糖代谢相关蛋白质IbpPGM及其生物材料和应用
本发明属于生物,尤其涉及糖代谢相关蛋白质ibppgm及其生物材料和应用。
背景技术:
1、淀粉是植物中碳水化合物的主要储藏形式。在白天,叶片通过光合作用在叶绿体中合成过渡型淀粉(transitory starch),其在夜间又被降解成蔗糖并运往非光合器官作为能量来源。叶绿体中过渡型淀粉的合成底物是adp-葡萄糖(adp-glucose,adpg),其主要由质体型葡萄糖磷酸异构酶(ppgi)、葡萄糖磷酸变位酶(phosphoglucomutase,pgm)以及adp-葡萄糖焦磷酸化酶(adpase)催化合成,之后adpg在淀粉合成酶等的催化下生成直链淀粉以及支链淀粉。
2、pgm是葡萄糖-6-磷酸(g6p)与葡萄糖-1-磷酸(g1p)相互转变的催化酶,而g1p则是adpg的前体物质。植物pgm分为质体型(ppgm)和胞质型(cpgm),二者在光合碳产物的代谢分配中起着相互协同的作用。在植物细胞中,pgm通过参与3个催化步骤来维持淀粉和蔗糖之间的平衡:1、ppgm将自养器官(叶绿体)或异养器官(造粉体)中的g6p催化转变为g1p;2、ppgm将淀粉磷酸化酶磷酸解淀粉形成的g1p催化转变为g6p;3、cpgm通过将细胞质中的g6p催化转变为g1p参与蔗糖合成。马铃薯、菠菜、豌豆中均有1个ppgm成员和1个cpgm成员,而在拟南芥、烟草、以及玉米中则有1个ppgm成员和2个cpgm成员。
3、拟南芥和烟草ppgm基因缺失突变体的叶片中几乎检测不到淀粉含量,但是可溶性糖的含量提高。豌豆中编码ppgm基因的位点rug3突变后导致叶片和种子中的淀粉含量均降低,从而证明了g6p是豌豆造粉体中磷酸己糖的来源。对马铃薯中ppgm或cpgm基因进行抑制表达后其块茎淀粉含量降低,同时叶片光合速率下降。拟南芥atppgm突变体同样出现了光合抑制现象,然而过表达拟南芥atppgm基因或atcpgm基因的烟草植株并未出现光合速率的变化。虽然pgm在光合碳产物的代谢分配中有重要作用,但是其如何影响光合速率仍然需要进一步研究。此外,研究发现pgm功能的缺失会导致配子体败育,原因可能是生殖器官中碳能源物质的供应不足。因此,对花粉中淀粉或蔗糖代谢途径进行调控是诱导雄性不育的有效手段,该手段也被广泛应用于育种实践中。
4、近年来,越来越多的证据表明植物中存在淀粉合成分支途径,即作为淀粉合成底物的adpg直接在细胞质中由蔗糖合成酶催化生成并转运至叶绿体中。分别抑制马铃薯ppgm或cpgm基因的表达会导致其块茎淀粉含量的下降,然而当这两个基因被同时抑制表达时转基因马铃薯块茎淀粉含量同野生型相比则无显著差异,这说明淀粉合成的前体物质(如udpg、adpg、g1p)可被转运至异养器官中来代替g6p。在谷类胚乳中,adpg的前提物质g6p可从胞质中转运至造粉体,而adpg也可直接由胞质型apgase合成并由adpg转运蛋白转运至造粉体中进行淀粉合成。因此,除了依靠pgm进行质体内g6p和g1p的催化转换,自养器官和异养器官中均存在跨质体膜的转运途径来向质体内提供淀粉合成的前体物质。
5、甘薯是重要的根茎类作物,其块根富含淀粉,既可食用也可作为生物能源原料。甘薯中淀粉合成途径的关键基因功能已有报道。然而作为淀粉合成途径中有重要功能的pgm基因的功能却尚未明确。
技术实现思路
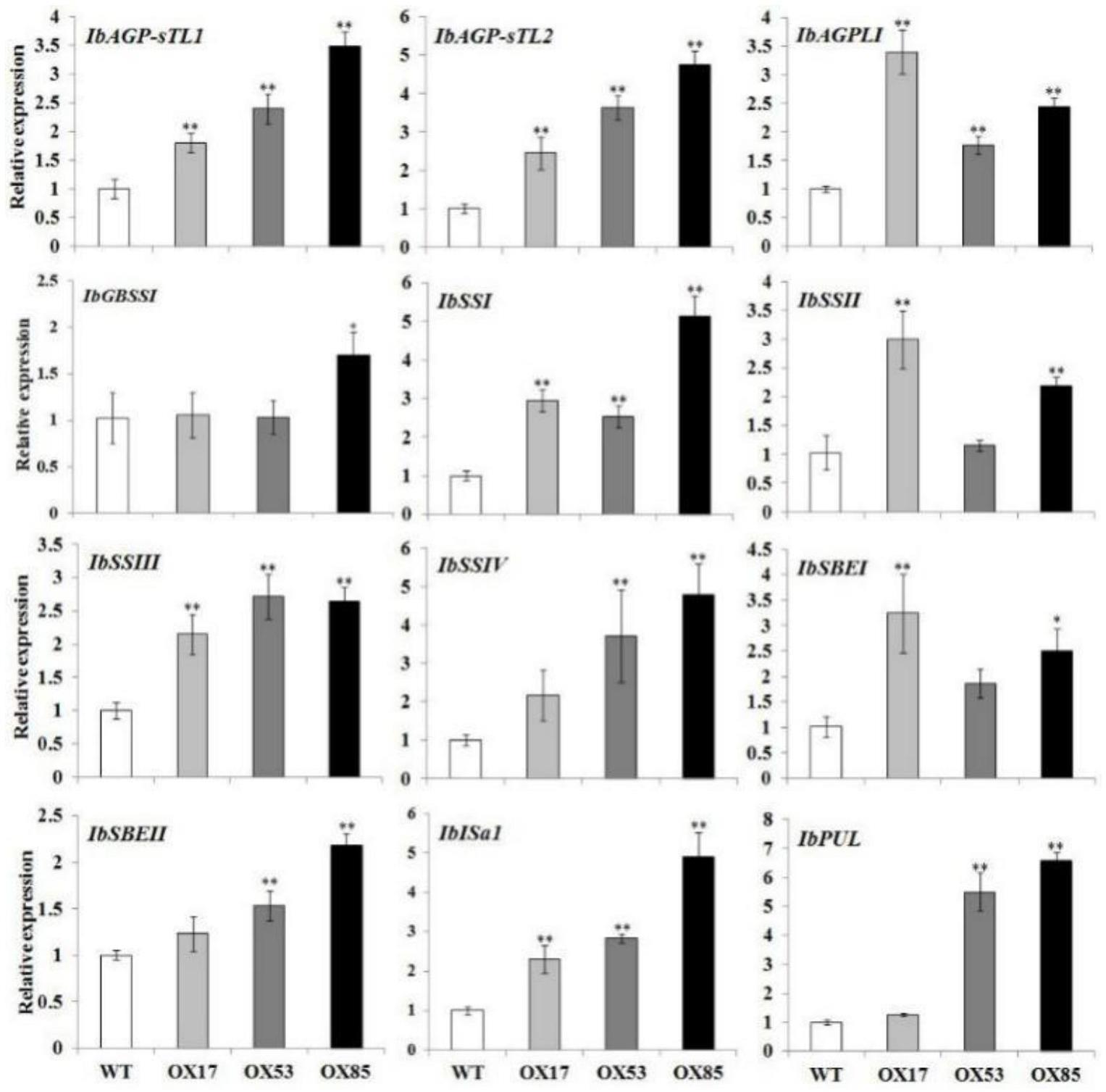
1、本发明所要解决的技术问题是如何调控植物蔗糖含量和/或葡萄糖含量和/或果糖含量和/或淀粉含量以及adp-葡萄糖焦磷酸化酶小亚基基因1(ibagp-stl1)、葡萄糖焦磷酸化酶小亚基基因2(ibagp-stl2)、adp-葡萄糖焦磷酸化酶大亚基基因(ibagp-tli)、颗粒结合型淀粉合成酶(ibgbssi)、可溶性淀粉合成酶基因i(ibssi)、可溶性淀粉合成酶基因ii(ibssii)、可溶性淀粉合成酶基因iii(ibssiii)、可溶性淀粉合成酶基因iv(ibssiv)、淀粉分支酶基因i(ibsbei)、淀粉分支酶基因ii(ibsbeii)、异淀粉酶基因(ibisa1)或普鲁兰酶基因(ibpul)的表达量。
