
4 0.5g
·
l-1
、k2hpo
4 1.5g
·
l-1
、
mgso40.2g
·
l-1
、nacl 1g
·
l-1
、微量元素溶液2ml/l,ph7.0;
33.所述微量元素溶液为:cocl2·
6h2o 0.1g
·
l-1
、mncl2·
4h2o 0.425g
·
l-1
、zncl20.05g
·
l-1
、nicl2·
6h2o 0.01g
·
l-1
、cuso4·
5h2o 0.015g
·
l-1
、na2moo4·
2h2o 0.01g
·
l-1
、na2seo4·
2h2o 0.01g
·
l-1
。
34.在本发明的一种实施方式中,所述埃希氏菌hb1或重组埃希氏菌或埃希氏菌hb333在种子培养基中活化,具体步骤包括:
35.(a)将埃希氏菌hb1或重组埃希氏菌或埃希氏菌hb333单克隆在一级种子培养基中30-39℃,160-200rpm振荡培养7-12h,获得一级种子液;
36.(b)将一级种子液以1-3%的接种量接种到二级种子培养基中30-39℃,160-200rpm振荡培养13-18h,获得二级种子液,用于发酵培养基接种。
37.在本发明的一种实施方式中,所述一级种子培养基为含有50g/l葡萄糖、50mm khco3和100mm mops的无机盐培养基;
38.所述二级种子培养基为含有100g/l葡萄糖、100mm碳酸氢钾和1mm甜菜碱的无机盐培养基;
39.所述无机盐培养基为:nh4cl 1g
·
l-1
、kh2po
4 0.5g
·
l-1
、k2hpo
4 1.5g
·
l-1
、mgso40.2g
·
l-1
、nacl 1g
·
l-1
、微量元素溶液2ml/l,ph7.0;
40.所述微量元素溶液为:cocl2·
6h2o 0.1g
·
l-1
、mncl2·
4h2o 0.425g
·
l-1
、zncl241.0.05g
·
l-1
、nicl2·
6h2o 0.01g
·
l-1
、cuso4·
5h2o 0.015g
·
l-1
、na2moo4·
2h2o 0.01g
·
l-1
、na2seo4·
2h2o 0.01g
·
l-1
。
42.在本发明的一种实施方式中,将冷藏保存的埃希氏菌hb1或重组埃希氏菌或埃希氏菌hb333在含有2%葡萄糖的无机盐固体培养基上划线,平板倒扣放置在温度为37℃的培养箱中培养24h,挑取单克隆,所述单克隆用于接种至一级种子培养基中培养。
43.在本发明的一种实施方式中,所述无机盐固体培养基为:在无机盐培养基基础上添加琼脂粉15g
·
l-1
。
44.与现有技术相比,本发明具有下述有益技术效果:
45.1、本发明利用传统微生物分离手段,筛选了一种抗噬菌体的埃希氏菌hb1,该菌株可以发酵生产琥珀酸;
46.2、本发明以抗噬菌体的埃希氏菌hb1为出发菌,利用基因编辑技术敲除ldha基因、pta基因、pflb基因和adhe基因,获得埃希氏菌hb333,该埃希氏菌hb333可利用价廉易得的葡萄糖为原料,通过厌氧发酵生产琥珀酸,其在96h内产生85.4g/l的琥珀酸,葡萄糖转化率可达90%,显著提高了琥珀酸的生产效率,而且埃希氏菌hb333可以耐受高浓度葡萄糖,发酵过程中无需补加葡萄糖,也无需添加酵母膏,降低了生产成本;该埃希氏菌hb333发酵过程还降低了乳酸、乙酸、甲酸、乙醇等副产物的产生,大大减轻了下游琥珀酸产品的纯化压力;同时埃希氏菌hb333具有出发菌的抗噬菌体特性,解决了发酵生产琥珀酸中易发生噬菌体感染的问题,更适于大规模工业化生产。
附图说明
47.图1为噬菌体抗性检测的双层琼脂固体培养基接种埃希氏菌hb1(a)和对照菌株大肠杆菌dh5α(b)的菌落形态电镜图。
48.图2为埃希氏菌hb1的形态照片,其中a为菌落形态,b和c为扫描电镜拍摄的单个细菌形态。
49.图3为埃希氏菌hb1的16srdna扩增后的核酸凝胶电泳图。
50.图4为埃希氏菌hb1的系统进化树。
51.图5为埃希氏菌hb1的生长曲线。
52.图6为埃希氏菌hb1(a)及衍生菌株hb73(b)和hb74(c)的厌氧发酵结果。
53.图7为埃希氏菌hb332与hb333的厌氧发酵结果。
54.图8为利用red同源重组技术对埃希氏菌hb1进行基因敲除改造的流程图;
55.图9为琥珀酸标准品的液相色谱图;
