抗PD-1抗体的制作方法
抗pd-1抗体
1.本技术是申请日为2017年8月10日、申请号为201780050367.6、发明名称为“抗pd-1抗体”的发明专利申请的分案申请。
技术领域
2.本发明涉及一种抗pd-1抗体,更具体而言,涉及具有包含大鼠抗牛pd-1抗体的互补链决定区(cdr)的可变区和大鼠以外的动物的抗体的恒定区的抗pd-1抗体。
背景技术:
3.免疫抑制受体programmed death 1(pd-1)和作为其配体的programmed death ligand 1(pd-l1)是由京都大学本庶佑等人鉴定为抑制过度的免疫响应、与免疫耐受密切相关的因子的分子(非专利文献1:ishida y,agata y,shibahara k,honjo t the embo j.,11(11):3887-3895;nov.1992.)。近年来已表明还参与肿瘤中的免疫抑制,在人类医疗中,开发出阻碍pd-1的作用的抗体药并得到实用化(小野药品工业株式会社“opdivo(注册商标)”)。
4.此前,本发明人等进行了针对动物难治性疾病的以pd-1或pd-l1为标靶的免疫细胞疗法的开发,阐明该新型免疫细胞疗法可以横跨疾病并且横跨动物地应用(非专利文献2:ikebuchi r,konnai s,shirai t,sunden y,murata s,onuma m,ohashi k.vet.res.,42:103;sep.26,2011.、非专利文献3:ikebuchi r,konnai s,okagawa t,yokoyama k,nakajima c,suzuki y,murata s,ohashi k.vet.res.,44:59;jul.22,2013.、非专利文献4:ikebuchi r,konnai s,okagawa t,yokoyama k,nakajima c,suzuki y,murata s,ohashi k.immunology.142(4):551-61;aug.2014.、非专利文献5:maekawa n,konnai s,ikebuchi r,okagawa t,adachi m,takagi s,kagawa y,nakajima c,suzuki y,murata s,ohashi k.plos one.,9(6):e98415;jun.10,2014.、非专利文献6:mingala cn,konnai s,ikebuchi r,ohashi k.comp.immunol.microbiol.infect.dis.,34(1):55-63;jan.2011.)。
5.但是,由于本发明人等此前所制作的抗体为大鼠抗体,因此无法对大鼠以外的动物反复施用。
6.现有技术文献
7.非专利文献
8.非专利文献1:ishida y,agata y,shibahara k,honjo t.the embo journal.11(11):3887-3895;nov.1992.
9.非专利文献2:ikebuchi r,konnai s,shirai t,sunden y,murata s,onuma m,ohashi k.vet.res.,42:103;sep.2011.
10.非专利文献3:ikebuchi r,konnai s,okagawa t,yokoyama k,nakajima c,suzuki y,murata s,ohashi k.vet.res.,44:59;jul.22,2013.
11.非专利文献4:ikebuchi r,konnai s,okagawa t,yokoyama k,nakajima c,
suzuki y,murata s,ohashi k.immunology,142(4):551-61;aug.2014.
12.非专利文献5:maekawa n,konnai s,ikebuchi r,okagawa t,adachi m,takagi s,kagawa y,nakajima c,suzuki y,murata s,ohashi k.plos one,9(6):e98415;jun.10,2014.
13.非专利文献6:mingala cn,konnai s,ikebuchi r,ohashi k.comp.immunol.microbiol.infect.dis.,34(1):55-63;jan.2011.
技术实现要素:
14.发明所要解决的问题
15.本发明的目的在于,提供一种对于大鼠以外的动物也能够反复施用的抗pd-1抗体。
16.用于解决问题的方法
17.本发明人等确定了能够阻碍牛pd-1与pd-l1的结合的大鼠抗牛pd-1单克隆抗体(5d2)的可变区,并将该可变区基因与牛免疫球蛋白(牛igg1,其中为了抑制adcc活性而对ch2结构域的fcγ受体预想结合部位施加了突变(参照图1、图11。氨基酸序号及突变:251e
→
p,252l
→
v,253p
→
a,254g
→
缺失,348a
→
s,349p
→
s;ikebuchi r,konnai s,okagawa t,yokoyama k,nakajima c,suzuki y,murata s,ohashi k.immunology 2014aug;142(4):551-561.)。)的恒定区基因组合而得到嵌合抗体基因,培养增殖导入了所得的嵌合抗体基因的中国仓鼠卵巢细胞(chinese hamster ovary cell:cho细胞),由此成功地制作出大鼠-牛嵌合抗牛pd-1抗体。此外,确定了大鼠抗牛pd-1单克隆抗体(5d2)的可变区的cdr。本发明是根据这些见解而完成的发明。
18.本发明的主旨如下所示。
19.(1)一种抗pd-1抗体,其包含(a)l链和(b)h链,所述l链具有包含具有qsleysdgyty(序列号16)的氨基酸序列的cdr1、具有gvs的氨基酸序列的cdr2及具有fqathdpdt(序列号17)的氨基酸序列的cdr3的l链可变区、和大鼠以外的动物抗体的l链恒定区,所述h链具有包含具有gfsltsyy(序列号18)的氨基酸序列的cdr1、具有irsggst(序列号19)的氨基酸序列的cdr2及具有artssgyeggfdy(序列号20)的氨基酸序列的cdr3的h链可变区和大鼠以外的动物抗体的h链恒定区。
20.(2)根据(1)中记载的抗体,其中,l链可变区和h链可变区来自于大鼠。
21.(3)根据(2)中记载的抗体,其中,l链可变区为大鼠抗牛pd-1抗体的l链可变区,h链可变区为大鼠抗牛pd-1抗体的h链可变区。
22.(4)根据(3)中记载的抗体,其中,l链可变区具有序列号1的氨基酸序列,h链可变区具有序列号2的氨基酸序列。
23.(5)根据(1)~(4)中任一项记载的抗体,其中,大鼠以外的动物抗体的l链恒定区具有lambda链或kappa链的恒定区的氨基酸序列。
24.(6)根据(1)~(5)中任一项记载的抗体,其中,大鼠以外的动物抗体的h链恒定区具有相当于人的igg4的免疫球蛋白的恒定区的氨基酸序列,或者被导入了使adcc活性和/或cdc活性降低的突变。
25.(7)根据(6)中记载的抗体,其中,大鼠以外的动物为牛,牛抗体的l链恒定区具有
lambda链的恒定区的氨基酸序列,并且牛抗体的h链恒定区被导入了使adcc活性和/或cdc活性降低的突变。
26.(8)根据(7)中记载的抗体,其中,牛抗体的l链恒定区具有序列号3的氨基酸序列,牛抗体的h链恒定区具有序列号4的氨基酸序列。
27.(9)根据(1)~(8)中任一项记载的抗体,其具有两条l链和两条h链的四链结构。
28.(10)一种医药组合物,其包含(1)~(9)中任一项记载的抗体作为有效成分。
29.(11)根据(10)中记载的医药组合物,其用于癌症和/或感染症的预防和/或治疗。
30.(12)根据(11)中记载的医药组合物,其中,癌症和/或感染症选自肿瘤性疾病、白血病、约内氏病、无浆体病(
アナプラズマ
病)、细菌性乳腺炎、真菌性乳腺炎、支原体感染症(例如支原体性乳腺炎、支原体性肺炎等)、结核病、小型梨浆虫病(小型
ピロプラズマ
病)、隐孢子虫病、球虫病、锥虫病及利什曼病中。
31.(13)一种人工基因dna,其包含(a’)编码l链的dna和(b’)编码h链的dna,所述l链具有包含具有qsleysdgyty(序列号16)的氨基酸序列的cdr1、具有gvs的氨基酸序列的cdr2及具有fqathdpdt(序列号17)的氨基酸序列的cdr3的l链可变区、和大鼠以外的动物抗体的l链恒定区,所述h链具有包含具有gfsltsyy(序列号18)的氨基酸序列的cdr1、具有irsggst(序列号19)的氨基酸序列的cdr2及具有artssgyeggfdy(序列号20)的氨基酸序列的cdr3的h链可变区和大鼠以外的动物抗体的h链恒定区。
32.(14)一种载体,其包含(13)中记载的人工基因dna。
33.(15)一种宿主细胞,其被利用(14)中记载的载体进行了转化。
34.(16)一种抗体的制造方法,其包括培养(15)中记载的宿主细胞、并从培养物中采集抗pd-1抗体的操作。
35.(17)一种dna,其编码l链,所述l链具有包含具有qsleysdgyty(序列号16)的氨基酸序列的cdr1、具有gvs的氨基酸序列的cdr2及具有fqathdpdt(序列号17)的氨基酸序列的cdr3的l链可变区、和大鼠以外的动物抗体的l链恒定区。
36.(18)一种dna,其编码h链,所述h链具有包含具有gfsltsyy(序列号18)的氨基酸序列的cdr1、具有irsggst(序列号19)的氨基酸序列的cdr2及具有artssgyeggfdy(序列号20)的氨基酸序列的cdr3的h链可变区、和大鼠以外的动物抗体的h链恒定区。
37.发明效果
38.根据本发明,可以得到新型的抗pd-1抗体。该抗体对于大鼠以外的动物也可以利用。
39.本说明书包含作为本技术的优先权的基础的日本专利申请日本特愿2016-159090及日本特愿2017-099615的说明书和/或附图中记载的内容。
附图说明
40.图1为大鼠-牛嵌合抗牛pd-1抗体ch5d2的氨基酸序列。表示出大鼠抗牛pd-1抗体5d2的l链可变区及h链可变区中的cdr1、cdr2及cdr3区域,此外,还表示出对牛igg1(ch2结构域)施加了突变的氨基酸(氨基酸序号及突变:251e