2、为了解决上述问题,本技术提供了蛋白质。
3、所述蛋白质为如下述任一项:
4、a1)氨基酸序列是序列2所示的蛋白质;
5、a2)将a1)所述蛋白质的经过氨基酸残基的取代和/或缺失和/或添加得到的与a1)所示的蛋白质具有80%以上的同一性且具调控植物蔗糖含量和/或葡萄糖含量和/或果糖含量和/或淀粉含量的蛋白质;
6、a3)将a1)或a2)的n末端或/和c末端连接蛋白质标签得到的融合蛋白质。
7、序列2如下所示(seq id no.2):
8、masfcarieptsisiskqfksarnafplsppirclsffnspssnfplrktvpsssspifaassspss
9、ssssppatvaesqalkiksvptkpiegqktgtsglrkkvkvfmqdnylanwiqalfnslape
10、dykdqllvlggdgryfnreaaqliiqiaagngvgqimigkdgimstpavsavirkrkangg
11、fimsashnpggpdydwgikfnyssgqpapesitdkiygntlsiseikmadipdvdlsqlgvtr
12、ygnfsvevvdpvgdylelmqevfdfslirdllsrpnfrfvfdamhavtgayakpifvdmlg
13、aspesivngvpledfghghpdpnltyakdlvnvmfgengpdfgaasdgdgdrnmilgrqf
14、fvtpsdsvaiiaanakeaipyfksgpkglarsmptsgaldrvaeklnllfyevptgwkffgn
15、lmdagklsvcgeesfgtgsdhirekdgiwavlawlsiiayrnkdkkpgealvsvgdvvkq
16、hwatygrnffsrydyeecesegankmvaylrelistskagdkygsyvlkfaddfsyvdpv
17、dgsvaskqgvrfvftdgsriifrlsgtgsagatvrvyieqfesdaskhdvdaqialkpliela
18、lslsklkeftgrekptvit
19、上述蛋白质中,所述蛋白标签(protein-tag)是指利用dna体外重组技术,与目的蛋白一起融合表达的一种多肽或者蛋白,以便于目的蛋白的表达、检测、示踪和/或纯化。所述蛋白标签可为flag标签、his标签、mbp标签、ha标签、myc标签、gst标签和/或sumo标签等。
20、上述蛋白质中,同一性是指氨基酸序列的同一性。可使用国际互联网上的同源性检索站点测定氨基酸序列的同一性,如ncbi主页网站的blast网页。例如,可在高级blast2.1中,通过使用blastp作为程序,将expect值设置为10,将所有filter设置为off,使用blosum62作为matrix,将gap existence cost,per residue gap cost和lambda ratio分别设置为11,1和0.85(缺省值)并进行检索一对氨基酸序列的同一性进行计算,然后即可获得同一性的值(%)。
21、上述蛋白质中,所述80%以上的同一性可为至少81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、95%、96%、98%、99%或100%的同一性。
22、上述蛋白质中,序列2(seq id no.2)由个638个氨基酸残基组成。
23、上述蛋白质中,所述蛋白质来源于甘薯。
24、
技术实现要素:
25、所述蛋白质的名称可为ibppgm,可来源于甘薯。具体可为甘薯块根。所述甘薯可为甘薯高淀粉品系徐781、甘薯高淀粉品种郑红22或郑红23号。
