本发明属于生物工程领域,具体涉及一种合成三萜化合物和/或甾醇类化合物的重组酵母及其构建方法。
背景技术:
1、鲨烯是一种开链三萜化合物,也是合成许多具有生物活性成分化合物的重要中间体,由鲨烯可以合成β-香树脂醇、麦角甾醇、羊毛甾醇等衍生化合物,因此鲨烯在三萜以及甾醇类化合物合成中起到很关键的作用。
2、β-香树脂醇是一种五环三萜化合物,具有抗炎、抗细菌和真菌、抗病毒及癌细胞防治功效,在降血糖、降血脂方面也具有良好效果。
3、酿酒酵母(saccharomyces cerevisiae)作为真核模式微生物,具有遗传背景清晰、基因操作成熟、完整的膜结构等多方面优势,被认为是萜类合成的适宜底盘宿主。萜类化合物在酿酒酵母中的异源合成,一方面会对菌株产生代谢负担,另一方面合成的终产物或中间产物的胞内积累可能会对细胞产生毒害作用,限制细胞合成萜类化合物的能力。
4、在酿酒酵母三萜化合物合成过程中鲨烯的氧化和2,3-氧化鲨烯的环化过程是限制产物合成的关键因素,其催化效率的高低直接影响了下游化合物的产量,引起鲨烯的积累。此外,鲨烯还处在法尼基焦磷酸(fpp)代谢形成三萜以及直接参与2,3-氧化鲨烯合成途径的关键节点,其直接前体物fpp对细胞有较大的毒害作用。
技术实现思路
1、为解决现有技术的不足,本技术将上清液蛋白因子(supernatant proteinfactor,spf)导入酵母菌中,转运鲨烯和2,3-氧化鲨烯到膜附近被进一步氧化,不仅可以减少鲨烯和2,3-氧化鲨烯的累积,还可以加快胞内代谢速率,提高目标化合物的产量。具体的,在酵母中引入外源spf,验证其对酵母中三萜化合物和/或甾醇类化合物合成的影响。通过诱导型表达的方式在酵母中对外源spf进行功能鉴定;研究spf对酵母细胞生长代谢的影响,并将其应用于三萜化合物(特别是鲨烯或β-香树脂醇)和/或甾醇类化合物(特别是麦角甾醇和/或羊毛甾醇)的生产中。通过研究spf与酵母细胞的适配性,确定蛋白的最佳表达模式;通过调节转运蛋白与内膜系统(例如磷脂合成转录因子ino2)、甲羟戊酸途径关键酶(例如erg1),以及,产物合酶(例如β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶)的关系,构建高效的鲨烯转运体系,强化产物(特别是β-香树脂醇、麦角甾醇和/或羊毛甾醇)的合成,提高产物的产量。为同样需要转运系统的萜类化合物的合成提供有价值的参考资料。
2、为实现上述目的,本发明采用以下技术方案:
3、本发明的第一方面,提供了一种合成三萜化合物和/或甾醇类化合物的重组酵母。
4、优选的,所述的重组酵母表达上清液蛋白因子。
5、进一步优选的,所述的上清液蛋白因子来源于南方多乳鼠、智人和/或原鸡。
6、在本发明的一个具体实施方式中,所述的来源于南方多乳鼠的上清液蛋白因子的编码基因包含seq idno:1所示,或,包含与seq id no:1具有90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或99.9%同一性的核苷酸序列。
7、在本发明的一个具体实施方式中,所述的来源于智人的上清液蛋白因子的编码基因包含seq id no:2所示,或,包含与seq id no:2具有90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或99.9%同一性的核苷酸序列。
8、在本发明的一个具体实施方式中,所述的来源于原鸡的上清液蛋白因子的编码基因包含seq id no:3所示,或,包含与seq id no:3具有90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或99.9%同一性的核苷酸序列。
