CsHO1在调控茶叶中茶氨酸的积累中的应用
csho1在调控茶叶中茶氨酸的积累中的应用
技术领域
1.本发明涉及茶叶中茶氨酸含量调控技术领域,具体涉及csho1在调控茶叶中茶氨酸的积累中的应用。
背景技术:
2.茶树是世界上最重要的经济作物之一。它含有丰富的次级代谢物,这些代谢物赋予茶叶的感官品质和健康益处。其中,茶氨酸(γ-谷氨酰乙酰胺)是茶树中一种独特的非蛋白性氨基酸,是赋予绿茶独特鲜味的成分。此外,茶氨酸具有许多健康益处,包括促进放松和镇静,改善认知功能(学习和记忆),以及改善人类和动物由焦虑引起的睡眠状况。据推测,茶氨酸增加抑制性神经递质的水平,包括γ-氨基丁酸(gaba)、五羟色胺和多巴胺。因此,茶氨素含量在很大程度上决定了绿茶的质量和价格。
3.茶氨酸生物合成以谷氨酸(glu)和乙胺(ea)为前体,由茶氨酸合成酶(cstsi)催化。管辖酰胺合成酶csgs1.1和csgs2也有助于幼嫩新梢中的茶氨酸合成。谷氨酸合酶(gogats)和谷氨酸脱氢酶(gdhs)催化茶树中的glu生物合成,而ea是由丙氨酸脱羧酶(csaladc)从丙氨酸合成的。最近鉴定了编码gogats、gdhs和csladc的候选基因。茶氨酸的分解代谢可能由茶树中的cspdx2.1催化。茶氨酸生物合成和分解代谢过程受到许多环境因素的调节,特别是营养水平、光照强度和盐胁迫等。
4.白化和黄化茶树品种,如“黄山白茶”和“黄魁”等,是对高质量绿茶生产具有重要价值的茶树种质资源,生产的白化或黄化茶叶深受茶叶市场的欢迎。这是因为白化或黄化茶叶含有较高茶氨酸和较低茶多酚,茶汤鲜爽味突出,没有明显的苦涩味。然后,白化和黄化的茶叶中为什么会积累高水平的茶氨酸,背后的分子机理在很大程度上仍然是未知的。
5.鉴于上述问题,本发明创作者经过长时间的研究和实践终于获得了本发明。
技术实现要素:
6.本发明的目的在于解决如何将影响白化和黄化的csho1应用于调控茶叶中茶氨酸积累的问题,提供了csho1在调控茶叶中茶氨酸积累水平中的应用。
7.为了实现上述目的,本发明公开了csho1在调控茶叶中茶氨酸积累水平中的应用,通过降低csho1表达量,提高茶叶中茶氨酸含量。所述csho1的碱基序列如seq id no.1所示。
8.所述csho1的表达促进茶氨酸在新梢中降解为谷氨酸和乙胺。
9.所述csho1调控了茶树新梢各器官和不同季节新梢种茶氨酸的积累。
10.所述茶叶为茶树新梢。
11.与现有技术比较本发明的有益效果在于:本发明通过发现csho1在黄化和白化茶树品种中的表达量远低于正常绿色品种,从而对csho1的表达量与茶叶中茶氨酸的积累之间的关系进行了研究,将csho1用于对茶叶中茶氨酸的积累进行调控,通过降低csho1的表达量提高茶叶中的茶氨酸积累量。
附图说明
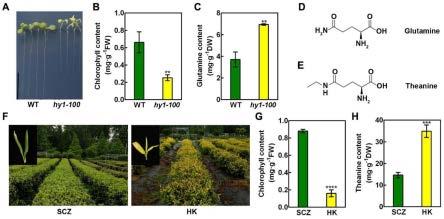
12.图1为黄化的拟南芥和茶树幼苗中谷氨酰胺、茶氨酸、叶绿素含量;(a)拟南芥野生型(wt)和hy1-100突变体的表型;(b)叶绿素含量;(c)谷氨酰胺含量;(d)谷氨酰胺的化学式;(e)茶氨酸的化学式;(f)正常绿色茶树品种“舒茶早”(scz)和黄化品种“黄魁”(hk)的叶片颜色;(g)叶绿素含量;(h)茶氨酸含量;