26、为了解决上述问题,本技术还提供了生物材料。
27、所述生物材料为下述任一种:
28、b1)、编码上所述蛋白质的核酸分子;
29、b2)、含有b1)所述核酸分子的表达盒;
30、b3)、含有b1)所述核酸分子的重组载体、或含有b2)所述表达盒的重组载体;
31、b4)、含有b1)所述核酸分子的重组微生物、或含有b2)所述表达盒的重组微生物、或含有b3)所述重组载体的重组微生物;
32、b5)、含有b1)所述核酸分子的转基因植物细胞系、或含有b2)所述表达盒的转基因植物细胞系、或含有b3)所述重组载体的转基因植物细胞系;
33、b6)、含有b1)所述核酸分子的转基因植物组织、或含有b2)所述表达盒的转基因植物组织、或含有b3)所述重组载体的转基因植物组织;
34、b7)、含有b1)所述核酸分子的转基因植物器官、或含有b2)所述表达盒的转基因植物器官、或含有b3)所述重组载体的转基因植物器官。
35、上文中,b1)所述的核酸分子为下列任一中:
36、c1)编码链的核苷酸序列为序列1的dna分子;
37、c2)将c1)所述核酸分子的经过核苷酸的取代和/或缺失和/或添加得到的与c1)所示的核酸分子具有80%以上的同一性且具调控植物淀粉含量和/或葡萄糖含量和/或果糖含量和/或蔗糖含量的核酸分子
38、上文中,所述b1)所述的核酸分子为序列1所示的dna分子。
39、序列1如下所示(seq id no.1):
40、atggcgtcgttttgtgcgaggatcgaaccgacctccatttccatctcgaagcagttcaaa
41、tccgccaggaatgcctttcctctctctccaccaatccgatgcctctccttcttcaattccc
42、cttcctctaatttccctctcaggaaaactgtgccttcttcttcgtctcccatcttcgccgc
43、ttcttcatctccttcctcctcctcctcttctcctcccgctaccgtcgccgaatctcaagca
44、ctcaagattaaatcggttccgacaaagccaatcgaaggacagaagacgggaactagtg
45、ggctccgtaagaaggttaaagtttttatgcaagataattaccttgcgaattggattcagg
46、cattgtttaattcgttggcgcctgaggattataaggaccagttgttggttctcggaggtg
47、atggccgatattttaatcgcgaagctgcacagttaatcattcaaattgctgctggcaatg
48、gggttggtcaaattatgattggcaaggatggaataatgtctactccagctgtgtctgctg
49、tgatacgaaagagaaaggctaatggtggctttataatgagtgcaagccataatcctggtg
50、gtccagactatgattggggcatcaagttcaattacagcagtggtcaaccagcaccagaa
51、tctattactgacaaaatatacgggaacacgctttctatttctgaaattaagatggctgaca
52、ttcctgatgttgatctctctcaacttggagttactagatatgggaattttagtgttgaagt
53、ggttgacccagtaggtgactatttggagctaatgcaggaagtgtttgatttttcacttat
54、cagagatcttctttccagaccaaatttcaggtttgtgtttgatgccatgcatgctgtcac
55、tggtgcttatgcaaagcctatttttgttgacatgctaggagctagcccggaatctattgtt