→
p,252l
→
v,253p
→
a,254g
→
缺失,348a
→
s,349p
→
s)。
41.图2为pdn112载体与大鼠-牛嵌合抗牛pd-1抗体ch5d2的示意图。
42.图3为大鼠-牛嵌合抗牛pd-1抗体ch5d2的产生量及纯化纯度。
43.图4为大鼠-牛嵌合抗牛pd-1抗体ch5d2的结合性。
44.图5为大鼠-牛嵌合抗牛pd-1抗体ch5d2的牛pd-1/pd-l1结合阻碍活性。
45.图6为向blv实验感染牛施用后的大鼠-牛嵌合抗牛pd-1抗体ch5d2的血中浓度的推移。
46.图7为施用了大鼠-牛嵌合抗牛pd-1抗体ch5d2的blv实验感染牛的对blv抗原的t细胞的细胞增殖响应。
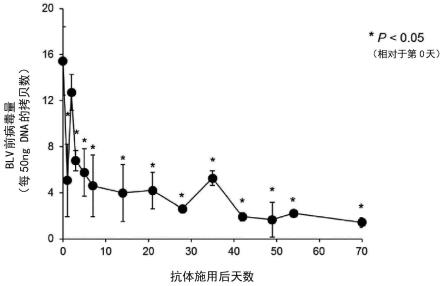
47.图8为施用了大鼠-牛嵌合抗牛pd-1抗体ch5d2的blv实验感染牛的blv前病毒量的变化。
48.图9为大鼠抗牛pd-1抗体5d2与绵羊pd-1的交叉反应性。
49.图10为大鼠抗牛pd-1抗体5d2与水牛的t细胞的交叉反应性。
50.图11为牛igg1恒定区的立体结构及fcγ受体预想结合部位。
51.图12为pdc6载体。
52.图13为大鼠-牛嵌合抗牛pd-1抗体ch5d2 igg1 wt及igg1 adcc-的纯化纯度。
53.图14为大鼠-牛嵌合抗牛pd-1抗体ch5d2 igg1 wt及igg1 adcc-的与各牛fcγ受体的结合性。
具体实施方式
54.以下,对本发明进行详细说明。
55.本发明提供一种抗pd-1抗体,其包含(a)l链和(b)h链,所述l链具有包含具有qsleysdgyty(序列号16)的氨基酸序列的cdr1、具有gvs的氨基酸序列的cdr2及具有fqathdpdt(序列号17)的氨基酸序列的cdr3的l链可变区、和大鼠以外的动物抗体的l链恒定区,所述h链具有包含具有gfsltsyy(序列号18)的氨基酸序列的cdr1、具有irsggst(序列号19)的氨基酸序列的cdr2及具有artssgyeggfdy(序列号20)的氨基酸序列的cdr3的h链可变区和大鼠以外的动物抗体的h链恒定区。
56.大鼠抗牛pd-1抗体5d2(后述)的l链可变区中的cdr1~3分别为包含qsleysdgyty(序列号16)的氨基酸序列的区域、包含gvs的氨基酸序列的区域、包含fqathdpdt(序列号17)的氨基酸序列的区域(参照图1)。
57.另外,大鼠抗牛pd-1抗体5d2的h链可变区中的cdr1~3分别为包含gfsltsyy(序列号18)的氨基酸序列的区域、包含irsggst(序列号19)的氨基酸序列的区域及包含artssgyeggfdy(序列号20)的氨基酸序列的区域(参照图1)。
58.在qsleysdgyty(序列号16)的氨基酸序列、gvs的氨基酸序列及fqathdpdt(序列号17)的氨基酸序列、以及gfsltsyy(序列号18)的氨基酸序列、irsggst(序列号19)的氨基酸序列及artssgyeggfdy(序列号20)的氨基酸序列中,可以缺失、置换或加入1个、2个、3个、4个或5个氨基酸,即使导入这些突变,也可以具有作为pd-1抗体的l链可变区的cdr或h链可变区的cdr的功能。
59.本说明书中,所谓抗体,是除了包括全长抗体以外、还包括fab、f(ab)
’2、scfv、二聚抗体、vh、v
l
、sc(fv)2、双特异性sc(fv)2、微型抗体、scfv-fc单体、scfv-fc二聚体等被低分子化了的抗体的概念。
60.本发明的抗pd-1抗体中,l链可变区和h链可变区可以来自于大鼠。例如,可以是l链可变区为大鼠抗牛pd-1抗体的l链可变区,h链可变区为大鼠抗牛pd-1抗体的h链可变区。
61.将大鼠抗牛pd-1抗体的l链可变区的氨基酸序列及h链可变区的氨基酸序列分别表示于序列号1及2中,然而在序列号1及2的氨基酸序列中,可以缺失、置换或加入1或多个(例如5个以下、至多10个左右)氨基酸,即使导入这些突变,也可以具有作为pd-1抗体的l链可变区或h链可变区的功能。
62.大鼠以外的动物抗体的l链恒定区及h链恒定区可以来自于产生与大鼠抗牛pd-1抗体5d2发生交叉反应的pd-1的动物。
63.在抗体的l链中,有kappa链(卡帕链)和lambda链(兰姆达链),本发明的抗pd-1抗体中,大鼠以外的动物的抗体的l链恒定区无论具有kappa链或lambda链的哪个链的恒定区的氨基酸序列都可以,对于存在比率,就牛、绵羊、猫、狗、马而言是lambda链的一方高,就小鼠、大鼠、人、猪而言是kappa链的一方高。由于认为优选存在比率高的链的一方,因此就牛、绵羊、猫、狗、马而言优选具有lambda链的恒定区的氨基酸序列,就小鼠、大鼠、人、猪而言优选具有kappa链的恒定区的氨基酸序列。
64.大鼠以外的动物的抗体的h链恒定区可以具有相当于人的igg4的免疫球蛋白的恒定区的氨基酸序列。h链根据恒定区的差别被分为γ链、μ链、α链、δ链、ε链,根据该差别分别形成igg、igm、iga、igd、ige的5种类型(同种型)的免疫球蛋白。
65.免疫球蛋白g(igg)占人免疫球蛋白的70-75%,是血浆中最多的单体的抗体。具有两条轻链和两条重链的四链结构。人igg1、igg2、igg4的分子量约为146000,而人igg3的连接fab区域与fc区域的铰链部长,分子量也大到170000。人igg1占人igg的65%左右,人igg2占25%左右,人igg3占7%左右,人igg4占3%左右。平均地分布于血管内外。人igg1由于对效应细胞表面的fc受体、补体因子具有强亲和性,因此诱导抗体依赖性细胞毒作用(adcc),另外,将补体活化而诱导补体依赖性细胞毒作用(cdc)。人igg2和人igg4由于对fc受体、补体因子的亲和性低,因此adcc活性及cdc活性低。
66.免疫球蛋白m(igm)是占人免疫球蛋白的约10%的、结合5个基本的四链结构而得的五聚体的抗体。分子量为970000。通常仅存在于血中,是针对感染微生物最先产生、担任早期免疫的免疫球蛋白。
67.免疫球蛋白a(iga)占人免疫球蛋白的10-15%。分子量为160000。分泌型iga是结合2个iga而得的二聚体的抗体。iga1存在于血清、鼻涕、唾液、母乳中,在肠液中存在大量iga2。
68.免疫球蛋白d(igd)是人免疫球蛋白的1%以下的单体的抗体。存在于b细胞表面,参与抗体产生的诱导。
69.免疫球蛋白e(ige)是人免疫球蛋白的0.001%以下的仅极微量存在的单体的抗体。被认为参与针对寄生虫的免疫反应,而在寄生虫稀少的发达国家中,特别是与支气管哮喘、过敏有很大关系。
70.就狗而言,作为igg的h链,鉴定出igg-a(相当于人igg2)、igg-b(相当于人igg1)、igg-c(相当于人igg3)、igg-d(相当于人igg4)的序列。本发明的抗体中,优选不同时具有adcc活性、cdc活性的iggh链恒定区(就人而言是igg4)。在没有鉴定出相当于人igg4的免疫球蛋白的恒定区的情况下,可以使用通过对相当于人igg1的免疫球蛋白的该区域施加突变
而不同时具有adcc活性、cdc活性的区域。
71.就牛而言,作为igg的h链,鉴定出igg1、igg2、igg3的序列。本发明的抗体中,优选不同时具有adcc活性、cdc活性的iggh链恒定区(就人而言是igg4)。已知人igg1的恒定区就野生型而言具有adcc活性及cdc活性,而通过对特定的部分施加氨基酸置换、缺失,可以使它们的活性降低。在牛中,由于没有鉴定出相当于人igg4的免疫球蛋白的恒定区,因此可以对相当于人igg1的免疫球蛋白的该区域施加突变并使用该区域。作为其一例,将对牛抗体的h链恒定区(igg1链,genbank:x62916)的ch2结构域施加了突变的氨基酸序列和核苷酸序列(将密码子最佳化了的序列)分别表示于序列号4及8中。
72.更优选如下的抗pd-1抗体,即,牛抗体的l链恒定区具有lambda链的恒定区的氨基酸序列,并且牛抗体的h链恒定区被导入了使adcc活性和/或cdc活性降低的突变。
73.本发明的抗pd-1抗体包含大鼠-牛嵌合抗体、牛化抗体、完全牛型抗体,然而动物并不限定为牛,可以例示出入、狗、猪、猴、小鼠、猫、马、山羊、绵羊、水牛、兔、仓鼠、豚鼠等等。
74.例如,本发明的抗pd-1抗体可以是牛抗体的l链恒定区具有序列号3的氨基酸序列、牛抗体的h链恒定区具有序列号4的氨基酸序列的抗pd-1抗体。
75.在序列号3及4的氨基酸序列中,可以缺失、置换或加入1或多个(例如5个以下、至多10个左右)氨基酸,即使导入这些突变,也可以具有作为抗体的l链恒定区或h链恒定区的功能。
76.本发明的抗pd-1抗体可以是具有两条l链和两条h链的四链结构。
77.本发明的抗pd-1抗体可以如下所示地制造。合成包含鉴定了的大鼠抗牛pd-1抗体的可变区序列和大鼠以外的动物(例如牛等)的抗体(优选为对相当于人igg1的免疫球蛋白的该区域施加突变、使adcc活性和/或cdc活性降低的抗体)的恒定区序列的人工基因,将该人工基因插入载体(例如质粒)后,导入宿主细胞(例如cho细胞等哺乳类细胞),并培养该宿主细胞,由此从培养物中采集抗体。
78.将本发明人等鉴定了的大鼠抗牛pd-1抗体的l链可变区的氨基酸序列和核苷酸序列分别表示于序列号1及5中。此外,将密码子最佳化后的核苷酸序列表示于序列号11中。
79.将本发明人等鉴定了的大鼠抗牛pd-1抗体的h链可变区的氨基酸序列和核苷酸序列分别表示于序列号2及6中。此外,将密码子最佳化后的核苷酸序列表示于序列号12中。
80.将牛抗体的l链恒定区(lambda链,genbank:x62917)的氨基酸序列和核苷酸序列分别表示于序列号3及7中。此外,将密码子最佳化后的核苷酸序列表示于序列号13中。
81.将牛抗体的h链恒定区(igg1链,改变genbank:x62916)的氨基酸序列和核苷酸序列(密码子最佳化后)分别表示于序列号4及8中。