56.图10为埃希氏菌hb333发酵96h琥珀酸发酵样品液相色谱图。
具体实施方式
57.本发明通过下述实施例进一步阐明,但任何实施例或其组合不应当理解为对本发明的范围或实施方式的限制。本发明的范围由所附权利要求书限定,结合本说明书和本领域一般常识,本领域普通技术人员可以清楚地明白权利要求书所限定的范围。在不偏离本发明的精神和范围的前提下,本领域技术人员可以对本发明的技术方案进行任何修改或改变,这种修改和改变也包含在本发明的范围内。
58.下述实施例中所使用的实验方法如无特殊说明,均为常规方法;所用的试剂和材料等,如无特殊说明,均可从商业途径获得。
59.一、实验材料
60.耐噬菌体菌株分离的土壤样品取自浙江省杭州市临安区指南村餐厨垃圾有机肥堆放地的土壤。
61.琥珀酸(99%纯度)购自阿拉丁(aladdin)公司,其cas:110-15-6;
62.氯霉素(chloramphenicol)购于阿拉丁(aladdin)公司,其cas:56-75-7;
63.氨苄青霉素购于阿拉丁(aladdin)公司,其cas:7177-48-2。
64.丰富培养基:
65.lb液体培养基:胰蛋白胨10g
·
l-1
、酵母提取物5g
·
l-1
、氯化钠10g
·
l-1
,使用2mol
·
l-1
hcl溶液或2mol
·
l-1
naoh溶液调节ph值至7.0;
66.lb固体培养基:在lb液体培养基基础上添加琼脂粉15g
·
l-1
;
67.发酵培养基:含100g/l葡萄糖和100mm碳酸氢钾的无机盐培养基;
68.种子培养基:含50g/l葡萄糖和100mm碳酸氢钾的无机盐培养基;
69.无机盐培养基:
70.nh4cl 1g
·
l-1
、kh2po
4 0.5g
·
l-1
、k2hpo
4 1.5g
·
l-1
、mgso
4 0.2g
·
l-1
、nacl 1g
·
l-1
、微量元素溶液2ml/l,ph7.0;
71.微量元素溶液:cocl2·
6h2o 0.1g
·
l-1
、mncl2·
4h2o 0.425g
·
l-1
、zncl
2 0.05g
·
l-1
、nicl2·
6h2o 0.01g
·
l-1
、cuso4·
5h2o 0.015g
·
l-1
、na2moo4·
2h2o 0.01g
·
l-1
、na2seo4·
2h2o0.01g
·
l-1
。
72.无机盐固体培养基:在无机盐培养基基础上添加琼脂粉15g
·
l-1
。
73.二、检测方法
74.1、高效液相色谱(hplc)测定琥珀酸、乳酸、乙酸、甲酸、葡萄糖含量:
75.琥珀酸、乳酸、乙酸、甲酸、葡萄糖含量测定:选用高效液相色谱仪器是agilent1260(美国安捷伦公司),配置bio-rad公司的有机酸色谱柱aminex hpx-87h(300
×
7.8mm,5μm),配置uv检测器210nm;色谱分析条件如下:流动相是5mm硫酸溶液,柱温是40℃,流速是0.5ml
·
min-1
,进样量是10μl。根据所测得的高效液相色谱出峰面积与对应标准品浓度构建线性回归方程,通过所得线性回归方程计算各有机酸和葡萄糖的浓度。
76.2、气相色谱测定乙醇含量:使用安捷伦6890n气相色谱仪,配备火焰电离检测器和15-m hp plot q megabore色谱柱(zheng hb.et al,appl.environ.microbiol.2012,78(12):4346)。
77.3、耐噬菌体菌株的活性检测
78.利用双层琼脂平板法进行噬菌体检测。首先,融化下层琼脂培养基,倒平板(约10ml/皿),待用;第二步,融化上层琼脂培养基,冷却至50℃,每管中加入0.2ml待测试的菌株及对照菌株(大肠杆菌dh5α),再加入0.2ml噬菌体裂解液,混合后立即倒入上层平板铺平(约5ml/皿),37℃,培养6~12h。由于对照菌株不耐噬菌体,在双层培养基的上层出现圆形透亮的空斑噬菌斑。
79.三、计算方法
80.1、琥珀酸生产强度(g/l/h)=琥珀酸产量(g/l)/发酵时间(h);
81.2、葡萄糖转化率=琥珀酸产量(g/l)/葡萄糖消耗量(g/l)
82.实施例1耐噬菌体菌株(埃希氏菌hb1)的筛选和鉴定
83.1、耐噬菌体菌株hb1的分离和纯化
