本发明涉及一种放线菌代谢动态调控方法及其应用,涉及分子生物学。
背景技术:
1、次级代谢产物是指微生物生长到一定阶段后通过次级代谢合成的分子结构十分复杂、对该生物无明显生理功能,或并非是该生物生长和繁殖所必需的小分子物质,如抗生素、毒素、激素、色素等。通过分子生物学手段,调控微生物体内关键酶的合成,是提高次级代谢产物产量的关键。传统的静态调控策略,包括基因敲除、过表达等,往往会导致细胞生长与产物合成之间的代谢流不均衡,影响产物的合成效率。而动态调控策略可以有效克服或弱化以上缺陷,其能够实时响应细胞环境变化,在合适时间点对代谢网络进行干预,重塑产物合成与细胞生长之间的代谢流分配,提高产物的合成能力。目前,动态调控策略主要包括以下三种:
2、1)基于外源添加诱导剂的动态调控系统
3、通过外源诱导剂的添加可以在任何恰当的时间点方便的控制目的基因的表达,还可以通过调整诱导剂的添加量来控制目的基因的表达强度。总体上来看,通过此系统可以达到减小产物以及副产物对细胞的毒害作用,提高生物量,进一步提高产率和产量。缺点是需要外源添加诱导剂,而且需要人为的去摸索和检测细胞所处的生长状态,需要花费一定的人力和财力成本。
4、2)途径依赖型动态调控系统
5、途径依赖型动态调控系统一般都需要一个能够响应目标产物或者中间代谢物的效应蛋白,该蛋白接受特定信号后才能发挥作用,去打开或者关闭某些代谢通路。途径依赖型动态调控系统不需要人为的添加诱导剂,通过自动响应特定的小分子能够灵敏的开启或者是关闭某些代谢通路,使代谢流更多的流向目的产物的合成,以提高产率或产量。但是这类动态调控系统的效应蛋白往往仅能识别一种特异的小分子(中间代谢物或者终产物),底物识别谱非常狭窄,很难应用于其他类型的重要产物。到目前为止,此类系统的研究相对有限,主要在大肠杆菌和酿酒酵母等模式工业菌株中开展,也仅在包括脂肪酸、谷氨酸、lycopene和biodiesel等几种产物中应用,大大限制了此类系统的广泛应用。
6、3)途径非依赖型动态调控系统
7、为了解决以上问题,目前还开发了途径非依赖型动态调控系统。这类动态调控系统的使用不再限定于某一种或者某一类特定的小分子,也不需要外源添加诱导剂和人为的监测细胞的生长状态,能够全自动的进行动态调控,理论上能够普遍适用。目前,研究得最为清楚、使用最为广泛的是基于quorum-sensing(qs)系统的动态调控。
8、qs系统是指细菌通过自诱导分子(ai,autoinducer),根据群体密度的变化调节基因表达的机制,用来进行群体间的交流以协调群体行为。这类自诱导分子一般是细菌自身合成并分泌到胞外的信号分子,包括n-(3-oxohexanoyl)-homoserine lactone(高丝氨酸内酯,ahl)、γ-butyrolactone(γ-丁内酯,gbl)等。自诱导分子的浓度随着菌体密度的升高而升高,达到一定阈值时启动qs系统,调控下游基因的表达,参与多种细胞生命活动,比如群集运动,致病性,抗生素的合成等。qs系统最早的详细研究来源于vibrio fischeri(费氏弧菌),其中包括关键蛋白luxr、luxi和luxr特异性识别的启动子pluxi。luxi负责合成信号分子ahl,ahl能够与调控蛋白luxr结合。在ahl浓度达到一定水平的情况下luxr可以激活启动子pluxi下游基因的表达。在ahl浓度水平处在阈值之下时,luxr不能激活启动子pluxi下游基因的表达。qs系统的优点在于此系统跟细胞密度密切相关,当细胞密度处于较低水平时该系统不会被激活,当细胞密度达到一定阈值时系统才会被激活。该系统中使用的小分子(比如ahl)能够自由的透过细胞膜,从而使整个细胞群体的状态高度同步。该系统也研究的较为充分,应用也比较广泛。但是,通过群体效应系统发挥作用的细胞密度阈值和胞内目标次级代谢产物的合成通常并不同步,限制了途径非依赖型动态调控系统的应用。