82.另外,序列号9表示包含大鼠抗牛pd-1抗体的l链可变区和牛抗体的l链恒定区(lambda链,genbank:x62917)的嵌合l链的氨基酸序列。将包含大鼠抗牛pd-1抗体的l链可变区和牛抗体的l链恒定区(lambda链,genbank:x62917)的嵌合l链的核苷酸序列(密码子最佳化后)表示于序列号14中。
83.序列号10表示包含大鼠抗牛pd-1抗体的h链可变区和牛抗体的h链恒定区(igg1链,改变genbank:x62916)的嵌合h链的氨基酸序列。将包含大鼠抗牛pd-1抗体的h链可变区和牛抗体的h链恒定区(igg1链,改变genbank:x62916)的嵌合h链的核苷酸序列(密码子最
佳化后)表示于序列号15中。
84.除此以外,大鼠以外的动物的l链恒定区及h链恒定区的氨基酸序列和核苷酸序列可以从公知的数据库获得,可以利用这些序列。
85.将牛的l链恒定区及h链恒定区的氨基酸序列和核苷酸序列集中整理于下述的表中。
86.(表)
87.[0088][0089][0090]
将绵羊、水牛、人的l链恒定区及h链恒定区的氨基酸序列和核苷酸序列集中整理
于下述的表中。
[0091]
(表)
[0092]
[0093]
[0094][0095]
在序列号3、21~28、37、39、41、43、45、47、49、51、53、55、57、以及59的氨基酸序列中,可以缺失、置换或加入1或多个(例如5个以下、至多10个左右)氨基酸,即使导入这些突变,也可以具有作为ig重链或轻链的恒定区的功能。
[0096]
已知人igg1的恒定区就野生型而言具有adcc活性及cdc活性,然而通过对特定的部分施加氨基酸置换、缺失,可以降低它们的活性。在人以外的动物中,在没有鉴定出相当于人igg4的免疫球蛋白的恒定区的情况下,可以使用对相当于人igg1的免疫球蛋白的该区域施加突变、降低了adcc活性及cdc活性的区域。
[0097]
本发明提供一种人工基因dna,其包含(a’)编码l链的dna和(b’)编码h链的dna,所述l链具有包含具有qsleysdgyty(序列号16)的氨基酸序列的cdr1、具有gvs的氨基酸序列的cdr2及具有fqathdpdt(序列号17)的氨基酸序列的cdr3的l链可变区、和大鼠以外的动物抗体的l链恒定区,所述h链具有包含具有gfsltsyy(序列号18)的氨基酸序列的cdr1、具有irsggst(序列号19)的氨基酸序列的cdr2及具有artssgyeggfdy(序列号20)的氨基酸序列的cdr3的h链可变区和大鼠以外的动物抗体的h链恒定区。另外,本技术发明还提供一种编码l链的dna,所述l链具有包含具有qsleysdgyty(序列号16)的氨基酸序列的cdr1、具有gvs的氨基酸序列的cdr2及具有fqathdpdt(序列号17)的氨基酸序列的cdr3的l链可变区、和大
鼠以外的动物抗体的l链恒定区。此外,本发明还提供一种编码h链的dna,所述h链具有包含具有gfsltsyy(序列号18)的氨基酸序列的cdr1、具有irsggst(序列号19)的氨基酸序列的cdr2及具有artssgyeggfdy(序列号20)的氨基酸序列的cdr3的h链可变区、和大鼠以外的动物抗体的h链恒定区。
[0098]
对于(a)具有包含具有qsleysdgyty(序列号16)的氨基酸序列的cdr1、具有gvs的氨基酸序列的cdr2及具有fqathdpdt(序列号17)的氨基酸序列的cdr3的l链可变区、和大鼠以外的动物抗体的l链恒定区的l链、以及(b)具有包含具有gfsltsyy(序列号18)的氨基酸序列的cdr1、具有irsggst(序列号19)的氨基酸序列的cdr2及具有artssgyeggfdy(序列号20)的氨基酸序列的cdr3的h链可变区和大鼠以外的动物抗体的h链恒定区的h链,已经在前面叙述。(a’)的dna为编码(a)的l链的dna(基因),(b’)的dna为编码(b)的h链的dna(基因),包含(a’)的dna和(b’)的dna的人工基因dna可以使用市售的合成机合成。也可以向人工基因dna加入限制性酶识别部位、kozak序列、poly(a)加尾信号序列、启动子序列、内含子序列等。
[0099]
另外,本发明还提供包含所述人工基因dna的载体。
[0100]
作为载体,可以使用大肠杆菌来源的质粒(例如pbr322、pbr325、puc12、puc13)、枯草杆菌来源的质粒(例如pub110、ptp5、pc194)、酵母来源质粒(例如psh19、psh15)、λ噬菌体等噬菌体、逆转录病毒、痘苗病毒等动物病毒、杆状病毒等昆虫病原病毒等。后述的实施例中,使用了pdn1 12(marzi a,yoshida r,miyamoto h,ishijima m,suzuki y,higuchi m,matsuyama y,igarashi m,nakayama e,kuroda m,saijo m,feldmann f,briningd,feldmann h,takada a.plos one,7:e36192,apr.27,2012;ikebuchi r,konnai s,okagawa t,yokoyama k,nakajima c,suzuki y,murata s,ohashi k.immunology,142(4):551-561,aug.2014.)。
[0101]
也可以向载体中加入启动子、增强子、剪接信号、poly(a)加尾信号、内含子序列、选择标记、sv40复制起始点等。
[0102]
此外,本发明还提供利用所述载体进行了转化的宿主细胞。通过培养该宿主细胞,并从培养物中采集抗体,可以制造抗pd-1抗体。由此,本发明还提供一种抗体的制造方法,该制造方法包括培养所述宿主细胞、并从培养物中采集抗pd-1抗体的操作。本发明的抗体的制造方法中,可以将整合了包含编码l链的dna和编码h链的dna的人工基因dna的载体转染到宿主细胞中,也可以将整合了编码l链的dna的载体和整合了编码h链的dna的载体共转染到宿主细胞中。
[0103]
作为宿主细胞,可以例示出细菌细胞(例如埃希菌属细菌、芽孢杆菌属细菌、枯草杆菌等)、真菌细胞(例如酵母、曲霉等)、昆虫细胞(例如s2细胞、sf细胞等)、动物细胞(例如cho细胞、cos细胞、hela细胞、c127细胞、3t3细胞、bhk细胞、hek293细胞等)、植物细胞等。其中,优选作为二氢叶酸还原酶缺损细胞的cho-dg44细胞(cho-dg44(dhfr-/-))。
[0104]
要将重组载体导入宿主,可以利用分子克隆第二版,j.sambrook et al.,cold springharborlab.press,1989中记载的方法(例如磷酸钙法、deae-葡聚糖法、转染法、显微注射法、脂质体转染法、电穿孔法、转化法、刮棒负载法(
スクレ
一
プロ
一
デイング
法)、鸟枪法(
ショツトガン
法)等)或感染来进行。
[0105]
可以用培养基培养转化体,从培养物中采集本发明的抗pd-1抗体。在抗体被分泌
到培养基中的情况下,只要回收培养基并从该培养基中分离、纯化抗体即可。在抗体产生于受到转化的细胞内的情况下,只要将该细胞溶解并从其溶解物中分离、纯化抗体即可。
[0106]
作为培养基,可以例示出opticho培养基、dynamis培养基、cd cho培养基、acticho培养基、forticho培养基、ex-cell cd cho培养基、balancd cho培养基、procho5培养基、cellvento cho-100培养基等,然而并不限定于它们。
[0107]
培养基的ph根据所培养的细胞而不同,然而一般而言为ph6.8~7.6,多数情况下适合为ph7.0~7.4。
[0108]
在所培养的细胞为cho细胞的情况下,cho细胞的培养可以使用本领域人员公知的方法进行。例如,通常可以在气相的co2浓度为0-40%、优选为2-10%的气氛下、在30-39℃、优选在37℃左右进行培养。
[0109]
适合的培养期间通常为1天~3个月,优选为1天~3周。
[0110]
抗体的分离及纯化可以利用公知的方法进行。作为公知的分离、纯化法,可以使用盐析、溶剂沉淀法等利用溶解度的差的方法、透析法、超滤法、凝胶过滤法、以及sds-聚丙烯酰胺凝胶电泳法等利用分子量的差的方法、离子交换色谱等利用电荷的差的方法、亲和层析等利用特异性亲和性的方法、反相高速液体色谱等利用疏水性的差的方法、等电点电泳法等利用等电点的差的方法等。
[0111]
本发明的抗pd-1抗体可以作为动物用或人用的抗体药利用。由此,本发明提供包含上述的抗pd-1抗体作为有效成分的医药组合物。
[0112]
本发明的医药组合物可以用于癌症和/或感染症的预防和/或治疗。作为癌症和/或感染症,可以例示出肿瘤性疾病(例如恶性黑色素瘤、肺癌、胃癌、肾癌、乳腺癌、膀胱癌、食道癌、卵巣癌等)、白血病、约内氏病、无浆体病、细菌性乳腺炎、真菌性乳腺炎、支原体感染症(例如支原体性乳腺炎、支原体性肺炎等)、结核病、小型梨浆虫病、隐孢子虫病、球虫病、锥虫病及利什曼病等,然而并不限定于它们。
[0113]
可以将本发明的抗pd-1抗体溶解于pbs等缓冲液、生理盐水、灭菌水等中,根据需要用过滤器等进行过滤灭菌后,利用注射向受试动物(也包括人)施用。另外,可以向该溶液中添加添加剂(例如着色剂、乳化剂、悬浮剂、表面活性剂、溶解助剂、稳定化剂、保存剂、抗氧剂、缓冲剂、等渗剂、ph调节剂等)等。作为施用路径,可以是静脉、肌肉、腹腔、皮下、皮内施用等,另外,也可以经鼻、经口施用。
[0114]
本发明的抗pd-1抗体的施用量、施用的次数及频率根据受试动物的症状、年龄、体重、施用方法、施用形态等而不同,例如通常可以对每一成年动物以至少一次、可以获得所期望的效果的频率施用0.1~100mg/kg体重、优选1~10mg/kg体重。
[0115]
本发明的医药组合物可以单独使用,也可以与外科手术、放疗、癌症疫苗等其他免疫细胞疗法、分子靶向治疗药组合使用。由此可以期待协同效应。
[0116]
实施例
[0117]
以下,基于实施例对本发明进行详细说明,然而本发明并不限定于这些实施例。
[0118]
〔实施例1〕大鼠-牛嵌合抗牛pd-1抗体的建立
[0119]
序论
[0120]