84.向装有100ml的无机盐培养基的250ml锥形瓶中(灭菌后使用),加入土壤样品10g、实验室保存的噬菌体菌液100μl、酵母提取物0.2g、葡萄糖0.1g,在37℃,180r
·
min-1
振荡培养2d后,补加噬菌体菌液100μl,37℃,180r
·
min-1
振荡培养2d;将培养液以10%接种量转接至含100ml无机盐培养基的锥形瓶中,再加入噬菌体菌液100μl、酵母提取物0.1g和葡萄糖0.05g,37℃,180r
·
min-1
振荡培养24d;将经过三轮驯化的菌液涂在含有50μl噬菌体菌液的lb固体培养基上,放入37℃恒温培养箱避光培养16h,挑取单菌落,转接至新lb固体培养基上,经历3次分离纯化,获得耐噬菌体的单一菌株,命名为hb1。
85.2、耐噬菌体菌株hb1的保存与鉴定
86.a、耐噬菌体菌株hb1的保存
87.将获得的单一菌株hb1接种到lb液体培养基中,37℃,180r
·
min-1
振荡培养16h后,在无菌操作台中取0.5ml菌液加入到事先灭菌的装有0.5ml40%甘油的离心保存管中,用胶带封口之后放入-80℃超低温冰箱保存。
88.b、耐噬菌体菌株hb1的形态学特征
89.菌株hb1在lb固体培养基上,37℃恒温、避光培养16h,菌落形态如图1a所示,菌落呈圆形、白色不透明并且边缘整齐表面有黏稠性,挑取单一菌落送至浙江省农业科学研究院测试中心进行电子显微镜观察,如图2b和2c所示,单个细菌在电镜下为短杆状、无芽孢、有鞭毛。
90.c、耐噬菌体菌株hb1的分子生物学鉴定
91.挑取对数生长期的耐噬菌体菌株hb1,送至杭州有康生物公司进行测序:扩增模板
为细菌的总dna,细菌的16s rdna的pcr扩增引物为通用引物(正向引物27f,5
′‑
agagtttgatcctggctcag-3
′
;反向引物1492r,5
′‑
tacggctaccttgttacg actt-3
′
),其16srdna序列如seq id no:1所示;其16srdna扩增后的核酸凝胶电泳图如图3所示。
92.测序完成后将获得序列通过blast与genbank中已知序列进行同源性比较,确定与耐噬菌体菌株hb1同源性程度高的序列,基于获得的耐噬菌体菌株hb1的16s rdna基因序列(1439bp),使用mega 7.0软件对耐噬菌体菌株hb1及其近缘属种进行遗传距离分析,系统发育进化树的构建采用邻接法进行,系统发育进化树如图4所示。
93.耐噬菌体菌株hb1与埃希氏菌(escherichia)为同一类群,与埃希氏菌k-12(escherichia k-12(nc000913.3)亲缘关系最近,结合扫描电镜以及其他表征,将耐噬菌体菌株hb1鉴定为埃希氏菌属,命名为埃希氏菌hb1。
94.埃希氏菌hb1已经于2022年10月13日保藏于中国典型培养物保藏中心(cctcc),地址为中国湖北武汉,武汉大学,保藏编号为cctcc no:m 20221565。
95.如图8所示,本发明采用两步法同源重组技术敲除或替换基因组中目的基因,没有残留外源片段。利用同源重组技术在埃希氏菌基因组中进行遗传操作,引入氯霉素抗性基因以及利用蔗糖致死的反向筛选需要cat-sacb大片段参与。cat-sacb大片段由擎科生物全基因合成,与载体pcr2.1-topo(invitrogen公司)连接,形成质粒poh5899(图8a),利用菌落pcr和基因测序技术确认质粒正确性,cat-sacb核苷酸序列如seq id no:2所示。
96.下述实施例中所有基因的敲除采用的相同步骤,如下:
97.第一步、导入cat-sacb大片段替换目的基因,从而引入氯霉素抗性基因cat和蔗糖致死基因sacb;具体步骤如下:
98.1.1)设计长引物(同源臂序列50bp加上配对序列26bp),以质粒poh5899为模板,以表1所示的配对长引物进行pcr扩增,获得约3.44kb的大片段,也即是cat-sacb片段,加入dpni进行酶切消除质粒模板,用试剂盒纯化pcr产物;
99.1.2)将步骤1.1)所得pcr产物通过电转化导入含pkd46质粒的埃希氏菌hb1感受态细胞内,通过氯霉素抗性筛选,所用抗性筛选培养基为:含有40mg
·
l-1
氯霉素、及50g
·
l-1
阿拉伯糖(用作λred重组酶的诱导剂;gene bridges)或100g
·
l-1