9、优选的,所述的重组酵母还表达甲羟戊酸途径关键酶、磷脂合成转录因子,和/或,产物合酶。
10、优选的,所述的甲羟戊酸途径关键酶包括但不限于角鲨烯单加氧酶、hmg合酶、fpp合酶或鲨烯合酶中的一种或两种以上的组合。
11、在本发明的一个具体实施方式中,所述的甲羟戊酸途径关键酶包括角鲨烯单加氧酶。
12、优选的,所述的产物合酶包括但不限于α-香树脂醇合酶、β-香树脂醇合酶、达玛烯二醇合酶、羽扇豆醇合酶、环阿屯醇合酶、麦角甾醇合酶或羊毛甾醇合酶中的一种或两种以上的组合。
13、在本发明的一个具体实施方式中,所述的产物合酶为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶。
14、在本发明的一个具体实施方式中,所述的重组酵母表达下列任一组:
15、a)上清液蛋白因子;
16、b)上清液蛋白因子和角鲨烯单加氧酶erg1;
17、c)上清液蛋白因子和磷脂合成转录因子ino2;
18、d)上清液蛋白因子和产物合酶;
19、e)上清液蛋白因子、角鲨烯单加氧酶erg1和磷脂合成转录因子ino2;
20、f)上清液蛋白因子、角鲨烯单加氧酶erg1和产物合酶;
21、g)上清液蛋白因子、磷脂合成转录因子ino2和产物合酶;
22、h)上清液蛋白因子、角鲨烯单加氧酶erg1、磷脂合成转录因子ino2和产物合酶。
23、在本发明的一个具体实施方式中,所述的重组酵母表达来源于南方多乳鼠的上清液蛋白因子、角鲨烯单加氧酶erg1、磷脂合成转录因子ino2和产物合酶(优选为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶)。
24、在本发明的一个具体实施方式中,所述的重组酵母表达来源于智人的上清液蛋白因子、角鲨烯单加氧酶erg1、磷脂合成转录因子ino2和产物合酶(优选为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶)。
25、在本发明的一个具体实施方式中,所述的重组酵母表达来源于原鸡的上清液蛋白因子、角鲨烯单加氧酶erg1、磷脂合成转录因子ino2和产物合酶(优选为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶)。
26、优选的,所述的三萜化合物包括羽扇豆醇、环阿屯醇、达玛烯二醇、α-香树脂醇、鲨烯或β-香树脂醇中的一种或两种以上的组合。
27、在本发明的一个具体实施方式中,所述的三萜化合物为鲨烯或β-香树脂醇。
28、优选的,所述的甾醇类化合物包括羊毛甾醇和/或麦角甾醇。
29、优选的,所述的重组酵母包括但不限于酿酒酵母、毕赤酵母、解脂假丝酵母、解脂耶氏酵母、异常汉逊氏酵母、粟酒裂殖酵母、黏红酵母、热带假丝酵母或产朊假丝酵母中的一种或两种以上的组合。
30、在本发明的一个具体实施方式中,所述的重组酵母为酿酒酵母。
31、优选的,所述的重组酵母为向重组酵母中导入基因获得。所述的导入方式可以为现有技术的任何方式。
32、优选的,所述的导入基因包括上清液蛋白因子的编码基因,还可以包括甲羟戊酸途径关键酶的编码基因、磷脂合成转录因子的编码基因,和/或,产物合酶的编码基因。
33、优选的,所述的导入基因可以为单拷贝或多拷贝。
34、优选的,所述的导入基因在质粒中表达或在染色体上表达。
35、本发明的第二方面,提供了一种构建上述重组酵母的质粒。
36、优选的,所述的质粒包含上清液蛋白因子的编码基因。所述的上清液蛋白因子可以来源于南方多乳鼠、智人和/或原鸡。