免疫抑制受体programmed death 1(pd-1)和作为其配体的programmed death ligand 1(pd-l1)是由京都大学本庶佑等人鉴定为抑制过度的免疫响应、与免疫耐受密切
相关的因子的分子。近年来还阐明参与肿瘤中的免疫抑制。本实施例中,以建立对牛的感染症的新型治疗法为目的,制作了大鼠-牛嵌合抗牛pd-1抗体ch5d2,并确认了体外(in vitro)及体内(in vivo)的效果,所述大鼠-牛嵌合抗牛pd-1抗体ch5d2是培养增殖表达嵌合抗体基因的中国仓鼠卵巢细胞(chinese hamsterovary cell:cho细胞)而得,所述嵌合抗体基因是将能够阻碍牛pd-1与pd-l1的结合的大鼠抗牛pd-1单克隆抗体5d2的可变区基因、和牛免疫球蛋白(牛igg1及igλ。其中,为了抑制adcc活性,对牛igg1 ch2结构域的fcγ受体预想结合部位施加了突变(图1、图11)。氨基酸序号及突变:250e
→
p,251l
→
v,252p
→
a,253g
→
缺失,347a
→
s,348p
→
s;ikebuchi r,konnai s,okagawa t,yokoyama k,nakajima c,suzuki y,murata s,ohashi k.immunology,142(4):551-561;aug.2014.)的恒定区基因组合而得。
[0121]
材料、方法、以及实验结果
[0122]
2.1.牛pd-1及pd-l1表达细胞的构建
[0123]
对牛pd-1基因(genbank登记号ab510901;ikebuchi r,konnai s,sunden y,onuma m,ohashi k.microbiol.immunol.,54(5):291-298;may 2010.)及牛pd-l1基因(genbank登记号ab510902;ikebuchi r,konnai s,shirai t,sunden y,murata s,onuma m,ohashi k.vet.res.,42:103;sep.26,2011.)确定cdna全长的碱基序列,根据其基因信息制作出牛pd-1或pd-l1表达细胞。首先,为了制作牛pd-1或pd-l1表达质粒,以所合成的牛外周血单核细胞(pbmc)来源cdna为模板,使用向5
′
末端侧加入限制性酶noti及hindiii(牛pd-1)、或nhei及xhoi(牛pd-l1)识别部位的引物(bopd-1-myc f及r、或bopd-l1-egfp f及r)进行了pcr。对所得的pcr产物利用noti(takara公司)及hindiii(takara公司;牛pd-1)、或nhei(takara公司)及xhoi(takara公司;牛pd-l1)进行处理后,使用fastgene gel/pcr提取试剂盒(nippon genetics公司)进行纯化,导入进行了同样的限制性酶处理的pcmv-tag1载体(agilent technologies公司;牛pd-1)或pegfp-n2载体(clontech公司;牛pd-l1)中,进行了克隆。使用qiagen plasmid midi kit(qiagen公司)提取所得的目标的表达质粒,在用于实验前以-30℃保存。以后,将所制作的表达质粒表示为pcmv-tag1-bopd-1或pegfp-n2-bopd-l1。
[0124]
引物(bopd-1-myc f):
[0125]
atatgcggccgcatggggaccccgcgggcgct(序列号61);
[0126]
引物(bopd-1-myc r):
[0127]
gcgcaagctttcagaggggccaggagcagt(序列号62);
[0128]
引物(bopd-l1-egfp f):
[0129]
ctagctagcaccatgaggatatatagtgtcttaac(序列号63);
[0130]
引物(bopd-l1-egfp r):
[0131]
caatctcgagttacagacagaagatgactgc(序列号64)。
[0132]
依照以下的步骤,制作出牛pd-1表达细胞。首先,使用lipofectamine ltx(invitrogen公司)将2.5μg的pcmv-tag1-bopd-1导入4
×
106个的cho-dg44细胞中。48小时后,将培养基更换为包含g418(enzo life science公司)800μg/ml、glutamax添加剂(life technologies公司)20ml/l、10%pluronic f-68(life technologies公司)18ml/1的cd dg44培养基(life technologies公司),进行了选择。使所得的表达细胞与大鼠抗牛pd-1抗
体5d2在室温下反应,清洗后,与抗大鼠igg微珠标记抗体(miltenyi biotec公司)在室温下进一步反应。使用auto macs(miltenyi biotec公司)分离高表达牛pd-1的细胞,为了进一步提高纯度,利用同様的步骤进行了再分离。对所制作出的表达细胞利用极限稀释法进行克隆,得到牛pd-1高表达cho dg44细胞(牛pd-1表达细胞)。
[0133]
依照以下的步骤,制作出牛pd-l1膜表达细胞。首先,使用lipofectamine ltx(invitrogen公司)将2.5μg的pcmv-tag1-bopd-l1或作为阴性对照的pegfp-n2导入4
×
106个的cho-dg44细胞中。48小时后,将培养基更换为包含g418(enzo life science公司)800μg/ml、glutamax添加剂(life technologies公司)20ml/l、10%pluronic f-68(life technologies公司)18ml/l的cd dg44培养基(life technologies公司),在进行选择的同时利用极限稀释法进行了克隆(牛pd-l1表达细胞)。为了确认所制作出的表达细胞中的牛pd-l1的表达,利用倒立型共聚焦激光显微镜lsm700(zeiss公司),使egfp的细胞内局部可视化。
[0134]
2.2.可溶性牛pd-1的构建
[0135]
依照以下的步骤,构建出牛pd-1-ig表达质粒。制成使牛pd-1(genbank登记号ab510901)的信号肽及细胞外区域与已知的牛igg1(genbank登记号x62916)的恒定区结合了的基因序列,对cho细胞进行密码子的最佳化后,以将选择性酶(noti)识别序列、kozak序列、牛pd-1信号肽序列、牛pd-1基因细胞外区域序列、牛igg1 fc区域序列、选择性酶(xbai)识别序列依照上述的顺序配置的方式进行了基因合成。需要说明的是,牛igg1为了抑制adcc活性,对ch2结构域的fcγ受体预想结合部位施加了突变(突变插入部位:185e
→
p,186l
→
v,187p
→
a,189g
→
缺失,281a
→
s,282p
→
s;ikebuchi r,konnai s,okagawa t,yokoyama k,nakajima c,suzuki y,murata s,ohashi k.immunology,142(4):551-561;aug.2014.该论文的figure 2中给出了pd-1-ig的氨基酸序列、突变插入部位)。将所合成出的基因链利用noti(takara公司)及xbai(takara公司)处理后,使用fastgene gel/pcr提取试剂盒(nippon genetics公司)纯化,整合到进行了同样的选择性酶处理的表达用载体pdn11(由北海道大学人畜共患感染症研究中心铃木定彦教授分与)的克隆位点(位于pcmv下游、inrbg与pabgh之间的noti及xbai选择性酶识别序列)中,构建出牛pd-1-ig表达载体。使用qiagen plasmid midi kit(qiagen公司)纯化表达质粒,在用于实验前以-30℃保存。以后,将所制作的表达质粒表示为pdn11-bopd-1-ig。
[0136]
依照以下的步骤,构建出牛pd-1-his表达质粒。以扩增牛pd-1(genbank登记号ab510901)的信号肽及细胞外区域的方式,设计了向5
′
末端侧加入选择性酶noti及xhoi识别部位的引物(bopd-1-his f及r)。需要说明的是,向反向引物中加入了编码6
×
组氨酸(his)标签的基因序列。以所合成出的牛pbmc来源cdna为模板进行pcr,对各个pcr产物利用noti(takara公司)及xhoi(takara公司)进行处理后,使用fastgene gel/pcr提取试剂盒(nippon genetics公司)纯化,导入进行了同样的选择性酶处理的pcxn2.1(+)载体(niwa h,yamamura k,miyazaki j.gene,108(2):193-199;dec.15,1991;由顺天堂大学大学院医学研究科横沟岳彦教授分与),进行了克隆。将表达质粒利用fastgene xpress plasmid plus kit(nippon genetics公司)纯化,在用于实验前以-30℃保存。以后,将所制作的表达质粒表示为pcxn2.1-bopd-1-his。
[0137]
引物(bopd-1-his f):
[0138]
ataagaatgcggccgccaccatggggaccccgcgggcgct(序列号65);
[0139]
引物(bopd-1-his r):
[0140]
gccctcgagttaatggtgatggtgatggtggatgaccaggctctgcatct(序列号66)。
[0141]
依照以下的步骤,制作出可溶性牛pd-1-ig表达细胞。使用lipofectamine ltx(invitrogen公司)将2.5μg的pdn11-bopd-1-ig导入4
×