13.图2为系统发育树、多序列比对以及csho1的亚细胞定位;(a)山茶(cs)、拟南芥(at)、甘蓝(bo)、烟草(nt)、番茄茄(sl)、葡萄(vv)、tremula(pt)、可可(tc)、大豆(gm)、水稻(os)、玉米(zm)中hos的系统发育关系;(b)csho1与其他植物ho1蛋白的比对;(c)拟南芥原生质体中csho1蛋白的亚细胞定位;
14.图3为正常、黄化和白化茶树品种中csho1的表达和茶氨酸含量;
15.图4为不同组织、不同时间点csho1表达与茶氨酸含量的关系;
16.图5为hy1-100和csho1转基因hy1-100突变体中的茶氨酸积累;(a)野生型(wt)、hy1-100和csho1转基因hy1-100突变体(com1和com2)在ms培养基中添加或不添加0、5、10或15mm茶氨酸14天的生长表型;(b)拟南芥品系的茶氨酸含量;(c)拟南芥品系的根长;
17.图6为抑制茶树新捎中csho1表达对茶叶中茶氨酸积累的影响;(a)新梢经40μm sdon或asodn处理24h后,实时pcr分析csho1的表达;(b)新梢中的茶氨酸含量;
具体实施方式
18.以下结合附图,对本发明上述的和另外的技术特征和优点作更详细的说明。
19.本实施例使用的8年树龄茶树品种。这些品种包括“舒茶早”(scz)、“浙农113”(zn113)、“中茶302”(zc302)、“黄魁”(hk)和“黄山白茶”(hsbc),在郭河茶园(31
°
n,117
°
e,中国)种植。组织包括根、茎、叶芽和新梢的第1、2、3、4、5片叶。
20.拟南芥hy1-100突变体来自中国南京农业大学生命科学学院沈文彪和谢延杰教授。按照常规对种子进行杀菌,并在固态(ms)培养基上培养。种子在4℃下分层培养2d后转入生长室。条件设置为昼夜光照期16h/8h,光照强度150μmol m-2
s-1
辐照度,23℃/18℃,70%的相对湿度。拟南芥野生型(wt)、hy1-100突变系和转基因系在添加或不添加0、5、10或15mm茶氨酸的ms培养基上生长14d。
21.一、拟南芥黄化突变体hy1-100和茶树品种“黄魁”中gln或茶氨酸的超积累
22.haem oxygenase 1(ho1/hy1)在拟南芥叶绿素合成中起着至关重要的作用,用95%(v/v)乙醇在黑暗中提取幼苗叶绿素24h,然后通过检测649nm和665nm处的吸光度计测定叶绿素含量,通过验证拟南芥ho1突变体hy1-100表现出白化表型,叶绿素含量低得多(图1a,b)。hy1-100比野生型(wt)幼苗积累了更多的gln,即茶氨酸的类似物(图1c-e)。同样,黄化茶树品种“黄魁”(hk)比正常的绿茶品种“舒茶早”(scz)积累了更低水平的叶绿素和更高水平的茶氨酸(图1f-h)。这些结果表明,ho1调节拟南芥中gln的积累。
23.二、csho1在序列和亚细胞定位
24.使用csho1特异性引物从茶树品种”舒茶早”的cdna文库中扩增该基因。将无终止密码子的pcr片段插入pan580载体中,与绿色荧光蛋白(gfp)融合。将获得的pan580-csho1-gfp和空载体pan580-gfp通过聚乙二醇(peg)介导的基因转化转化为拟南芥原生质体。室温下在黑暗中孵育16小时后,使用激光扫描共聚焦显微镜观察荧光。