37、在本发明的一个具体实施方式中,所述的来源于南方多乳鼠的上清液蛋白因子的编码基因包含seq idno:1所示,或,包含与seq id no:1具有90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或99.9%同一性的核苷酸序列。
38、在本发明的一个具体实施方式中,所述的来源于智人的上清液蛋白因子的编码基因包含seq id no:2所示,或,包含与seq id no:2具有90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或99.9%同一性的核苷酸序列。
39、在本发明的一个具体实施方式中,所述的来源于原鸡的上清液蛋白因子的编码基因包含seq id no:3所示,或,包含与seq id no:3具有90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或99.9%同一性的核苷酸序列。
40、优选的,所述的质粒还包含甲羟戊酸途径关键酶的编码基因、磷脂合成转录因子的编码基因和/或产物合酶的编码基因。
41、优选的,所述的甲羟戊酸途径关键酶包括但不限于角鲨烯单加氧酶、hmg合酶、fpp合酶或鲨烯合酶中的一种或两种以上的组合。
42、在本发明的一个具体实施方式中,所述的甲羟戊酸途径关键酶包括角鲨烯单加氧酶。
43、优选的,所述的产物合酶包括但不限于α-香树脂醇合酶、β-香树脂醇合酶、达玛烯二醇合酶、羽扇豆醇合酶、环阿屯醇合酶、麦角甾醇合酶或羊毛甾醇合酶中的一种或两种以上的组合。
44、在本发明的一个具体实施方式中,所述的产物合酶为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶。
45、在本发明的一个具体实施方式中,所述的质粒包含下列任一组:
46、a)上清液蛋白因子的编码基因;
47、b)上清液蛋白因子的编码基因和角鲨烯单加氧酶erg1的编码基因;
48、c)上清液蛋白因子的编码基因和磷脂合成转录因子ino2的编码基因;
49、d)上清液蛋白因子的编码基因和产物合酶的编码基因;
50、e)上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因和磷脂合成转录因子ino2的编码基因;
51、f)上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因和产物合酶的编码基因;
52、g)上清液蛋白因子的编码基因、磷脂合成转录因子ino2的编码基因和产物合酶的编码基因;
53、h)上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因、磷脂合成转录因子ino2的编码基因和产物合酶的编码基因。
54、在本发明的一个具体实施方式中,所述的质粒包含来源于南方多乳鼠的上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因、磷脂合成转录因子ino2的编码基因和产物合酶(优选为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶)的编码基因。
55、在本发明的一个具体实施方式中,所述的质粒包含来源于智人的上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因、磷脂合成转录因子ino2的编码基因和产物合酶(优选为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶)的编码基因。