106个cho-dg44细胞中。48小时后,将培养基更换为包含g418(enzo life science公司)800μg/ml、glutamax添加剂(life technologies公司)20ml/l的cd opticho培养基(life technologies公司),培养3周后进行了选择。利用使用了抗牛iggf(c)兔多克隆抗体(rockland公司)的elisa法测定出所得的细胞株的培养上清液中的fc融合重组蛋白的浓度,进行了高表达fc融合重组蛋白的细胞株的挑选。将所得的高表达细胞株转移到不包含g418的培养基中,进行14天振荡培养后回收培养上清液。将包含fc融合重组蛋白的培养上清液用centricon plus-70(millipore公司)超滤后,使用ab-capcherextra(protenova公司)纯化fc融合重组蛋白。纯化后,利用pd-10脱盐柱(ge healthcare公司)将缓冲液置换为磷酸缓冲生理盐水(pbs;ph 7.4),在用于实验前以-30℃保存(牛pd-1-ig)。利用使用了抗牛iggf(c)兔多克隆抗体(rockland公司)的elisa法测定出纯化后的牛pd-1-ig的浓度。在elisa的各清洗操作中使用了全自动洗板机biowasher 50(ds pharma biomedical公司),在吸光度的测定中使用了微板读数仪mtp-650fa(corona电气公司)。
[0142]
依照以下的步骤,制作出可溶性牛pd-1-his表达细胞。使用expifectamine(life technologies公司)将30μg的pcxn2.1-bopd-1-his导入7.5
×
107个expi293f细胞(life technologies公司)中,进行7天振荡培养后回收培养上清液。从培养上清液中使用talon金属亲和树脂(clontech公司;牛pd-1-his)纯化重组蛋白。纯化后,利用pd minitrap g-25(ge healthcare公司)将缓冲液置换为pbs(ph 7.4),在用于实验前以-30℃保存(牛pd-1-his)。对纯化后的牛pd-1-his浓度利用使用nanodrop8000分光光度计(thermo fisher scientific公司)测定出的吸光度(280nm)进行定量。
[0143]
2.3.大鼠抗牛pd-1单克隆抗体产生细胞的制作
[0144]
将牛pd-1-ig(前述)对大鼠的足跖进行免疫,使用髂骨淋巴结法建立杂交瘤,得到大鼠抗牛pd-1单克隆抗体产生杂交瘤5d2株。对于大鼠抗牛pd-1单克隆抗体的建立方法,在以下的非专利文献中记载有其详情(ikebuchi r,konnai s,okagawa t,yokoyama k,nakajima c,suzuki y,murata s,ohashi k.vet.res.,44:59;jul.22,2013)。
[0145]
2.4.大鼠-牛嵌合抗牛pd-1抗体表达载体的制作
[0146]
建立以大鼠抗牛pd-1抗体5d2作为抗体可变区融合了牛igg1及牛igλ的抗体恒定区的大鼠-牛嵌合抗牛pd-1抗体ch5d2。
[0147]
首先,从产生大鼠抗牛pd-1抗体5d2的杂交瘤中利用race法鉴定出可变区(重链及轻链)的基因。然后,制成使大鼠抗牛pd-1抗体5d2的重链及轻链可变区序列与已知的牛igg1(重链;改变genbank登记号x62916)及牛igλ(轻链;genbank登记号x62917)的恒定区结合的基因序列,进行了密码子最佳化(序列号9及10(氨基酸序列)、序列号14及15(密码子最佳化后核苷酸序列))。需要说明的是,牛igg1中为了抑制adcc活性,对ch2结构域的fcγ受体预想结合部位施加了突变(参照图1、图11。氨基酸序号及突变:251e
→
p,252l
→
v,253p
→
a,254g
→
缺失,348a
→
s,349p
→
s;ikebuchi r,konnai s,okagawa t,yokoyama k,
nakajima c,suzuki y,murata s,ohashi k.immunology,142(4):551-561;aug.2014.)。然后,以将noti选择性酶识别序列、kozak序列、嵌合抗体轻链序列、poly(a)加尾信号序列(pabgh)、启动子序列(pcmv)、saci选择性酶识别序列、内含子序列(inrbg)、kozak序列、嵌合抗体重链序列、xbai选择性酶识别序列依照上述的顺序配置的方式人工地合成出基因。将所合成出的基因链利用noti(takara公司)及xbai(takara公司)处理后,使用fastgene gel/pcr提取试剂盒(nippon genetics公司)纯化,导入进行了同样的选择性酶处理的表达质粒pdn112(由北海道大学人畜共患感染症研究中心铃木定彦教授分与)的克隆位点(位于pcmv下游、inrbg与pabgh之间的noti及xbai选择性酶识别序列),进行了克隆(图2)。使用qiagen plasmid midi kit(qiagen公司)提取所得的目标的表达质粒,在用于实验前以-30℃保存。以后,将所制作的表达质粒表示为pdn112-bopd-1ch5d2。
[0148]
2.5.大鼠-牛嵌合抗牛pd-1抗体的表达(图3)
[0149]
将所制作出的pdn112-bopd-1ch5d2使用lipofectamine ltx(life technologies公司)导入作为二氢叶酸还原酶缺损(dfhr-/-)细胞的cho-dg44细胞。48小时后,将培养基更换为包含2mm glutamax添加剂(life technologies公司)及g418硫酸盐800μg/ml(enzo life science公司)的cd opticho培养基(life technologies公司),培养3周而进行了基于表达细胞的选择及极限稀释法的克隆。然后,利用使用了抗牛iggf(c)兔多克隆抗体(rockland公司)的斑点印迹法及elisa法测定培养上清液中所含的嵌合抗体的浓度,筛选出高表达克隆。此外,对筛选出的大鼠-牛嵌合抗牛pd-1抗体高表达克隆在包含60nm的甲氨蝶呤(mtx;和光纯药工业公司)的培养基中施加负荷,由此进行了基因扩增处理。将如上所述地建立的大鼠-牛嵌合抗牛pd-1抗体稳定表达细胞转移到不包含mtx的cd opticho培养基中,进行14天的振荡培养(125rpm,37℃,5%co2)。利用使用了抗牛iggf(c)兔多克隆抗体(rockland公司)的elisa法,对培养上清液中的嵌合抗体产生量进行定量。需要说明的是,在elisa的各清洗操作中使用了全自动洗板机biowasher50(ds pharma biomedical公司),在吸光度的测定中使用了微板读数仪mtp-650fa(corona电气公司)。将第14天的培养上清液以10000g离心10分钟而除去细胞后,将离心上清液通过steritop-gp 0.22μm过滤器(millipore公司)而灭菌,在用于纯化前以4℃保存。
[0150]
将结果表示于图3a中。大鼠-牛嵌合抗牛pd-1抗体表达细胞株当中产生量最高的克隆在14天的振荡培养中向培养上清液中分泌出91.7mg/l的嵌合抗体。
[0151]
2.6.大鼠-牛嵌合抗牛pd-1抗体的纯化
[0152]
从利用上述的方法准备的培养上清液中,使用ab capcherextra(protenova公司)将各嵌合抗体纯化。与树脂的结合使用开管柱法,作为平衡化缓冲液及清洗缓冲液使用1.5m甘氨酸/3m nacl(ph 8.0)。作为洗脱缓冲液使用0.1m甘氨酸-hcl(ph 2.8),作为中和缓冲液使用1m tris(ph 9.0)。对纯化了的抗体使用pd-10脱盐柱(ge healthcare公司)及amicon ultra-15(50kda、millipore公司)进行了置换为pbs(ph 7.4)的缓冲液置换及浓缩。将纯化了的嵌合抗体通过0.22μm注射器过滤器(pall life sciences公司)而进行灭菌,在用于实验前以4℃保存。
[0153]
2.7.大鼠-牛嵌合抗牛pd-1抗体的纯化纯度的确认(图3)
[0154]
为了确认纯化了的大鼠-牛嵌合抗牛pd-1抗体的纯度,利用sds-page及cbb染色进行了抗体蛋白质的检测。将纯化了的大鼠-牛嵌合抗牛pd-1抗体ch5d2悬浮于laemmli样品
缓冲液(bio-rad公司)中,在还原条件下(利用2-巯基乙醇(sigma-aldrich公司)还原)或非还原条件下进行变性处理(95℃、5分钟)。对所制备的样品使用10%聚丙烯酰胺凝胶进行电泳。此时,作为分子量标记使用了precision plus protein全蓝蛋白质标准品(bio-rad公司)。电泳后,利用quick-cbb(和光纯药工业公司)进行凝胶的染色,接下来在蒸馏水中进行脱色。
[0155]
将结果表示于图3b中。在还原条件下在25kda(轻链)及50kda(重链)的预估的位置确认到大鼠-牛嵌合抗牛pd-1抗体的条带,在非还原条件下在150kda的预估的位置确认到大鼠-牛嵌合抗牛pd-1抗体的条带。
[0156]
2.8.大鼠-牛嵌合抗牛pd-1抗体的结合特异性(图4)
[0157]
利用流式细胞仪法确认了大鼠-牛嵌合抗牛pd-1抗体与牛pd-1表达细胞(前述)特异性地结合。首先,使大鼠抗牛pd-1抗体5d2或大鼠-牛嵌合抗牛pd-1抗体ch5d2与牛pd-1表达细胞在室温下反应30分钟。清洗后,使别藻蓝蛋白(apc)标记抗大鼠ig山羊抗体(southernbiotech公司)或alexa fluor 647标记抗牛igg(h+l)山羊f(ab
′
)2(jackson immunoresearch公司)在室温下反应30分钟。作为阴性对照抗体,使用了大鼠igg2a(κ)同种型对照(bd biosciences公司)或牛igg1抗体(bethyl公司)。清洗后,利用facs verse(bd biosciences公司)检测出结合于细胞表面的各大鼠抗体或大鼠-牛嵌合抗体。需要说明的是,在所有的清洗操作及抗体的稀释中,使用了加入有1%牛血清白蛋白(sigma-aldrich公司)的pbs。
[0158]
将实验结果表示于图4中。显示出大鼠-牛嵌合抗牛pd-1抗体ch5d2与大鼠抗牛pd-1抗体5d2同样地与牛pd-1表达细胞结合。
[0159]
2.9.大鼠-牛嵌合抗牛pd-1抗体的pd-1结合亲和性
[0160]