蔗糖(用于sacb的反向选择)的lb液体培养基,在39℃、180r
·
min-1
条件下振动培养12-16小时;pcr验证,获得基因组上目标基因被cat-sacb大片段替换成功的菌株,命名为hb1衍生菌株。
100.第二步、反向筛选的同源重组,sacb基因编码果聚糖蔗糖转移酶,催化蔗糖水解成葡萄糖和果糖,果糖聚合成高分子量的果聚糖,此化合物对细胞有毒性,造成细胞死亡。具体步骤如下:
101.2.1)利用重叠pcr方法将目标基因上下游序列的同源臂dna片段连接在一起,长度约1.0kb;
102.2.2)将步骤2.1)所得pcr产物纯化后通过电转化导入含pkd46质粒的hb1衍生菌株感受态细胞内,在三角摇瓶内进行菌株复苏和筛选,所用培养基为不含nacl、含10%蔗糖的lb液体培养基,在30℃、180r
·
min-1
条件下振动培养12-16小时,待菌体浓度较高时,在lb固体培养基上划线培养,挑取单克隆,影印到含6%蔗糖的lb固体培养基上,重复验证蔗糖敏感性,最终获得发生第二步重组的菌株。
103.下述实施例中,以埃希氏菌hb1为出发菌株,也可利用埃希氏菌hb1衍生菌株或其
他修饰后可以生产琥珀酸的大肠杆菌作为出发菌株。
104.实施例2埃希氏菌hb73的构建
105.对筛选的埃希氏菌hb1进行发酵生产琥珀酸,结果发下发酵液中乙酸含量较高(如图6所示)。以埃希氏菌hb1作为出发菌株,根据大肠杆菌基因组注释,磷酸乙酰转移酶基因pta与乙酸激酶基因acka基因在一个操纵子内,两个基因表达的酶依次催化乙酰辅酶a生成乙酸。为了减少副产物乙酸的积累,在构建琥珀酸菌的过程中,采用red同源重组技术在埃希氏菌hb1基因组中敲除了造成乙酸积累的乙酸激酶基因acka,获得hb73菌株,hb73菌株经鉴定为埃希氏菌属,命名为埃希氏菌hb73。
106.埃希氏菌hb73的具体构建步骤如下:
107.2.1、以质粒poh5899为模板,以表1所示的配对长引物对acka-5899-f/acka-5899-r进行pcr扩增,获得约3.44kb的大片段,加入dpni进行酶切消除质粒模板,用试剂盒纯化pcr产物;
108.2.2、将步骤2.1所得pcr产物通过电转化导入含pkd46质粒的埃希氏菌hb1感受态细胞内,通过氯霉素抗性筛选,所用抗性筛选培养基为:含有40mg
·
l-1
氯霉素、及50g
·
l-1
阿拉伯糖(用作λred重组酶的诱导剂;gene bridges)或100g
·
l-1
蔗糖(用于sacb的反向选择)的lb液体培养基,在39℃、180r
·
min-1
条件下振动培养12-16小时;pcr验证,获得基因组上目标基因acka被cat-sacb大片段替换成功的菌株,命名为hb1衍生菌株1。
109.2.3、利用重叠pcr方法将乙酸激酶基因acka上下游序列的同源臂dna片段连接在一起,长度约1.0kb;
110.其中,乙酸激酶基因acka上游序列的同源臂dna通过引物acka-up-f/acka-up-r扩增,下游序列的同源臂dna通过引物acka-dn-f/acka-dn-r扩增;以扩增的acka上下游序列的同源臂dna为共同的模板,以引物acka-up-f和acka-dn-r为引物,扩增acka上下游序列的同源臂dna拼接片段。
111.2.4、将步骤2.4.3所得pcr产物(acka上下游序列的同源臂dna拼接片段)纯化后通过电转化导入含pkd46质粒的hb1衍生菌株1感受态细胞内,在三角摇瓶内进行菌株复苏和筛选,所用培养基为不含nacl、含10%蔗糖的lb液体培养基,在30℃、180r
·
min-1
条件下振动培养12-16小时,待菌体浓度较高时,在lb固体培养基上划线培养,挑取单克隆,影印到含6%蔗糖的lb固体培养基上,重复验证蔗糖敏感性,最终获得敲除乙酸激酶基因acka的埃希氏菌菌株hb1-δacka,命名为埃希氏菌hb73。
112.实施例3埃希氏菌hb73的构建
113.对改造的筛选的埃希氏菌hb73进行发酵生产琥珀酸,结果发下发酵液中乙酸含量显著降低(如图6所示)。为了降低乙酸含量,本发明又以埃希氏菌hb1作为出发菌株,采用red同源重组技术在埃希氏菌hb1基因组中敲除了磷酸乙酰转移酶基因pta,获得hb74菌株,hb74菌株经鉴定为埃希氏菌属,命名为埃希氏菌hb74。