56、在本发明的一个具体实施方式中,所述的质粒包含来源于原鸡的上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因、磷脂合成转录因子ino2的编码基因和产物合酶(优选为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶)的编码基因。
57、本发明的第三方面,提供了包含上述质粒的细胞。
58、本发明的第四方面,提供了一种合成三萜化合物和/或甾醇类化合物的重组酵母的构建方法。
59、优选的,所述的构建方法包括向重组酵母中导入上清液蛋白因子的编码基因。所述的上清液蛋白因子可以来源于南方多乳鼠、智人和/或原鸡。
60、在本发明的一个具体实施方式中,所述的来源于南方多乳鼠的上清液蛋白因子的编码基因包含seq idno:1所示,或,包含与seq id no:1具有90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或99.9%同一性的核苷酸序列。
61、在本发明的一个具体实施方式中,所述的来源于智人的上清液蛋白因子的编码基因包含seq id no:2所示,或,包含与seq id no:2具有90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或99.9%同一性的核苷酸序列。
62、在本发明的一个具体实施方式中,所述的来源于原鸡的上清液蛋白因子的编码基因包含seq id no:3所示,或,包含与seq id no:3具有90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或99.9%同一性的核苷酸序列。
63、优选的,所述的构建方法还包括向重组酵母中导入甲羟戊酸途径关键酶的编码基因、磷脂合成转录因子的编码基因,和/或,产物合酶的编码基因。
64、优选的,所述的甲羟戊酸途径关键酶包括但不限于角鲨烯单加氧酶、hmg合酶、fpp合酶或鲨烯合酶中的一种或两种以上的组合。
65、在本发明的一个具体实施方式中,所述的甲羟戊酸途径关键酶包括角鲨烯单加氧酶。
66、优选的,所述的产物合酶包括但不限于α-香树脂醇合酶、β-香树脂醇合酶、达玛烯二醇合酶、羽扇豆醇合酶、环阿屯醇合酶、麦角甾醇合酶或羊毛甾醇合酶中的一种或两种以上的组合。
67、在本发明的一个具体实施方式中,所述的产物合酶为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶。
68、在本发明的一个具体实施方式中,所述的构建方法包括将下列任一组的编码基因导入重组酵母中:
69、a)上清液蛋白因子的编码基因;
70、b)上清液蛋白因子的编码基因和角鲨烯单加氧酶erg1的编码基因;
71、c)上清液蛋白因子的编码基因和磷脂合成转录因子ino2的编码基因;
72、d)上清液蛋白因子的编码基因和产物合酶的编码基因;
73、e)上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因和磷脂合成转录因子ino2的编码基因;
74、f)上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因和产物合酶的编码基因;
75、g)上清液蛋白因子的编码基因、磷脂合成转录因子ino2的编码基因和产物合酶的编码基因;
76、h)上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因、磷脂合成转录因子ino2的编码基因和产物合酶的编码基因。