56、aatggcgtgcctcttgaagattttggacatggtcatccagaccctaatcttacatatgcg
57、aaagatttggtcaatgtaatgtttggcgagaatggacctgattttggtgctgcaagtgat
58、ggggatggtgacagaaatatgattctaggtaggcaattttttgttactccgtcagattct
59、gtagcaattattgctgccaatgcaaaagaggccattccatacttcaaaagtggtcccaa
60、gggattggctcgctctatgcccactagtggtgctttggaccgtgttgcggaaaagctaa
61、atcttctgttttacgaggttcctactggatggaaattctttgggaatctaatggatgcag
62、gaaagttgtcagtttgtggggaagaaagttttgggacaggttctgaccacattcgtga
63、gaaagatggtatatgggctgtattagcttggctttcaataattgcatataggaacaagga
64、caagaaaccaggggaggcattggtttctgttggtgatgttgtcaagcagcattgggcaa
65、cttatgggaggaatttcttttctagatatgactatgaggaatgtgaatctgaaggagcca
66、ataagatggttgcatatcttagagaactaatctctaccagtaaggctggtgataagtatg
67、gaagttatgtcctcaaatttgccgatgacttctcctatgttgatccagtagatggaagtg
68、ttgcatccaaacagggggtccgatttgtgttcactgatggatcaaggatcatctttagat
69、tatcgggtactggttctgctggtgcaacagtaagagtgtatattgaacagtttgagtctg
70、atgcctctaagcatgatgtggatgcccaaattgcattgaaaccattgatagaactcgctc
71、tgtctttatcaaagctaaaggaatttaccggaagagagaagccaactgtcataacataab1)所述核酸分子中,本领域普通技术人员可以很容易地采用已知的方法,例如定向进化或点突变的方法,对本发明的编码蛋白质ibppgm的核苷酸序列进行突变。那些经过人工修饰的,具有与本发明分离得到的蛋白质ibppgm的核苷酸序列80%或80%以上同一性的核苷酸,只要编码蛋白质ibppgm且具有蛋白质ibppgm功能,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。
72、上述80%或80%以上同一性,可为81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性。
73、本文中,同一性是指氨基酸序列或核苷酸序列的同一性。可使用国际互联网上的同源性检索站点测定氨基酸序列的同一性,如ncbi主页网站的blast网页。例如,可在高级blast2.1中,通过使用blastp作为程序,将expect值设置为10,将所有filter设置为off,使用blosum62作为matrix,将gap existence cost,per residue gap cost和lambda ratio分别设置为11,1和0.85(缺省值)并进行检索以对氨基酸序列的同一性进行计算,然后即可获得同一性的值(%)。
74、上述生物材料中,b1)所述核酸分子可为所述蛋白质的编码基因。b1)所述核酸分子具体可为编码链的编码序列是序列1所示的dna分子。