利用使用了分子间相互作用分析装置(biacore x100;ge healthcare公司)的表面等离子体共振法,计测出大鼠抗pd-1抗体5d2及大鼠-牛嵌合抗牛pd-1抗体ch5d2与牛pd-1的结合亲和性。将牛pd-1-his(前述)作为配体固定于传感器芯片cm5(ge healthcare公司),以大鼠抗pd-1抗体5d2或大鼠-牛嵌合抗牛pd-1抗体ch5d2作为待分析物使之反应,进行了单动力学(
シングルカイネテイクス
)分析。在相同条件下重复3次实验,在各个实验中确定结合常数(kd值)和解离常数(ka值),求出结合亲和性(kd值)。
[0161]
将实验结果表示于下述的表中。大鼠-牛抗牛pd-1嵌合抗体与pd-1蛋白的结合亲和性是与大鼠抗牛pd-1抗体5d2相同的程度,在统计学上观察不到差别(p>0.05;welch的t检验)。
[0162][0163]
2.10.大鼠-牛嵌合抗pd-1抗体的牛pd-1/pd-l1结合阻碍活性(图5)
[0164]
使用牛pd-l1表达细胞(前述)及牛pd-1-ig(前述),进行了利用抗pd-1抗体的牛pd-1/pd-l1结合阻碍试验。首先,向96孔板中添加使用lightning-link type a生物素标记试剂盒(innova biosciences公司)进行了生物素标记的牛pd-1-ig(终浓度5μg/ml),使之与终浓度(0、0.39、0.78、1.56、3.12、6.25、12.5、25、50μg/ml)的大鼠抗牛pd-1抗体5d2或大
biosciences公司)。需要说明的是,在所有的清洗操作及抗体的稀释中,使用了加入有1%牛血清白蛋白(sigma-aldrich公司)的pbs。对于增殖了的t细胞(cfse
low
细胞)的比例,使用dunnett的方法进行了统计学检验。
[0175]
将实验结果表示于图7中。利用大鼠-牛嵌合抗牛pd-1抗体的施用,cd4
+
t细胞中的blv特异性细胞增殖响应与施用前相比,从刚刚施用后起在统计学上明显地增加。
[0176]
2.15.blv前病毒量的推移(图8)
[0177]
从分离出的牛pbmc中使用wizard dna purification kit(promega公司)提取dna。对所提取出的dna的浓度以使用nanodrop 8000分光光度计(thermo fisher scientific公司)测定出的吸光度(260nm)作为基准进行定量。为了测定pbmc中的blv前病毒量,使用cycleave pcr reaction mix sp(takara公司)及牛白血病病毒检测用探针/引物/阳性对照(takara公司)进行实时pcr。测定中使用了lightcycler480system ii(roche diagnosis公司)。对于测定出的前病毒量,使用dunnett的方法进行了统计学检验。
[0178]
将实验结果表示于图8中。利用大鼠-牛嵌合抗牛pd-1抗体的施用,pbmc中的blv前病毒量与施用前相比,从刚刚施用后起在统计学上明显地减少,保持低的值不变地推移至临床试验结束时(第70天)。
[0179]
〔实施例2〕抗pd-1抗体向其他动物种类的应用
[0180]
1.材料、方法、以及实验结果
[0181]
1.1.绵羊及水牛pd-1基因的鉴定
[0182]
为了确定绵羊及水牛pd-1cdna编码区(cds)全长,首先设计以牛及绵羊pd-1基因的碱基序列(genbank登记号bc123854及xm_012176227)为基础扩增基因的cds全长的引物(ovpd-1cds f及r、bupd-1cds f1、r1、f2及r2),以所合成出的绵羊或水牛pbmc来源cdna作为模板实行了pcr法。对所得的扩增产物,依照常法利用毛细管测序仪确定出碱基序列(mingala cn,konnai s,ikebuchi r,ohashi k.comp.immunol.microbiol.infect.dis.,34(1):55-63;jan.2011.鉴定水牛pd-1基因的论文)。
[0183]
引物(ovpd-1cds f):atggggaccccgcgggcgcc(序列号67);
[0184]
引物(ovpd-1cds r):tcagaggggccaggagcagtgtcca(序列号68);
[0185]
引物(bupd-1cds f1):atggggaccccgcgggcgct(序列号69);
[0186]
引物(bupd-1cds r1):gatgaccaggctctgcatct(序列号70);
[0187]
引物(bupd-1cds f2):aatgacagcggcgtctactt(序列号71);
[0188]
引物(bupd-1cds r2):tcagaggggccaggagcagt(序列号72)。
[0189]
1.2.绵羊pd-1表达cos-7细胞的构建
[0190]
为了制作绵羊pd-1表达质粒,以所合成出的绵羊pbmc来源cdna为模板、使用向5
′
末端侧加入选择性酶bglii及smai识别部位而设计的引物(ovpd-1-egfp f及r)进行了pcr。对所得的pcr产物利用bglii及smai(takara公司)处理后,使用fastgene gel/pcr提取试剂盒(nippon genetics公司)纯化,导入进行了同样的选择性酶处理的pegfp-n2载体(clontech公司),进行了克隆。使用fastgene xpress plasmid plus kit(nippon genetics公司)提取表达质粒,在用于实验前以-30℃保存。以后,将所制作的表达质粒表示为pegfp-n2-ovpd-1。
[0191]
引物(ovpd-1-egfp f):
[0192]
gaagatctatggggaccccgcgggcgccg(序列号73);
[0193]
引物(ovpd-1-egfp r):
[0194]
gacccggggaggggccaggagcagtgtcc(序列号74)。
[0195]
将5
×
104/cm2的cos-7细胞在6孔板中进行传代,在包含10%钝化胎牛血清(invitrogen公司)、0.01%l-谷氨酸(life technologies公司)的rpmi 1640培养基中在37℃、5%co2存在下培养一晚。将pegfp-n2-ovpd-1或作为阴性对照的pegfp-n20.4μg/cm2使用lipofectamine 2000(invitrogen公司)导入cos-7细胞并培养48小时(ovpd-1-egfp表达细胞)。为了确认所制作出的表达细胞中的绵羊pd-1的表达,利用全功能荧光显微镜bz-9000(keyence公司)使egfp的细胞内局部可视化。
[0196]
1.3.大鼠抗牛pd-1抗体5d2与绵羊pd-1的反应性(图9)
[0197]
利用流式细胞仪法确认到大鼠抗牛pd-1单克隆抗体与绵羊pd-1发生交叉反应。将绵羊pd-1-egfp表达cos-7细胞使用加入有10%钝化山羊血清(invitrogen公司)的pbs在室温下封闭15分钟,使10μg/ml的大鼠抗牛pd-1抗体5d2在室温下反应30分钟,清洗后使apc标记抗大鼠ig山羊抗体(beckman coulter公司)在室温下反应30分钟。作为阴性对照抗体,使用了大鼠igg2a(κ)同种型对照(bd biosciences公司)。在分析时使用了facs verse(bd biosciences公司)。需要说明的是,在所有的清洗操作及抗体的稀释中,使用了加入有1%牛血清白蛋白(sigma-aldrich公司)的pbs。
[0198]
将实验结果以图9表示。确认了大鼠抗牛pd-1抗体5d2与绵羊pd-1表达细胞结合。
[0199]
1.4.大鼠抗牛pd-1抗体5d2与水牛的淋巴细胞的反应性(图10)
[0200]
从水牛(bubalus bubalis;亚洲水牛)的外周血中,利用使用了percoll(ge healthcare公司)的密度梯度离心法分离出外周血单核细胞(pbmc)。将分离出的水牛pbmc悬浮于包含10%钝化胎牛血清(cell culture technologies公司)、青霉素200u/ml、链霉素200μg/ml、0.01%l-谷氨酸(life technologies公司)的rpmi 1640培养基(sigma-aldrich公司)中,调整为2
×
106个/ml。向该pbmc中添加醋酸佛波豆蔻酯(phorbol12-myristate acetate)(pma)20ng/ml及离子霉素(ionomycin)1μg/ml(sigma-aldrich公司),在37℃、5%co2条件下培养2天。回收所培养的pbmc,使用加入有10%钝化山羊血清(invitrogen公司)的pbs在室温下封闭15分钟,使大鼠抗牛pd-1抗体5d2、小鼠抗牛cd8抗体(38.65,abd serotec公司)在37℃反应30分钟。作为阴性对照抗体,使用了大鼠igg2a(κ)同种型对照(bd biosciences公司)。清洗后,使apc标记山羊抗大鼠ig抗体(beckman coulter公司)、pe标记山羊抗小鼠igg抗体(beckman coulter公司)在室温下反应30分钟。进一步清洗后,使alexa flour488标记小鼠抗牛cd4抗体(cc30,abd serotec公司)及pe/cy7标记抗牛igm小鼠抗体(il-a30,abd serotec公司)在室温下反应30分钟。作为抗体的标记使用了zenon mouse igg1标记试剂盒(life technologies公司)或lightning-link kits(innova biosciences公司)。在分析时使用了facs verse(bd biosciences公司)。需要说明的是,在所有的清洗操作及抗体的稀释中,使用了加入有10%钝化山羊血清(invitrogen公司)的pbs。
[0201]
将实验结果以图10表示。大鼠抗牛pd-1抗体5d2与利用pma/离子霉素刺激活化了的水牛的cd4
+
t细胞(igm-cd4
+
)及cd8
+
t细胞(igm-cd8
+
)强烈地结合。
[0202]
〔实施例3〕具有野生型或突变型牛igg1的大鼠-牛嵌合抗牛pd-1抗体与牛fcγ受