114.埃希氏菌hb74的具体构建步骤如下:
115.3.1、以质粒poh5899为模板,以表1所示的配对长引物对pta-5899-f/pta-5899-r进行pcr扩增,获得约3.44kb的大片段,加入dpni进行酶切消除质粒模板,用试剂盒纯化pcr产物;
116.3.2、将步骤3.1所得pcr产物通过电转化导入含pkd46质粒的埃希氏菌hb1感受态
细胞内,通过氯霉素抗性筛选,所用抗性筛选培养基为:含有40mg
·
l-1
氯霉素、及50g
·
l-1
阿拉伯糖(用作λred重组酶的诱导剂;gene bridges)或100g
·
l-1
蔗糖(用于sacb的反向选择)的lb液体培养基,在39℃、180r
·
min-1
条件下振动培养12-16小时;pcr验证,获得基因组上目标基因pta被cat-sacb大片段替换成功的菌株,命名为hb1衍生菌株2。
117.3.3、利用重叠pcr方法将磷酸乙酰转移酶基因pta上下游序列的同源臂dna片段连接在一起,长度约1.0kb;
118.其中,磷酸乙酰转移酶基因pta上游序列的同源臂dna通过引物pta-up-f/pta-up-r扩增,下游序列的同源臂dna通过引物pta-dn-f/pta-dn-r扩增;以扩增的pta上下游序列的同源臂dna为共同的模板,以引物pta-up-f和pta-dn-r为引物,扩增pta上下游序列的同源臂dna拼接片段。
119.3.4、将步骤3.3所得pcr产物(pta上下游序列的同源臂dna拼接片段)纯化后通过电转化导入含pkd46质粒的hb1衍生菌株2感受态细胞内,在三角摇瓶内进行菌株复苏和筛选,所用培养基为不含nacl、含10%蔗糖的lb液体培养基,在30℃、180r
·
min-1
条件下振动培养12-16小时,待菌体浓度较高时,在lb固体培养基上划线培养,挑取单克隆,影印到含6%蔗糖的lb固体培养基上,重复验证蔗糖敏感性,最终获得敲除磷酸乙酰转移酶基因pta的埃希氏菌菌株hb1-δpta,命名为埃希氏菌hb74。
120.实施例4埃希氏菌hb332的构建
121.本实施例中,以埃希氏菌hb1作为出发菌株,根据大肠杆菌基因组注释,磷酸乙酰转移酶基因pta与乙酸激酶基因acka基因在一个操纵子内,两个基因表达的酶依次催化乙酰辅酶a生成乙酸。在构建琥珀酸菌的过程中,采用red同源重组技术在埃希氏菌hb1基因组中依次敲除了造成副产物积累的基因,包含甲酸代谢途径的丙酮酸甲酸裂解酶基因pflb、乳酸代谢途径的乳酸脱氢酶基因ldha、乙醇代谢途径的乙醇脱氢酶基因adhe和乙酸代谢途径的乙酸激酶基因acka,获得hb332菌株,hb332菌株经鉴定为埃希氏菌属,命名为埃希氏菌hb332。
122.4.1、敲除丙酮酸甲酸裂解酶基因pflb
123.4.1.1、以质粒poh5899为模板,以表1所示的配对长引物对pflb-5899-f/pflb-5899-r进行pcr扩增,获得约3.44kb的大片段,加入dpni进行酶切消除质粒模板,用试剂盒纯化pcr产物;
124.4.1.2、将步骤4.1.1所得pcr产物通过电转化导入含pkd46质粒的埃希氏菌hb1感受态细胞内,通过氯霉素抗性筛选,所用抗性筛选培养基为:含有40mg
·
l-1
氯霉素、及50g
·
l-1
阿拉伯糖(用作λred重组酶的诱导剂;gene bridges)或100g
·
l-1
蔗糖(用于sacb的反向选择)的lb液体培养基,在39℃、180r
·
min-1
条件下振动培养12-16小时;pcr验证,获得基因组上目标基因pflb被cat-sacb大片段替换成功的菌株,命名为hb1衍生菌株。
125.4.1.3、利用重叠pcr方法将丙酮酸甲酸裂解酶基因pflb上下游序列的同源臂dna片段连接在一起,长度约1.0kb;
126.其中,丙酮酸甲酸裂解酶基因pflb上游序列的同源臂dna通过引物pflb-up-f/pflb-up-r扩增,下游序列的同源臂dna通过引物pflb-dn-f/pflb-dn-r扩增;以扩增的pflb上下游序列的同源臂dna为共同的模板,以pflb-up-f和pflb-dn-r为引物,扩增pflb上下游序列的同源臂dna拼接片段。