75、本文中,所述载体是本领域技术人员公知的,包括但不限于:质粒、噬菌体(如λ噬菌体或m13丝状噬菌体等)、黏粒(即柯斯质粒)、ti质粒或病毒载体。具体可为载体pet-28a和/或pmdc83和/或pbi121;
76、上述生物材料中,b2)所述的表达盒是指能够在宿主细胞中表达所述基因的dna,该dna不但可包括启动基因转录的启动子,还可包括终止基因转录的终止子。进一步,所述表达盒还可包括增强子序列。可用于本发明的启动子包括但不限于:组成型启动子,组织、器官和发育特异的启动子,和诱导型启动子。启动子的例子包括但不限于:花椰菜花叶病毒的组成型启动子35s;来自西红柿的创伤诱导型启动子,亮氨酸氨基肽酶("lap",chao等人(1999)plant physiol 120:979-992);来自烟草的化学诱导型启动子,发病机理相关1(pr1)(由水杨酸和bth(苯并噻二唑-7-硫代羟酸s-甲酯)诱导);西红柿蛋白酶抑制剂ii启动子(pin2)或lap启动子(均可用茉莉酮酸曱酯诱导);热休克启动子(美国专利5,187,267);四环素诱导型启动子(美国专利5,057,422);种子特异性启动子,如谷子种子特异性启动子pf128(cn101063139b(中国专利200710099169.7)),种子贮存蛋白质特异的启动子(例如,菜豆球蛋白、napin,oleosin和大豆beta conglycin的启动子(beachy等人(1985)embo j.4:3047-3053))。它们可单独使用或与其它的植物启动子结合使用。此处引用的所有参考文献均全文引用。合适的转录终止子包括但不限于:农杆菌胭脂碱合成酶终止子(nos终止子)、花椰菜花叶病毒camv 35s终止子、tml终止子、豌豆rbcs e9终止子和胭脂氨酸和章鱼氨酸合酶终止子(参见,例如:odell等人(1985)nature,313:810;rosenberg等人(1987)gene,56:125;guerineau等人(1991)mol.gen.genet,262:141;proudfoot(1991)cell,64:671;
77、sanfacon等人genes dev.,5:141;mogen等人(1990)plant cell,2:1261;munroe等人(1990)gene,91:151;ballad等人(1989)nucleicacids res.17:7891;joshi等人(1987)nucleicacid res.,15:9627)。
78、上述b3)中,可用植物表达载体构建含有所述基因表达盒的重组表达载体。所述植物表达载体可为gateway系统载体或双元农杆菌载体等,如pgwb411、pgwb412、pgwb405、pbin438、pcambia1302、pcambia2301、pcambia1301、pcambia1300、pbi121、pcambia1391-xa或pcambia1391-xb。使用ibppgm构建重组表达载体时,在其转录起始核苷酸前可加上任何一种增强型、组成型、组织特异型或诱导型启动子,如花椰菜花叶病毒(camv)35s启动子、泛素基因ubiqutin启动子(pubi)等,它们可单独使用或与其它的植物启动子结合使用;此外,使用本发明的基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。
79、作为一个具体实施例,本技术使用的原核表达载体可为pet-28a。双元载体可为pmdc83或pbi121。
80、为了解决上述问题,本技术提供了下述用途:
81、材料的下述任一种用途:
82、u1、所述材料在调控植物淀粉含量和/或葡萄糖含量和/或果糖含量和/或蔗糖含量
83、u2、所述材料在制备调控植物淀粉含量和/或葡萄糖含量和/或果糖含量和/或蔗糖含量的产品;
84、u3、所述材料在植物育种中的用途;
