一种促进长日照条件下大豆开花时间提前的方法
本发明属于大豆分子遗传育种领域,具体涉及一种促进长日照条件下大豆开花时间提前的方法。
背景技术:
1、开花标志着植物由营养生长向生殖生长的转变,这一过程受复杂的遗传网络和环境条件所调控。已有研究表明,在模式植物拟南芥中有6条成花转变途径,分别是光周期途径、自主调节途径、春化途径、苗龄依赖途径、赤霉素途径和环境温度影响途径。大豆是典型的短日照作物,光周期是影响大豆开花和成熟以及生态适应性的重要因素。近年来,在大豆光周期调控开花领域取得了许多突破性进展,光周期调控大豆开花的遗传网路逐步建立。
2、在大豆光周期开花途径中,大豆e3(gmphya3)和e4(gmphya2)是拟南芥光敏色素phytochromea的同源基因,起抑制开花的作用。在长日照下,e3突变体比对照早开花;在远红光条件下,e4突变体下胚轴比对照长。e1是控制大豆成熟期的位点,e1突变体在自然条件下具有早花表型;e1蛋白是一个含有b3结构域的核蛋白,通过抑制floweringlocus t同源基因(gmft2a和gmft5a)的表达而抑制大豆开花。e2是拟南芥gigantea的同源基因,e2突变体中gmft2a表达量增加导致早花。过量表达pseudoresponseregulator同源基因(gmprr3a和gmprr3b)通过late elongated hypocotyl(lhy)促进e1的表达导致大豆晚花。constans的同源基因gmcol1a和gmcol1b在长日照下抑制大豆开花。过量表达gmft1a和gmft4抑制大豆开花,在短日照下,过量表达gmft1a的植株比野生型晚4-7天开花,而在长日照下,过量表达gmft1a的植株比野生型晚3-5天开花。在长日和短日照下,过量表达gmrav均抑制大豆开花。过量表达gmspa3c抑制大豆开花,gmspa3c功能丧失的突变体在长日照下具有早花表明,而短日照下开花时间没有显著差异。
3、除了上述几个开花抑制基因之外,大豆光周期途径中也发现一些开花促进基因。过量表达大豆gmft2a导致早花,而gmft2a突变体表现出晚花。过量表达大豆gmft5a在长日照下促进开花。大豆长童期基因j(earlyflowering 3同源基因)是生物钟夜间复合体重要组成成分,通过抑制e1的表达促进开花,j突变体晚开花,适合在低纬度种植。大豆gmgbp1是一个开花促进因子,gmgbp1表达下调的转基因株系在短日、长日和自然条件下比野生型晚开花。luxarrythmo是生物钟夜间复合体另一重要组成成分,它与j相互作用,抑制e1的表达;大豆lux1lux2双突变体表现出极度晚花。time offlowering 16(tof16)/lhy是控制大豆成熟期的位点,也是一个生物钟基因,在低纬度下,tof16cr大豆突变体早开花。大豆gmful(fruitful)通过促进gmft2a和gmft5a的表达促进开花。大豆soc1a/b促进开花,soc1a单突变体、soc1b单突变体和soc1asoc1b双突变体在长日和短日照下均具有晚花表型。过量表达大豆gmmde06通过促进gmft2a和gmft5a的表达促进开花。最近的研究表明大豆qne1(qtlnear e1)是tcp类型的转录因子;过量表达大豆qne1使大豆早开花。
4、由此可见,大豆光周期途径的一些重要开花基因的功能已初步确定,但还有很多基因的功能仍未知,其具体的调控机制有待于进一步探索。
技术实现思路