127.4.1.4、将步骤4.1.3所得pcr产物(pflb上下游序列的同源臂dna拼接片段)纯化后通过电转化导入含pkd46质粒的hb1衍生菌株感受态细胞内,在三角摇瓶内进行菌株复苏和筛选,所用培养基为不含nacl、含10%蔗糖的lb液体培养基,在30℃、180r
·
min-1
条件下振动培养12-16小时,待菌体浓度较高时,在lb固体培养基上划线培养,挑取单克隆,影印到含6%蔗糖的lb固体培养基上,重复验证蔗糖敏感性,最终获得敲除丙酮酸甲酸裂解酶基因pflb的埃希氏菌菌株,命名为埃希氏菌hb1-δpflb。
128.4.2、敲除乳酸脱氢酶基因ldha
129.4.4.1、以质粒poh5899为模板,以表1所示的配对长引物对ldha-5899-f/ldha-5899-r进行pcr扩增,获得约3.44kb的大片段,加入dpni进行酶切消除质粒模板,用试剂盒纯化pcr产物;
130.4.4.2、将步骤4.4.1所得pcr产物通过电转化导入含pkd46质粒的埃希氏菌hb1-δpflb感受态细胞内,通过氯霉素抗性筛选,所用抗性筛选培养基为:含有40mg
·
l-1
氯霉素、及50g
·
l-1
阿拉伯糖(用作λred重组酶的诱导剂;gene bridges)或100g
·
l-1
蔗糖(用于sacb的反向选择)的lb液体培养基,在39℃、180r
·
min-1
条件下振动培养12-16小时;pcr验证,获得基因组上目标基因ldha被cat-sacb大片段替换成功的菌株,命名为hb1-δpflb衍生菌株。
131.4.4.3、利用重叠pcr方法将乳酸脱氢酶基因ldha上下游序列的同源臂dna片段连接在一起,长度约1.0kb;
132.其中,乳酸脱氢酶基因ldha上游序列的同源臂dna通过引物ldha-up-f/ldha-up-r扩增,下游序列的同源臂dna通过引物ldha-dn-f/ldha-dn-r扩增;以扩增的ldha上下游序列的同源臂dna为共同的模板,以引物ldha-up-f和ldha-dn-r为引物,扩增ldha上下游序列的同源臂dna拼接片段。
133.4.4.4、将步骤4.4.3所得pcr产物(ldha上下游序列的同源臂dna拼接片段)纯化后通过电转化导入含pkd46质粒的hb1-δpflb衍生菌株感受态细胞内,在三角摇瓶内进行菌株复苏和筛选,所用培养基为不含nacl、含10%蔗糖的lb液体培养基,在30℃、180r
·
min-1
条件下振动培养12-16小时,待菌体浓度较高时,在lb固体培养基上划线培养,挑取单克隆,影印到含6%蔗糖的lb固体培养基上,重复验证蔗糖敏感性,最终获得敲除丙酮酸甲酸裂解酶基因pflb和乳酸脱氢酶基因ldha的埃希氏菌菌株,命名为埃希氏菌hb1-δpflb-δldha。
134.4.3、敲除乙醇脱氢酶基因adhe
135.4.3.1、以质粒poh5899为模板,以表1所示的配对长引物对adhe-5899-f/adhe-5899-r进行pcr扩增,获得约3.44kb的大片段,加入dpni进行酶切消除质粒模板,用试剂盒纯化pcr产物;
136.4.3.2、将步骤4.3.1所得pcr产物通过电转化导入含pkd46质粒的埃希氏菌hb1-δpflb-δldha感受态细胞内,通过氯霉素抗性筛选,所用抗性筛选培养基为:含有40mg
·
l-1
氯霉素、及50g
·
l-1
阿拉伯糖(用作λred重组酶的诱导剂;gene bridges)或100g
·
l-1
蔗糖(用于sacb的反向选择)的lb液体培养基,在39℃、180r
·
min-1
条件下振动培养12-16小时;pcr验证,获得基因组上目标基因adhe被cat-sacb大片段替换成功的菌株,命名为hb1-δpflb-δldha衍生菌株。
137.4.3.3、利用重叠pcr方法将乙醇脱氢酶基因adhe上下游序列的同源臂dna片段连
接在一起,长度约1.0kb;
138.其中,乙醇脱氢酶基因adhe上游序列的同源臂dna通过引物adhe-up-f/adhe-up-r扩增,下游序列的同源臂dna通过引物adhe-dn-f/adhe-dn-r扩增;以扩增的adhe上下游序列的同源臂dna为共同的模板,以引物adhe-up-f和adhe-dn-r为引物,扩增adhe上下游序列的同源臂dna拼接片段。