85、u4、所述材料在调控植物下述至少一种基因的表达:adp-葡萄糖焦磷酸化酶小亚基基因1、葡萄糖焦磷酸化酶小亚基基因2、adp-葡萄糖焦磷酸化酶大亚基基因、颗粒结合型淀粉合成酶、可溶性淀粉合成酶基因i、可溶性淀粉合成酶基因ii、可溶性淀粉合成酶基因iii、可溶性淀粉合成酶基因iv、淀粉分支酶基因i、淀粉分支酶基因ii、异淀粉酶基因和普鲁兰酶基因;
86、所述材料为下述任一种:
87、c1)上述的蛋白质;
88、c2)上述的生物材料;
89、c3)调控所述蛋白质的编码基因表达的物质;
90、c4)调控所述蛋白质的活性和/或含量的物质。
91、本文中,adp-葡萄糖焦磷酸化酶小亚基基因1为ibagp-stl1基因、葡萄糖焦磷酸化酶小亚基基因2为ibagp-stl2基因、adp-葡萄糖焦磷酸化酶大亚基基因为ibagp-tli基因、颗粒结合型淀粉合成酶为ibgbssi基因、可溶性淀粉合成酶基因i为ibssi基因、可溶性淀粉合成酶基因ii为ibssii基因、可溶性淀粉合成酶基因iii为ibssiii基因、可溶性淀粉合成酶基因iv为ibssiv基因、淀粉分支酶基因i为ibsbei基因、淀粉分支酶基因ii为ibsbeii基因、异淀粉酶基因为ibisa1基因和普鲁兰酶基因为ibpul基因。ibagp-stl1基因cds序列为genbank:z79635(feb-4-2011);ibagp-stl2基因cds序列为genbank:z79636(feb-4-2011);ibagp-tli基因cds序列为genbank:aj252316(jul-26-2016);ibgbssi基因cds序列为genbank:ab071604(aug-9-2006);ibssii基因cds序列为genbank:af068834(mar-10-2010);ibsbei基因cds序列为genbank:ab194725(aug-22-2006);ibsbeii基因cds序列为genbank:ab071286(aug-9-2006);ibisa1基因cds序列为gen bank:dq074643(jan-27-2009))。
92、上述基因可来源于甘薯。
93、本文中,所述调控植物淀粉含量和/或葡萄糖含量和/或果糖含量和/或蔗糖含量可为下调或抑制或降低植物蔗糖含量,和/或,上调或增强或提高植物淀粉含量和/或葡萄糖含量和/或果糖含量。
94、本文中,所述调控植物ibagp-stl1基因和/或ibagp-stl2基因和/或ibagp-tli基因和/或ibgbssi基因和/或ibssi基因和/或ibssii基因和/或ibssiii基因和/或ibssiv基因和/或ibsbei基因和/或ibsbeii基因和/或ibisa1基因和/或ibpul基因的表达可为上调或增强或提高ibagp-stl1基因和/或ibagp-stl2基因和/或ibagp-tli基因和/或ibgbssi基因和/或ibssi基因和/或ibssii基因和/或ibssiii基因和/或ibssiv基因和/或ibsbei基因和/或ibsbeii基因和/或ibisa1基因和/或ibpul基因的表达。
95、本文中,调控所述蛋白质活性和/或含量的物质可为调控基因表达的物质,所述基因编码的蛋白质为下述任一种:
96、a1)氨基酸序列是序列2所示的蛋白质;
97、a2)将a1)所述蛋白质的经过氨基酸残基的取代和/或缺失和/或添加得到的与a1)所示的蛋白质具有80%以上的同一性且具调控植物淀粉含量、葡萄糖含量、果糖含量和蔗糖含量的蛋白质;
98、a3)将a1)或a2)的n末端或/和c末端连接蛋白质标签得到的融合蛋白质。
99、上文中,所述调控基因表达的物质可为进行如下6种调控中至少一种调控的物质:1)在所述基因转录水平上进行的调控;2)在所述基因转录后进行的调控(也就是对所述基因的初级转录物的剪接或加工进行的调控);3)对所述基因的rna转运进行的调控(也就是对所述基因的mrna由细胞核向细胞质转运进行的调控);4)对所述基因的翻译进行的调控;5)对所述基因的mrna降解进行的调控;6)对所述基因的翻译后的调控(也就是对所述基因翻译的蛋白质的活性进行调控)。