25.通过茶树信息档案在茶树中搜索了ho同源体,并确定了3个可能的cshos。系统发育树、多序列比对和csho1的亚细胞定位如图2所示,由图可知,cshos属于两个亚科:ho1亚科和ho2亚科。csho1和csho3明显归为ho1亚家族,其中烟草、番茄茄、拟南芥、甘蓝、葡萄、胡杨、可可、甘氨酸、水稻和玉米中均有ho1、ho3和ho4(图2a)。csho1编码了289个氨基酸残基,其中58个氨基酸转运肽被chlorop算法识别。在各种植物物种中鉴定出了几个高度保守的ho1s结构域(图2b),说明在其他植物中,csho1保守于ho1s。
26.为了评估csho1的亚细胞定位,我们将csho1与camv 35s启动子(35s::csho1-gfp)驱动的绿色荧光蛋白(gfp)融合。35s::csho1-gfp和对照35s::gfp被转化为拟南芥原生质体。如图2c所示,细胞质和细胞核中都可见到游离的gfp荧光。相比之下,csho1-gfp的荧光信号与叶绿体中叶绿素的自荧光信号重叠。这些结果表明,csho1定位于叶绿体中。这种定位与紫花苜蓿、番茄、小麦和黄瓜中的ho1一致。
27.三、黄化和白化的茶树品种叶片中csho1的表达量
28.比较了csho1在三个正常茶树品种(scz,zn113,zc302)、黄化品种hk和白化品种hsbc新枝中的表达。结果表明,hk和hsbc的表达水平远低于正常绿色品种(图3)。在本研究和以往的研究中,hk和hsbc的叶绿素含量均明显低于绿色品种(图1g)。因此,这个结果表明了csho1在茶树中的叶绿素生物合成中具有积极的作用,就像它在其他植物中的同源物一样。
29.对这些品种新芽中的茶氨酸含量进行测定,如图3所示,茶氨酸含量在这些品种中表现出与csho1表达相反的模式,在hk和hsbc中茶氨酸含量较高,进一步分析csho1表达与茶氨酸含量的相关系数。结果表明,csho1表达量与茶氨酸含量呈高度负相关(r=-0.98,p《0.001)。这一结果支持了csho1负向调控茶树茶氨酸积累的观点。
30.四、csho1表达与茶氨酸含量关系
31.检测了茶树品种中csho1在不同器官中的表达和茶氨酸积累水平。被检测的组织包括根、嫩茎、叶芽、1
st
叶、2
nd
叶、3
rd
叶、4
th
叶、5
th
叶(图4)。在这些器官中,csho1的表达逐渐增加;相反,茶氨酸含量逐渐降低(图4b)。进一步的相关分析表明,csho1表达与这些组织中茶氨酸的积累呈显著负相关。
32.茶氨酸在茶树嫩枝中的积累具有高度的季节依赖性,因此,分别在冬季12月12日、春季(3月1日、3月23日、arp 13日)和夏季(6月14日)5个时间点检测了茶树叶芽中csho1的表达和茶氨酸含量。再次,csho1表达与茶氨酸含量呈现相反的趋势,相关系数为-0.97,p《0.01(图4c)。这些结果进一步表明,csho1对茶树中茶氨酸的积累具有负调控作用。
33.csho1在不同器官中的表达水平是不同的,也对季节有响应(图4),更重要的是,csho1的表达水平与茶氨酸含量呈高度负相关。这些结果表明,csho1可能调控了芽器官和不同季节茶氨酸的积累。考虑到茶氨酸主要在根中合成,并在新梢中分解,csho1可能促进茶氨酸在新梢中降解为谷氨酸和乙胺。
34.五、atho1和csho1对茶氨酸饲喂拟南芥的茶氨酸的积累的影响
35.用基因特异性引物rt-pcr扩增csho1编码区,克隆到含有camv35s启动子的pcambia1302载体中。将重组质粒转化为农杆菌根癌株gv3101,然后采用花浸法转化拟南芥hy1-100突变体。