139.4.3.4、将步骤4.3.3所得pcr产物(adhe上下游序列的同源臂dna拼接片段)纯化后通过电转化导入含pkd46质粒的hb1-δpflb-δldha衍生菌株感受态细胞内,在三角摇瓶内进行菌株复苏和筛选,所用培养基为不含nacl、含10%蔗糖的lb液体培养基,在30℃、180r
·
min-1
条件下振动培养12-16小时,待菌体浓度较高时,在lb固体培养基上划线培养,挑取单克隆,影印到含6%蔗糖的lb固体培养基上,重复验证蔗糖敏感性,最终获得敲除丙酮酸甲酸裂解酶基因pflb、乳酸脱氢酶基因ldha和乙醇脱氢酶基因adhe的埃希氏菌菌株,命名为埃希氏菌hb1-δpflb-δldha-δadhe。
140.4.4、敲除乙酸激酶基因acka
141.4.4.1、以质粒poh5899为模板,以表1所示的配对长引物对acka-5899-f/acka-5899-r进行pcr扩增,获得约3.44kb的大片段,加入dpni进行酶切消除质粒模板,用试剂盒纯化pcr产物;
142.4.4.2、将步骤4.4.1所得pcr产物通过电转化导入含pkd46质粒的埃希氏菌hb1-δpflb-δldha-δadhe感受态细胞内,通过氯霉素抗性筛选,所用抗性筛选培养基为:含有40mg
·
l-1
氯霉素、及50g
·
l-1
阿拉伯糖(用作λred重组酶的诱导剂;gene bridges)或100g
·
l-1
蔗糖(用于sacb的反向选择)的lb液体培养基,在39℃、180r
·
min-1
条件下振动培养12-16小时;pcr验证,获得基因组上目标基因acka被cat-sacb大片段替换成功的菌株,命名为hb1-δpflb-δldha-δadhe衍生菌株。
143.4.4.3、利用重叠pcr方法将乙酸激酶基因acka上下游序列的同源臂dna片段连接在一起,长度约1.0kb;
144.其中,乙酸激酶基因acka上游序列的同源臂dna通过引物acka-up-f/acka-up-r扩增,下游序列的同源臂dna通过引物acka-dn-f/acka-dn-r扩增;以扩增的acka上下游序列的同源臂dna为共同的模板,以引物acka-up-f和acka-dn-r为引物,扩增acka上下游序列的同源臂dna拼接片段。
145.4.4.4、将步骤4.4.3所得pcr产物(acka上下游序列的同源臂dna拼接片段)纯化后通过电转化导入含pkd46质粒的hb1-δpflb-δldha-δadhe衍生菌株感受态细胞内,在三角摇瓶内进行菌株复苏和筛选,所用培养基为不含nacl、含10%蔗糖的lb液体培养基,在30℃、180r
·
min-1
条件下振动培养12-16小时,待菌体浓度较高时,在lb固体培养基上划线培养,挑取单克隆,影印到含6%蔗糖的lb固体培养基上,重复验证蔗糖敏感性,最终获得敲除丙酮酸甲酸裂解酶基因pflb、乳酸脱氢酶基因ldha、乙醇脱氢酶基因adhe和乙酸激酶基因acka的埃希氏菌菌株hb1-δpflb-δldha-δadhe-δacka,命名为埃希氏菌hb332。
146.实施例5埃希氏菌hb333的构建
147.为了降低乙酸产量,我们在埃希氏菌hb1-δpflb-δldha-δadhe基础上敲除磷酸乙酰转移酶基因pta,获得hb333菌株,hb333菌株经鉴定为埃希氏菌属,命名为埃希氏菌hb333。
148.5.1、以质粒poh5899为模板,以表1所示的配对长引物对pta-5899-f/pta-5899-r进行pcr扩增,获得约3.44kb的大片段,加入dpni进行酶切消除质粒模板,用试剂盒纯化pcr产物;
149.5.2、将步骤5.1所得pcr产物通过电转化导入含pkd46质粒的埃希氏菌hb1-δpflb-δldha-δadhe感受态细胞内,通过氯霉素抗性筛选,所用抗性筛选培养基为:含有40mg
·
l-1
氯霉素、及50g
·
l-1
阿拉伯糖(用作λred重组酶的诱导剂;gene bridges)或100g
·
l-1
蔗糖(用于sacb的反向选择)的lb液体培养基,在39℃、180r
·
min-1
条件下振动培养12-16小时;pcr验证,获得基因组上目标基因pta被cat-sacb大片段替换成功的菌株,命名为hb1-δpflb-δldha-δadhe衍生菌株。
150.5.3、利用重叠pcr方法将磷酸乙酰转移酶基因pta上下游序列的同源臂dna片段连接在一起,长度约1.0kb;