igg1 adcc-,进行5~7天振荡培养后回收培养上清液。从培养上清液中使用ab capcherextra(protenova公司)纯化各嵌合抗体。与树脂的结合使用开管柱法,作为平衡化缓冲液及清洗缓冲液使用1.5m甘氨酸/3m nacl(ph 8.0)。作为洗脱缓冲液使用0.1m甘氨酸-hcl(ph 2.8),作为中和缓冲液使用1m tris(ph 9.0)。对纯化了的抗体使用pd-10脱盐柱(ge healthcare公司)及amicon ultra-15(50kda、millipore公司)进行了置换为pbs(ph 7.4)的缓冲液置换及浓缩。将纯化了的嵌合抗体通过0.22μm注射器过滤器(pall life sciences公司)而进行灭菌,在用于实验前以4℃保存。对纯化后的各嵌合抗体浓度利用使用nanodrop8000分光光度计(thermo fisher scientific公司)测定出的吸光度(280nm)进行了定量。
[0215]
2.3.大鼠-牛嵌合抗牛pd-1抗体的纯化纯度的确认(图13)
[0216]
为了确认纯化了的大鼠-牛嵌合抗牛pd-1抗体(ch5d2 igg1 wt及ch5d2 igg1 adcc-)的纯度,利用sds-page及cbb染色进行了抗体蛋白质的检测。将纯化了的各嵌合抗体悬浮于laemmli样品缓冲液(bio-rad公司)中,在还原条件下(利用2-巯基乙醇(sigma-aldrich公司)还原)或非还原条件下进行变性处理(95℃、5分钟)。对所制备的样品使用supersep ace 5%-20%梯度聚丙烯酰胺凝胶(和光纯药工业公司)进行电泳。此时,作为分子量标记使用了precision plus protein全蓝蛋白质标准品(bio-rad公司)。电泳后,利用quick-cbb(和光纯药工业公司)进行凝胶的染色,接下来在蒸馏水中进行脱色。
[0217]
将结果表示于图13中。还原条件下确认到25kda(轻链)及50kda(重链),非还原条件下在150kda的预估的位置确认到ch5d2 igg1 wt及ch5d2 igg1 adcc-的条带。
[0218]
2.4.可溶性牛fcγ受体(fcγr)的构建
[0219]
依照以下的步骤,构建了牛fcγri-his、fcγrii-his、fcγriii-his及fcγ2r-his表达质粒。以扩增牛fcγri、fcγrii、fcγriii及fcγ2r(genbank登记号nm_174538、nm_174539、nm_001077402及nm_001001138)的信号肽及细胞外区域的方式,设计了向5
′
末端侧加入有选择性酶noti及xhoi识别部位的引物(bofcγri-his f及r、bofcγriii-his f及r、或bofcγ2r-his f及r)或向5
′
末端侧加入有选择性酶nhei及ecorv识别部位的引物(bofcγriii-his f及r)。需要说明的是,向反向引物中加入了编码6
×
组氨酸(his)标签的基因序列。以所合成出的牛pbmc来源cdna为模板进行pcr,将各自的pcr产物利用noti(takara公司)及xhoi(takara公司)(fcγri-his、fcγriii-his及fcγ2r-his)或nhei(takara公司)及ecorv(takara公司)(fcγrii-his)处理后,使用fastgene gel/pcr提取试剂盒(nippon genetics公司)纯化,导入进行了同样的选择性酶处理的pcxn2.1(+)载体(niwa h,yamamura k,miyazaki j.gene,108(2):193-199;dec.15,1991;由顺天堂大学大学院医学研究科横沟岳彦教授分与),进行了克隆。利用fastgene xpress plasmid plus kit(nippon genetics公司)纯化表达质粒,在用于实验前以-30℃保存。以后,将所制作出的表达质粒表示为pcxn2.1-bofcγri-his、pcxn2.1-bofcγrii-his、pcxn2.1-bofcγriii-his或pcxn2.1-bofcγ2r-his。
[0220]
引物(bofcγri-his f):
[0221]
ataagaatgcggccgccaccatgtggctcataatagctct(序列号79);
[0222]
引物(bofcγri-his r):
[0223]
gccctcgagttaatggtgatggtgatggtgaggagttgttgactggaggc(序列号80);
[0224]
引物(bofcγrii-his f):
[0225]
ataagaatgctagccaccatggggatcccctcattcct(序列号81);
[0226]
引物(bofcγrii-his r):
[0227]
gccgatatcttaatggtgatggtgatggtgcgatgaggggccgctcgagc(序列号82);
[0228]
引物(bofcγriii-his f):
[0229]
ataagaatgcggccgccaccatgtg gcaactgctaccacc(序列号83);
[0230]
引物(bofcγriii-his r):
[0231]
gccctcgagttaatggtgatggtgatggtggtgccaaggtagaaagaatg(序列号84);
[0232]
引物(bofcγ2r-his f):
[0233]
ataagaatgcggccgccaccatggcccccaccctccctgccttgctct(序列号85);
[0234]
引物(bofcγ2r-his r):
[0235]
gccctcgagttaatggtgatggtgatggtgattctgcatcgtgtagtctg(序列号86)。
[0236]
依照以下的步骤,制作出可溶性牛fcγri-his、fcγrii-his、fcγriii-his及fcγ2r-his表达细胞。向7.5
×
107个的expi293f细胞(life technologies公司)中使用expifectamine(life technologies公司)导入30μg的pcxn2.1-bofcγri-his、pcxn2.1-bofcγrii-his、pcxn2.1-bofcγriii-his或pcxn2.1-bofcγ2r-his,进行5~7天振荡培养后回收培养上清液。从培养上清液中使用talon金属亲和树脂(clontech公司)纯化重组蛋白。纯化后,利用amicon ultra-15离心超滤装置(10kda、millipore公司)将缓冲液置换为pbs(ph 7.4),在用于实验前以-30℃保存(牛pd-1-his)。对纯化后的牛fcγri-his、fcγrii-his、fcγriii-his及fcγ2r-his浓度利用使用nanodrop8000分光光度计(thermo fisher scientific公司)测定出的吸光度(280nm)进行了定量。
[0237]
2.5.大鼠-牛嵌合抗牛pd-1抗体ch5d2 igg1 wt及igg1 adcc-与牛fcγr的结合性(图14)
[0238]
将大鼠-牛嵌合抗牛pd-1抗体ch5d2 igg1 wt或igg1 adcc-以终浓度50、25、12.5、6.25、3.12、1.5610nm在nunc maxisorp elisa板(nunc公司)中在37℃固相化2小时。其后,将各孔用加入有0.05%tween20的pbs(pbs-t)200μl清洗5次,使用superblock(pbs)封闭缓冲液(thermo fisher scientific公司)在37℃进行30分钟封闭。再次同样地进行清洗,将牛fcγri-his、fcγrii-his、fcγriii-his或fcγ2r-his以终浓度10μg/ml添加到各孔中,在37℃反应1小时。清洗后,使抗多聚组氨酸标签小鼠单克隆抗体(abcam公司)在37℃反应30分钟。然后,清洗各孔,使辣根过氧化物酶标记抗小鼠igg山羊多克隆抗体(mp biomedicals公司)在37℃反应30分钟。再次清洗各孔,加入tmb单组分底物(bethyl公司),进行了显色。其后,用0.18m稀硫酸停止酶反应,使用微板读数仪mtp-900(corona电气公司)测定出吸光度(450nm)。在板的各清洗操作中,使用了全自动洗板机biowasher 50(ds pharma biomedical公司)。
[0239]
将实验结果表示于图14中。igg1 wt与牛fcγri-his强结合,与牛fcγrii-his弱结合。另一方面,igg1 adcc-未与牛fcγri-his及fcγrii-his结合。另外,igg1 wt及igg1 adcc-均未观察到与牛fcγriii-his及牛fcγ2r-his的结合。
[0240]
将本说明书中引用的所有出版物、专利及专利申请原样不变地作为参考纳入本说明书中。
[0241]
产业上的可利用性
[0242]
本发明的抗pd-1抗体可以用于动物的癌症、感染症的预防和/或治疗。
[0243]
《序列号1》
[0244]
序列号1表示大鼠抗牛pd-1抗体的l链可变区的氨基酸序列。下划线部:从nh2末端起依次为cdr1、cdr2、cdr3。
[0245][0246]
《序列号2》
[0247]
序列号2表示大鼠抗牛pd-1抗体的h链可变区的氨基酸序列。下划线部:从nh2末端起依次为cdr1、cdr2、cdr3。
[0248][0249]
《序列号3》
[0250]
序列号3表示牛抗体的l链恒定区(牛ig lambda,genbank:x62917)的氨基酸序列。
[0251][0252]
《序列号4》
[0253]
序列号4表示牛抗体的h链恒定区(牛igg1,改变genbank:x62916)的氨基酸序列。在突变部位划有下划线。氨基酸序号及突变:123e
→
p,124l
→
v,125p
→
a,126g
→
缺失,218a
→
s,219p
→s[0254][0255]
《序列号5》
[0256]
序列号5表示大鼠抗牛pd-1抗体的l链可变区的核苷酸序列。
[0257][0258]