77、在本发明的一个具体实施方式中,所述的构建方法包括向重组酵母中导入来源于南方多乳鼠的上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因、磷脂合成转录因子ino2的编码基因和产物合酶(优选为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶)的编码基因。
78、在本发明的一个具体实施方式中,所述的构建方法包括向重组酵母中导入来源于智人的上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因、磷脂合成转录因子ino2的编码基因和产物合酶(优选为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶)的编码基因。
79、在本发明的一个具体实施方式中,所述的构建方法包括向重组酵母中导入来源于原鸡的上清液蛋白因子的编码基因、角鲨烯单加氧酶erg1的编码基因、磷脂合成转录因子ino2的编码基因和产物合酶(优选为β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶)的编码基因。
80、优选的,所述的构建方法包括构建包含待导入基因的载体(优选为质粒),然后转化进重组酵母中。
81、优选的,所述的导入的各个基因为单拷贝或多拷贝。
82、优选的,所述的导入的各个基因在载体上表达或在染色体上表达。
83、优选的,所述的三萜化合物包括羽扇豆醇、环阿屯醇、达玛烯二醇、α-香树脂醇、鲨烯或β-香树脂醇中的一种或两种以上的组合。在本发明的一个具体实施方式中,所述的三萜化合物为鲨烯或β-香树脂醇。
84、优选的,所述的甾醇类化合物包括羊毛甾醇和/或麦角甾醇。
85、优选的,所述的重组酵母包括但不限于酿酒酵母、毕赤酵母、解脂假丝酵母、解脂耶氏酵母、异常汉逊氏酵母、粟酒裂殖酵母、黏红酵母、热带假丝酵母或产朊假丝酵母中的一种或两种以上的组合。
86、在本发明的一个具体实施方式中,所述的重组酵母为酿酒酵母。
87、在本发明的一个具体实施方式中,所述的重组酵母为上述第一方面所述的重组酵母。
88、本发明的第五方面,提供了一种合成三萜化合物和/或甾醇类化合物的方法。
89、优选的,所述的方法包括发酵培养上述的重组酵母。
90、优选的,所述的发酵的培养基中包含碳源(例如葡萄糖)、氮源(包括有机氮源或无机氮源)、无机盐、磷源、氨基酸等等现有技术常规的培养基成分,只要可以满足菌种生长和/或产物合成所需的影响成分即可。
91、优选的,所述的发酵可以为两阶段发酵,例如首先增产细胞,然后生产产物。
92、在所述的生产产物的过程中可以采用低温诱导的方式。优选的,所述的低温为20-26℃中任一数值,优选为22-25℃中任一数值,例如20、21、22、23、24、25或26℃。
93、在本发明的一个具体实施方式中,所述的低温为24℃。
94、优选的,所述的三萜化合物包括羽扇豆醇、环阿屯醇、达玛烯二醇、α-香树脂醇、鲨烯或β-香树脂醇中的一种或两种以上的组合。在本发明的一个具体实施方式中,所述的三萜化合物为鲨烯或β-香树脂醇。
95、优选的,所述的甾醇类化合物包括羊毛甾醇和/或麦角甾醇。
96、本发明的第六方面,提供了一种上述重组酵母和/或上述的构建方法获得的重组酵母在合成三萜化合物和/或甾醇类化合物中的应用。