151.其中,磷酸乙酰转移酶基因pta上游序列的同源臂dna通过引物pta-up-f/pta-up-r扩增,下游序列的同源臂dna通过引物pta-dn-f/pta-dn-r扩增;以扩增的pta上下游序列的同源臂dna为共同的模板,以引物pta-up-f和pta-dn-r为引物,扩增pta上下游序列的同源臂dna拼接片段。
152.5.4、将步骤5.3所得pcr产物(pta上下游序列的同源臂dna拼接片段)纯化后通过电转化导入含pkd46质粒的hb1-δpflb-δldha-δadhe衍生菌株感受态细胞内,在三角摇瓶内进行菌株复苏和筛选,所用培养基为不含nacl、含10%蔗糖的lb液体培养基,在30℃、180r
·
min-1
条件下振动培养12-16小时,待菌体浓度较高时,在lb固体培养基上划线培养,挑取单克隆,影印到含6%蔗糖的lb固体培养基上,重复验证蔗糖敏感性,最终获得敲除丙酮酸甲酸裂解酶基因pflb、乳酸脱氢酶基因ldha、乙醇脱氢酶基因adhe和乙酸激酶基因acka的埃希氏菌菌株hb1-δpflb-δldha-δadhe-δpta,命名为埃希氏菌hb333。
153.埃希氏菌hb333已经于2022年10月13日保藏于中国典型培养物保藏中心(cctcc),地址为中国湖北武汉,武汉大学,保藏编号为cctcc no:m 20221566。
154.表1本发明中使用的引物
[0155][0156]
[0157]
实施例6埃希氏菌hb1、埃希氏菌hb73、埃希氏菌hb74、埃希氏菌hb332和埃希氏菌hb333发酵生产琥珀酸
[0158]
(1)挑单克隆:将冷藏保存的埃希氏菌hb1或埃希氏菌hb73、埃希氏菌hb74、埃希氏菌hb332或埃希氏菌hb333在含有2%葡萄糖的无机盐固体培养基上划线,平板倒扣放置在温度为37℃的培养箱中培养24h,挑取单克隆,用于接种至种子培养基。
[0159]
(2)种子培养:将挑取的新鲜埃希氏菌hb1、埃希氏菌hb73、埃希氏菌hb74、埃希氏菌hb332和埃希氏菌hb333单克隆接种到种子培养基中,37℃,180r
·
min-1
振荡培养培养12h,获得种子液。
[0160]
(3)发酵培养:将种子液按5%接种量接种于装有3l发酵培养基的发酵罐中,37℃,300rpm条件下发酵培养,培养过程中不通任何气体,使用混合碱液(2.4m k2co3溶解在1.2m koh中)调控发酵罐的ph在7.0。
[0161]
(4)发酵96h停止发酵,分别检测发酵液中葡萄糖和琥珀酸的浓度以及乳酸、甲酸、乙酸、乙醇含量。
[0162]
如图6所示,埃希氏菌hb1(6a)在厌氧发酵4天内琥珀酸产量为49.3mm,同时有甲酸、乙酸、乳酸和乙醇等副产物的产生;
[0163]
在埃希氏菌hb1基因组中敲除乙酸激酶基因acka获得的hb73菌株(6b),其发酵生产琥珀酸时,发酵液中副产物乙酸含量显著降低,琥珀酸含量有所提高;
[0164]
在埃希氏菌hb1基因组中敲除磷酸乙酰转移酶基因pta获得的hb74菌株(6c),其发酵生产琥珀酸时,发酵液中不含副产物乙酸,琥珀酸含量进一步提高。
[0165]
如图7所示,在埃希氏菌hb1基础上,进行ldha基因、pflb基因、adhe基因和acka基因的敲除改造,所得埃希氏菌hb332在厌氧发酵4天内积累了157.1mm的乙酸,主要产物琥珀酸含量达到711.4mm,经检测,发酵液中不含甲酸、乳酸和乙醇等副产物。
[0166]
在埃希氏菌hb1基础上,进行ldha基因、pflb基因、adhe基因和pta基因的敲除改造,所得埃希氏菌hb333在厌氧发酵4天内仅产生20.9mm的乙酸,而琥珀酸产量达到723.4mm,发酵液中不含甲酸、乳酸和乙醇等副产物;相较于埃希氏菌hb1,琥珀酸含量显著提高,且副产物积累量大大降低。
[0167]
以上仅是本发明的优选实施方式,本发明的保护范围并不仅局限于上述实施例,凡属于本发明思路下的技术方案均属于本发明的保护范围。应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理前提下的若干改进和润饰,应视为本发明的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1