100、上述的用途中,其所述植物育种包括培育或选育具有更高淀粉含量和/或葡萄糖含量和/或果糖含量和/或具有更低蔗糖含量的植物。
101、上文中,所述具有高淀粉含量和/或葡萄糖含量和/或果糖含量和/或具有更低蔗糖含量为植物的淀粉含量和/或葡萄糖含量和/或果糖含量高于所述出发植物,蔗糖含量低于所述出发植物。
102、所述出发植物可为通过基因工程手段进行育种的受体植物或通过有性繁殖手段进行育种的供体植物。
103、所述植物可为植物根部。
104、为了解决上述问题,本技术还公开了一种调控植物糖含量的方法。
105、所述调控植物糖含量的方法为a和/或b,所述a为下调或抑制或降低(植物蔗糖含量,所述b为上调或增强或提高植物淀粉含量和/或葡萄糖含量和/或果糖含量;所述方法包括上调或增强或提高目的植物中上述蛋白质的编码基因的表达或上述蛋白质的活性和/或含量来调控植物糖含量。
106、为了解决上述问题,本技术还公开了一种调控植物糖含量的方法。
107、所述培育糖含量改变的植物的方法包括上调或增强或提高目的植物中上所述蛋白质的编码基因的表达或上述蛋白质的活性和/或含量,得到糖含量改变的植物,所述糖含量改变的植物与目的植物相比,具有下述至少一种特性:蔗糖含量降低、淀粉含量升高、葡萄糖含量升高和果糖含量升高。
108、上文中,所述上调或增强或提高目的植物所述蛋白质的编码基因的表达量可通过转基因技术过表达上述蛋白编码基因来实现。
109、上文中,所述过表达上述蛋白编码基因载体可为植物表达载体。
110、所述植物表达载体可为上述b3)所述载体。作为一个具体实施例,本技术的植物表达载体可为pcambia3301载体。
111、上述的方法中,所述蛋白质的编码基因为下述任一种:
112、c1)编码链的核苷酸序列为序列1的dna分子;
113、c2)将c1)所述核酸分子的经过核苷酸的取代和/或缺失和/或添加得到的与c1)所示的核酸分子具有80%以上的同一性且具调控植物淀粉含量和/或葡萄糖含量和/或果糖含量和/或蔗糖含量的核酸分子。
114、上文中,所述蛋白质的编码基因可为序列1的dna分子。
115、为了解决上述问题,本技术还公开了一种上调或增强或提高甘薯中基因表达的方法。
116、所述上调或增强或提高甘薯中基因表达的方法包括上调或增强或提高目的植物中上述蛋白质的编码基因的表达或上述蛋白质的活性和/或含量,得到下述至少一种基因表达被上调或增强或提高的甘薯:基因为下述至少一种基因:adp-葡萄糖焦磷酸化酶小亚基基因1、葡萄糖焦磷酸化酶小亚基基因2、adp-葡萄糖焦磷酸化酶大亚基基因、颗粒结合型淀粉合成酶、可溶性淀粉合成酶基因i、可溶性淀粉合成酶基因ii、可溶性淀粉合成酶基因iii、可溶性淀粉合成酶基因iv、淀粉分支酶基因i、淀粉分支酶基因ii、异淀粉酶基因和普鲁兰酶基因。
117、上述的用途、上述的方法中,所述植物为如下任一种:
118、c1)双子叶植物或单子叶植物;c2)管状花目植物,c3)旋花科植物,c4)甘薯属植物,c5)甘薯。
119、所述甘薯可为甘薯高淀粉品系徐781、甘薯高淀粉品种郑红22或郑红23号。
120、有益效果:
121、本研究从甘薯中克隆到质体型pgm基因ibppgm并对其进行了表达分析以及初步功能鉴定,为通过基因工程手段提高甘薯块根淀粉含量提供了新的候选基因。本技术通过生物工程技术获得ibppgm基因过表达的转基因甘薯。实验证明ibppgm基因的过表达的转基因植株的淀粉含量和/或葡萄糖含量和/或果糖含量高于野生型和/或蔗糖含量低于野生型。本发明克隆并鉴定了一个糖代谢相关蛋白质ibppgm,将其过表达后显著提高甘薯淀粉含量和/或葡萄糖含量和/或果糖含量和/或显著降低蔗糖含量。为甘薯高淀粉含量和/或高葡萄糖含量和/或高果糖含量和/或低蔗糖含量育种提供了新策略、新思路。
- 还没有人留言评论。精彩留言会获得点赞!