36.测试了atho1是否调节了茶氨酸饲喂拟南芥的茶氨酸积累。将不同浓度的茶氨酸
(0、5、10和15mm)添加到ms培养基中,喂养拟南芥野生型(wt)和hy1-100突变体。从茶氨酸对根生长的抑制作用来看,hy1-100突变体对高浓度的茶氨酸(10和15mm)表现出高度敏感(图5a,c),在相同条件下,hy1-100突变体的茶氨酸积累量也明显高于wt(图5b)。这些结果表明,atho1调节了饲喂茶氨酸的拟南芥中茶氨酸的积累,也提示拟南芥可以作为研究茶氨酸积累调控的模式植物。
37.为了验证csho1在茶氨酸积累中的作用,将35s启动子驱动的csho1转入拟南芥hy1-100突变体中。csho1在hy1-100突变体中的表达恢复了两个独立转基因系(com1和com2)中hy1-100的黄化表型(图5a)。这一结果表明,在调节叶绿素生物合成方面,csho1与atho1在功能上是保守的。此外,在10和15mm饲养条件下,hy1-100突变体中csho1的表达也恢复了茶氨酸超积累表型(图5b)。这一结果为csho1在茶氨酸积累中的负调控作用提供了遗传学证据。
38.六、茶树新芽中csho1表达对茶氨酸的影响
39.利用soligo软件设计候选反义寡核苷酸(asodn),以csho1 cdna序列为输入。以相应的正义寡核苷酸(sodn)作为对照。新梢有一个芽和1
st
叶片用0.1ml 40μm asodn或sodn溶液处理。孵育24h后,采收新梢,-80℃保存。
40.为了csho1在茶树茶氨酸积累中的负面作用提供体内遗传学证据,用csho1特异性的正义寡核苷酸(sodn)或反义寡核苷酸(asodn)处理scz的新梢24h(图6a)。与对照sodn处理相比,asodn处理显著抑制了csho1的表达水平(图6b)。同时,在asodn处理的新梢中茶氨酸积累显著增加(图6b)。这些结果表明,csho1在基因上调控了茶树中茶氨酸的积累。
41.综上所述,在本发明中,通过发现csho1在黄化和白化茶树品种中的表达量远低于正常绿色品种(图3),更重要的是,当csho1在hy1-100突变体中表达时,该突变体的黄化表型和叶绿素含量得到了恢复(图5),以突变体中饲喂茶氨酸,突变体茶氨酸积累量较高的表型也得到了恢复。更重要的是,利用asodn在茶树幼苗中瞬时降低csho1的表达显著增加了茶氨酸的积累(图6),这些遗传分析为支持csho1在茶树中调节茶氨酸积累的作用提供了强有力的证据。因此,在这些黄化或白化的茶树中,csho1的低表达可能是导致叶绿素含量低和茶氨酸含量高的原因。
42.在hy1-100突变体反馈中血红素积累会抑制谷氨酸的叶绿素生物合成。因此,叶绿体生物合成的减少很可能导致黄化和白化植物谷氨酸积累增多在茶树中,谷氨酸既是茶氨酸生物合成的底物,又是茶氨酸降解的产物。黄化和白化茶树品种中谷氨酸的过积累可能会反馈抑制茶氨酸降解或促进茶氨酸生物合成,从而增加茶氨酸在这些品种中的积累。本研究揭示了茶树中茶氨酸积累的一种新的调控机制,也为创造茶业急需的黄化或白化茶树种质提供了靶基因。
43.以上所述仅为本发明的较佳实施例,对本发明而言仅仅是说明性的,而非限制性的。本专业技术人员理解,在本发明权利要求所限定的精神和范围内可对其进行许多改变,修改,甚至等效,但都将落入本发明的保护范围内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1