将序列号5的核苷酸序列的密码子最佳化后核苷酸序列表示于《序列号11》中。
[0259][0260]
《序列号6》
[0261]
序列号6表示大鼠抗牛pd-1抗体的h链可变区的核苷酸序列。
[0262][0263][0264]
将序列号6的核苷酸序列的密码子最佳化后核苷酸序列表示于《序列号12》中。
[0265][0266]
《序列号7》
[0267]
序列号7表示牛抗体的l链恒定区(牛ig lambda,genbank:x62917)的核苷酸序列。
[0268][0269]
将序列号7的核苷酸序列的密码子最佳化后核苷酸序列表示于《序列号13》中。
[0270][0271][0272]
《序列号8》
[0273]
序列号8表示牛抗体的h链恒定区(牛igg1,改变genbank:x62916)的核苷酸序列(密码子最佳化后)。
[0274][0275][0276]
《序列号9》
[0277]
序列号9表示包含大鼠抗牛pd-1抗体的l链可变区和牛抗体的l链恒定区的嵌合l链的氨基酸序列。
[0278][0279]
《序列号10》
[0280]
序列号10表示包含大鼠抗牛pd-1抗体的h链可变区和牛抗体的h链恒定区(牛igg1,改变genbank:x62916)的嵌合h链的氨基酸序列。
[0281][0282]
《序列号14》
[0283]
表示包含大鼠抗牛pd-1抗体的l链可变区和牛抗体的l链恒定区的嵌合l链的核苷酸序列(密码子最佳化后核苷酸序列)。
[0284][0285][0286]
《序列号15》
[0287]
表示包含大鼠抗牛pd-1抗体的h链可变区和牛抗体的h链恒定区(牛igg1,改变genbank:x62916)的嵌合h链的核苷酸序列(密码子最佳化后核苷酸序列)。
[0288][0289][0290]
《序列号16》
[0291]
序列号16表示大鼠抗牛pd-1抗体5d2的l链可变区的cdr1的氨基酸序列
(qsleysdgyty)。
[0292]
《序列号17》
[0293]
序列号17表示大鼠抗牛pd-1抗体5d2的l链可变区的cdr3的氨基酸序列(fqathdpdt)。
[0294]
《序列号18》
[0295]
序列号18表示大鼠抗牛pd-1抗体5d2的h链可变区的cdr1的氨基酸序列(gfsltsyy)。
[0296]
《序列号19》
[0297]
序列号19表示大鼠抗牛pd-1抗体5d2的h链可变区的cdr2的氨基酸序列(irsggst)。
[0298]
《序列号20》
[0299]
序列号20表示大鼠抗牛pd-1抗体5d2的h链可变区的cdr3的氨基酸序列(artssgyeggfdy)。
[0300]
《序列号21》
[0301]
序列号21表示牛抗体(igg1突变体1)的h链恒定区(ch1~ch3)的氨基酸序列。
[0302]
《序列号22》
[0303]
序列号22表示牛抗体(igg1突变体2)的h链恒定区(ch1~ch3)的氨基酸序列。
[0304]
《序列号23》
[0305]
序列号23表示牛抗体(igg1突变体3)的h链恒定区(ch1~ch3)的氨基酸序列。
[0306]
《序列号24》
[0307]
序列号24表示牛抗体(igg2突变体1)的h链恒定区(ch1~ch3)的氨基酸序列。
[0308]
《序列号25》
[0309]
序列号25表示牛抗体(igg2突变体2)的h链恒定区(ch1~ch3)的氨基酸序列。
[0310]
《序列号26》
[0311]
序列号26表示牛抗体(igg2突变体3)的h链恒定区(ch1~ch3)的氨基酸序列。
[0312]
《序列号27》
[0313]
序列号27表示牛抗体(igg3突变体1)的h链恒定区(ch1~ch3)的氨基酸序列。
[0314]
《序列号28》
[0315]
序列号28表示牛抗体(igg3突变体2)的h链恒定区(ch1~ch3)的氨基酸序列。
[0316]
《序列号29》
[0317]
序列号29表示牛抗体(igg1突变体1)的h链恒定区(ch1~ch3)的核苷酸序列。
[0318]
《序列号30》
[0319]
序列号30表示牛抗体(igg1突变体2)的h链恒定区(ch1~ch3)的核苷酸序列。
[0320]
《序列号31》
[0321]
序列号31表示牛抗体(igg1突变体3)的h链恒定区(ch1~ch3)的核苷酸序列。
[0322]
《序列号32》
[0323]
序列号32表示牛抗体(igg2突变体1)的h链恒定区(ch1~ch3)的核苷酸序列。
[0324]
《序列号33》
[0325]
序列号33表示牛抗体(igg2突变体2)的h链恒定区(ch1~ch3)的核苷酸序列。
[0326]
《序列号34》
[0327]
序列号34表示牛抗体(igg2突变体3)的h链恒定区(ch1~ch3)的核苷酸序列。
[0328]
《序列号35》
[0329]
序列号35表示牛抗体(igg3突变体1)的h链恒定区(ch1~ch3)的核苷酸序列。
[0330]
《序列号36》
[0331]
序列号36表示牛抗体(igg3突变体2)的h链恒定区(ch1~ch3)的核苷酸序列。
[0332]
《序列号37》
[0333]
序列号37表示绵羊抗体(igg1)的h链恒定区(ch1~ch3)的氨基酸序列。
[0334]
《序列号38》
[0335]
序列号38表示绵羊抗体(igg1)的h链恒定区(ch1~ch3)的核苷酸序列。
[0336]
《序列号39》
[0337]
序列号39表示绵羊抗体(igg2)的h链恒定区(ch1~ch3)的氨基酸序列。
[0338]
《序列号40》
[0339]
序列号40表示绵羊抗体(igg2)的h链恒定区(ch1~ch3)的核苷酸序列。
[0340]
《序列号41》
[0341]
序列号41表示绵羊抗体的l链(igkappa(ck))恒定区的氨基酸序列。
[0342]
《序列号42》
[0343]
序列号42表示绵羊抗体的l链(igkappa(ck))恒定区的核苷酸序列。
[0344]
《序列号43》
[0345]
序列号43表示绵羊抗体的l链(iglambda(cl))恒定区的氨基酸序列。
[0346]
《序列号44》
[0347]
序列号44表示绵羊抗体的l链(iglambda(cl))恒定区的核苷酸序列。
[0348]
《序列号45》
[0349]
序列号45表示水牛抗体(推定为igg1)的h链恒定区(ch1~ch3)的氨基酸序列。
[0350]
《序列号46》
[0351]
序列号46表示水牛抗体(推定为igg1)的h链恒定区(ch1~ch3)的核苷酸序列。
[0352]
《序列号47》
[0353]
序列号47表示水牛抗体(推定为igg2)的h链恒定区(ch1~ch3)的氨基酸序列。
[0354]
《序列号48》
[0355]
序列号48表示水牛抗体(推定为igg2)的h链恒定区(ch1~ch3)的核苷酸序列。
[0356]
《序列号49》
[0357]
序列号49表示水牛抗体(推定为igg3)的h链恒定区(ch1~ch3)的氨基酸序列。
[0358]
《序列号50》
[0359]
序列号50表示水牛抗体(推定为igg3)的h链恒定区(ch1~ch3)的核苷酸序列。
[0360]
《序列号51》
[0361]
序列号51表示水牛抗体的l链(推定为iglambda)恒定区(cl)的氨基酸序列。
[0362]
《序列号52》
[0363]
序列号52表示水牛抗体的l链(推定为iglambda)恒定区(cl)的核苷酸序列。
[0364]
《序列号53》
[0365]
序列号53表示人抗体(igg4突变体1)的h链恒定区(ch1~ch3)的氨基酸序列。
[0366]
《序列号54》
[0367]
序列号54表示人抗体(igg4突变体1)的h链恒定区(ch1~ch3)的核苷酸序列。
[0368]
《序列号55》
[0369]
序列号55表示人抗体(igg4突变体2)的h链恒定区(ch1~ch3)的氨基酸序列。
[0370]
《序列号56》
[0371]
序列号56表示人抗体(igg4突变体2)的h链恒定区(ch1~ch3)的核苷酸序列。
[0372]
《序列号57》
[0373]
序列号57表示人抗体(igg4突变体3)的h链恒定区(ch1~ch3)的氨基酸序列。
[0374]
《序列号58》
[0375]
序列号58表示人抗体(igg4突变体3)的h链恒定区(ch1~ch3)的核苷酸序列。
[0376]
《序列号59》
[0377]
序列号59表示人抗体的l链恒定区的氨基酸序列。
[0378]
《序列号60》
[0379]
序列号60表示人抗体的l链恒定区的核苷酸序列。
[0380]
《序列号61~74》
[0381]
序列号61~74依次表示引物bopd-1-myc f、bopd-1-myc r、bopd-l1-egfp f、bopd-l1-egfp r、bopd-1-his f、bopd-1-his r、ovpd-1cds f、ovpd-1cds r、bupd-1cds f1、bupd-1cds r1、bupd-1cds f2、bupd-1cds r2、ovpd-1-egfp f及ovpd-1-egfp r的核苷酸序列。
[0382]
《序列号75》
[0383]
序列号75表示包含大鼠抗牛pd-1抗体的h链可变区和牛抗体的h链恒定区(牛igg1,genbank:x62916)的嵌合h链的核苷酸序列。
[0384][0385][0386]
《序列号76》
[0387]
序列号75表示包含大鼠抗牛pd-1抗体的h链可变区和牛抗体的h链恒定区(牛igg1,genbank:x62916)的嵌合h链的氨基酸序列。
[0388][0389]
《序列号77~86》
[0390]
序列号77~86依次表示引物boigg1 ch1 f、boigg1 ch3 r、bofcγri-his f、bofcγri-his r、bofcγrii-his f、bofcγrii-his r、bofcγriii-his f、bofcγriii-his r、bofcγ2r-his f及bofcγ2r-his r的核苷酸序列。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1