9、基于上述三种方法的特点,本发明提供一种新的放线菌代谢动态调控方法,用于在合适时间点对放线菌的代谢网络进行干预,重塑产物合成与细胞生长之间的代谢流分配,提高目标化合物的合成能力。
技术实现思路
1、本发明提供一种放线菌代谢动态调控方法及其应用,用于在合适时间点对放线菌的代谢网络进行干预,重塑产物合成与细胞生长之间的代谢流分配,提高目标化合物的合成能力。
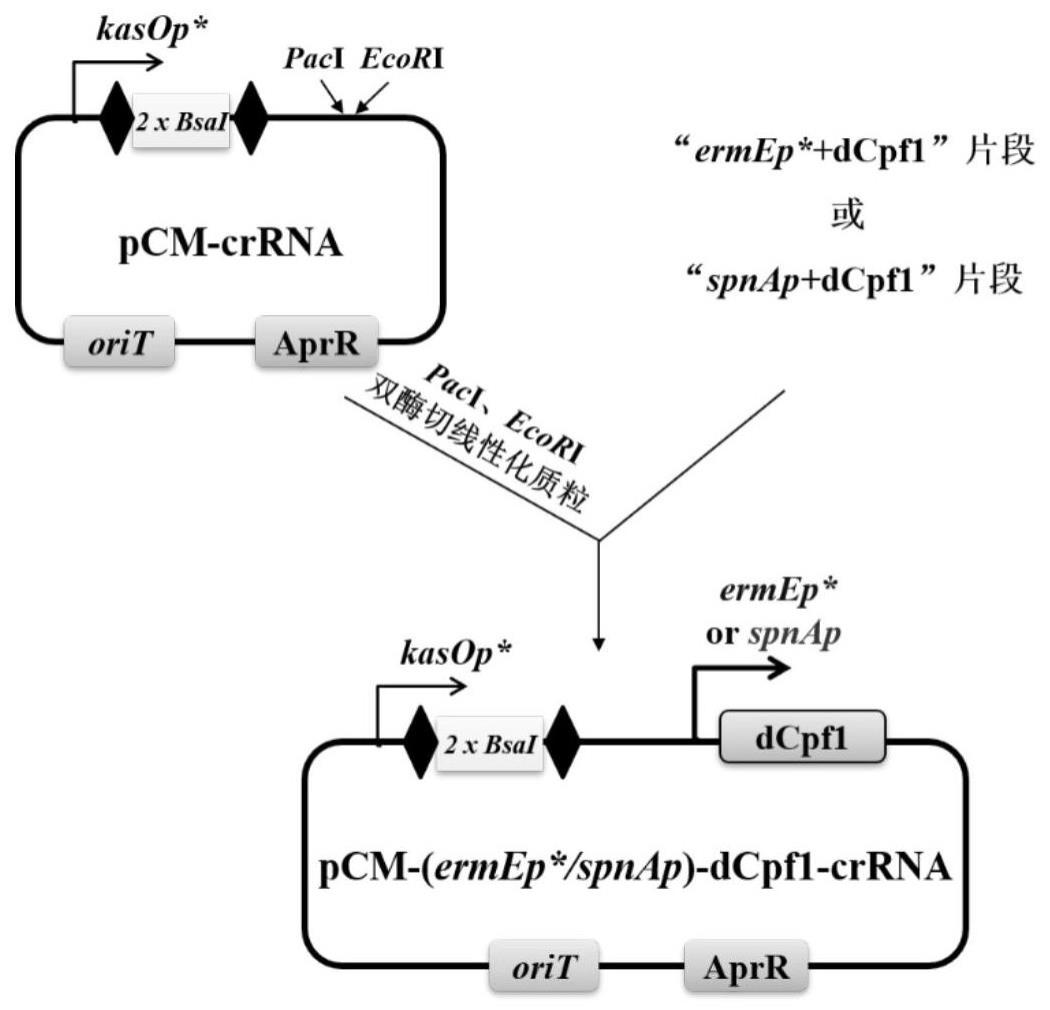
2、本发明第一方面提供一种重组质粒,所述重组质粒用于抑制微生物基因组内至少一组第一目标基因的表达,提高微生物次级代谢产物中目标化合物的产量,所述重组质粒包括定位单元和调控单元,其中:
3、所述定位单元包括第一启动子、位于所述第一启动子下游的靶向第一目标基因的crrna序列、以及位于所述crrna序列上下游的repeat序列;
4、所述调控单元包括第二启动子以及位于所述第二启动子下游的能够编码丧失切割活性的cas蛋白的基因序列,所述第二启动子具有时序性表达规律,并且,所述第二启动子在所述微生物体内,用于启动第二目标基因的表达,所述第二目标基因能够表达得到合成所述目标化合物的酶。
5、进一步地,所述第一启动子的核苷酸序列如seq id no:1所示。
6、进一步地,所述repeat序列的核苷酸序列如seq id no:2所示,所述丧失切割活性的cas蛋白的氨基酸序列如seq id no:3所示。
7、进一步地,所述重组质粒还包括接合转移单元、筛选单元中的一种或两种。
8、本发明第二方面提供上述任一所述重组质粒的构建方法,包括如下步骤:
9、确定所述定位单元和调控单元的基因序列;
10、将所述定位单元构建到出发质粒上,得到第一重组质粒;
11、对所述第一重组质粒进行酶切,将得到的线性化酶切产物与所述调控单元连接,得到所述重组质粒。
12、进一步地,所述出发质粒为整合型质粒pcm265,并且所述整合型质粒pcm265上不包括ⅱ-s型内切酶bsai切割位点。
13、本发明第三方面提供上述任一所述重组质粒在提高放线菌次级代谢产物中目标化合物产量中的应用。
14、本发明第四方面提供一种提高放线菌次级代谢产物中目标化合物产量的方法,包括如下步骤:
15、确定所述第一目标基因和第二目标基因;
16、根据所述第一目标基因设计crrna序列,根据所述第二目标基因确定所述第二启动子,并根据所述crrna序列和第二启动子构建得到如上述任一所述的重组质粒;
17、将所述重组质粒导入放线菌中,筛选阳性转化子,对所述阳性转化子进行培养,收集目标化合物。
18、进一步地,所述放线菌为刺糖多孢菌,所述第一目标基因为glta1、glta2、glta3、glta4、fabh1、fabh2、fabh3、fabh4、glgc中的一种或多种,所述第二启动子的核苷酸序列如seq id no:4-5中的一种所示。
19、进一步地,所述放线菌为天蓝色链霉菌,所述第一目标基因为柠檬酸合成酶基因glta,所述第二启动子的核苷酸序列如seq id no:6所示。
20、本发明利用具有时序性表达规律的启动子驱动crispri,对影响目标化合物合成的第一目标基因的表达进行抑制,无需外源诱导剂的添加,无需挖掘可感知中间代谢产物的生物传感器,在不影响微生物生长和初级代谢的情况下,使细胞代谢流向目标化合物的合成,实现目标化合物的动态调控;此外,本发明提供的方法对具有通用性,有望成为放线菌合成代谢工程改造的一种通用策略。