97、本发明在酿酒酵母中对不同来源的spf蛋白进行功能鉴定(例如发酵验证对代谢通路中萜类产物以及甾醇类产物合成的影响)、适配性表达与调控(例如spf蛋白在不同强度组成型和诱导型启动子作用下的调控表达以及在不同底盘宿主中的适配性表达,在此过程中显著改善了细胞的生长状态),在此基础上研究spf蛋白对酵母细胞生长代谢的影响,并将其应用于萜类化合物(特别是β-香树脂醇)和甾醇类化合物(特别是麦角甾醇和/或羊毛甾醇)的生产中。通过调节spf与内膜系统(例如磷脂合成转录因子ino2)、途径关键酶(例如erg1)和产物合酶(例如β-香树脂醇合酶、麦角甾醇合酶和/或羊毛甾醇合酶)之间的关系,构建高效的鲨烯转运体系,强化萜类化合物(特别是β-香树脂醇)和甾醇类化合物(特别是麦角甾醇和/或羊毛甾醇)的合成。最终通过途径调控结合发酵优化提高酵母合成萜类化合物(特别是β-香树脂醇)和甾醇类化合物(特别是麦角甾醇和/或羊毛甾醇)的产量。
98、其中,本发明导入外源spf蛋白,能够通过转运萜类化合物(特别是β-香树脂醇)和甾醇类化合物合成途径重要前体鲨烯并改善菌株的生长状态来增强酿酒酵母合成萜类化合物(特别是β-香树脂醇)和甾醇类化合物(特别是麦角甾醇和/或羊毛甾醇)的能力。
99、本发明所述的“包括”或“包含”是开放式的描述,含有所描述的指定步骤,以及不会实质上影响的其他步骤。当用于描述蛋白质或核酸的序列时,所述蛋白质或核酸可以是由所述序列组成,或者在所述蛋白质或核酸的一端或两端可以具有额外的氨基酸或核苷酸,但仍然具有与原序列相同或相似的活性。
100、本发明所述的“和/或”包含该术语所连接的项目的所有组合,应视为各个组合已经单独地在本文列出。例如,“a和/或b”包含了“a”、“a和b”以及“b”。又例如,“a、b和/或c”包含了“a”、“b”、“c”、“a和b”、“a和c”、“b和c”以及“a和b和c”。
101、本发明简写与全称,以及中英文对照:
102、spf:supernatant protein factor,鲨烯转运蛋白,又称为上清液蛋白因子;
103、nts:nontranscribed spacer,非转录间隔区;
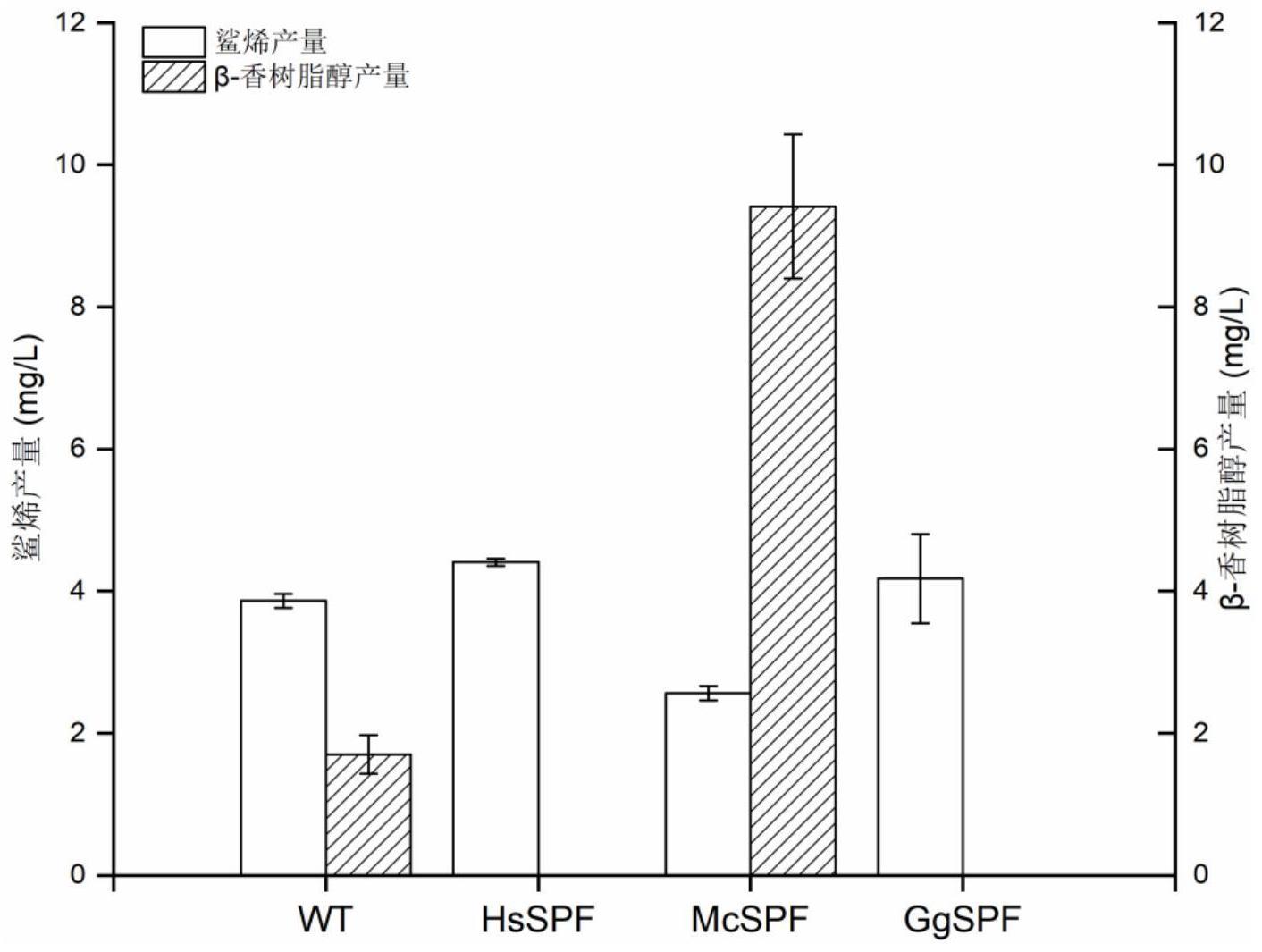
104、rdna:酿酒酵母基因组多拷贝位点;
105、erg1:角鲨烯单加氧酶;
106、ino2:磷脂合成转录因子;
107、ypd:酵母浸出粉胨葡萄糖培养基;
108、bas:β-香树脂醇合酶。
109、本发明的酿酒酵母工程菌具有以下优点:
110、1、本发明中含有mcspf的酵母菌株利用24℃低温诱导,能够实现在特定阶段表达外源spf蛋白,有效减轻异源蛋白表达对酿酒酵母(saccharomyces cerevisiae)自身的代谢负担,本发明所述的导入spf蛋白后的wz48菌株的摇瓶发酵生产β-香树脂醇的产量可达到750.64±54.47mg/l,5l发酵罐发酵生产β-香树脂醇的产量达到2112.48±90.86mg/l,推动了β-香树脂醇的工业化生产。
111、2、本发明在酿酒酵母(saccharomyces cerevisiae)中导入spf蛋白后,加快了从fpp(fpp对细胞有较大的毒害作用)到2,3-氧化鲨烯的次级代谢过程,菌株生长状态得到显著改善,减轻了细胞毒性,有效缩短了培养周期,在一定程度上能够降低菌株的发酵成本。
112、3、本发明中酿酒酵母(saccharomyces cerevisiae)工程菌利用强化中间产物鲨烯传质效率的策略,实现包括鲨烯和β-香树脂醇等三萜化合物的高效合成,同时还对羊毛甾醇、麦角甾醇等甾醇类化合物的合成具有一定的促进作用,可应用于以β-香树脂醇或甾醇类化合物为前体的下游其他代谢产物(如齐墩果酸、甘草次酸、薯蓣皂素等)的高效合成。