1、本发明的目的为促进长日照条件下大豆开花时间提前。
2、本发明首先保护一种融合蛋白gfp-gmcdf1-c17,其从n端至c端依次由gfp蛋白、连接肽蛋白和gmcdf1-c17蛋白组成;
3、所述连接肽蛋白由9-13个氨基酸残基组成;
4、所述gmcdf1-c17蛋白为如下b1)或b2)或b3)或b4):
5、b1)氨基酸序列是seq id no:1自5’末端起第250至266位所示的蛋白质;
6、b2)在seq id no:1自5’末端起第250至266位所示的蛋白质的n端或/和c端连接标签得到的融合蛋白质;
7、b3)将b1)或b2)所示的蛋白质经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的、来源于大豆且与大豆开花时间相关的蛋白质;
8、b4)与seq id no:1自5’末端起第250至266位限定的氨基酸序列具有80%或80%以上同源性、来源于大豆且与大豆开花时间相关的蛋白质。
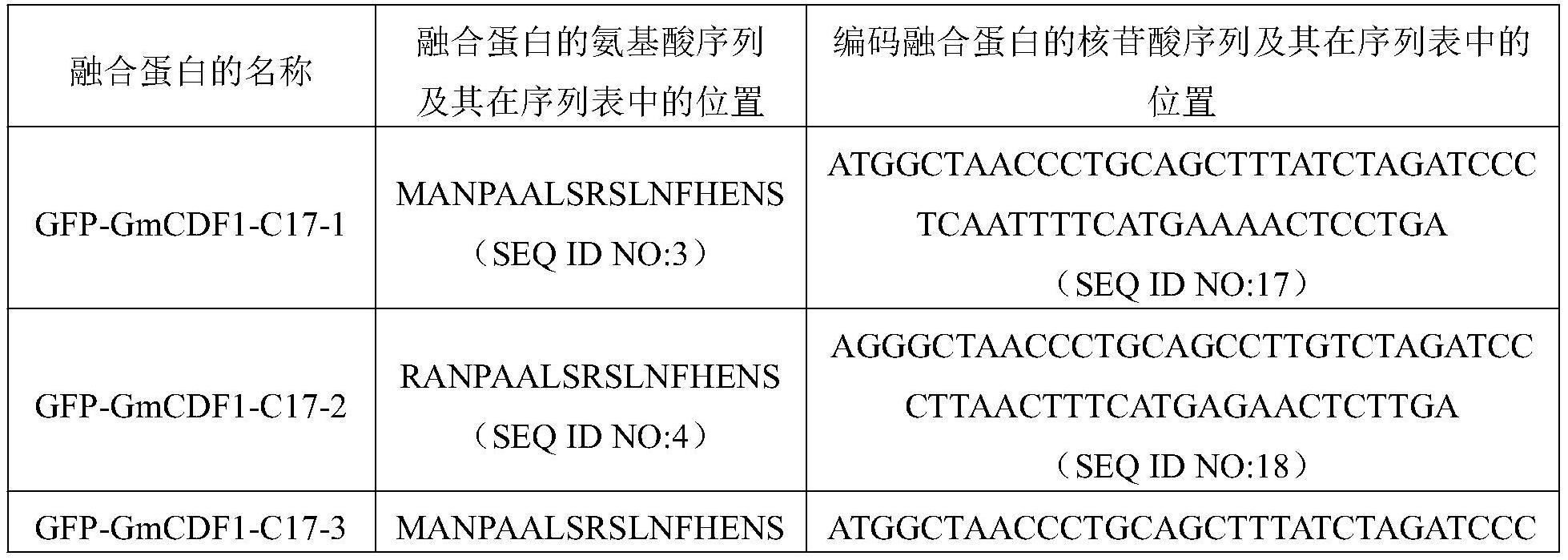
9、为了使b1)中的蛋白质便于纯化,可在seq id no:1自5’末端起第250至266位所示的蛋白质的氨基末端或羧基末端连接上如表1所示的标签。
10、表1.标签的序列
11、 标签 残基 序列 poly-arg 5-6(通常为5个) rrrrr flag 8 dykddddk strep-tagii 8 wshpqfek c-myc 10 eqkliseedl
12、上述b3)中的蛋白质,所述一个或几个氨基酸残基的取代和/或缺失和/或添加为不超过10个氨基酸残基的取代和/或缺失和/或添加。
13、上述b3)中的蛋白质可人工合成,也可先合成其编码基因,再进行生物表达得到。
14、上述b3)中的蛋白质的编码基因可通过将seq id no:2自5’末端起第748至798位所示的dna序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变,和/或在其5′端和/或3′端连上表1所示的标签的编码序列得到。
15、上述b4)中的蛋白质可为表2所示的融合蛋白
16、gfp-gmcdf1-c17-1—gfp-gmcdf1-c17-14,其氨基酸序列依次如seq id no:3—seq id no:16所示。编码表2所示的融合蛋白gfp-gmcdf1-c17-1—gfp-gmcdf1-c17-14的核苷酸序列依次如seq id no:17—seq id no:30所示。
17、表2
18、
19、
20、上述融合蛋白gfp-gmcdf1-c17中,所述gfp蛋白的氨基酸序列如seq id no:1自5’末端起第1至238位所示。
21、上述融合蛋白gfp-gmcdf1-c17中,所述连接肽蛋白的氨基酸序列如seq id no:1自5’末端起第239至249位所示。
22、所述融合蛋白gfp-gmcdf1-c17的氨基酸序列如seq id no:1所示。
23、编码上述任一所述融合蛋白gfp-gmcdf1-c17的核酸分子也属于本发明的保护范围。
24、所述核酸分子可为c1)或c2)或c3)或c4)所示的dna分子:
25、c1)编码区为seq id no:2所示的dna分子;
26、c2)核苷酸序列为seq id no:2所示的dna分子;
27、c3)与c1)或c2)限定的核苷酸序列具有75%或75%以上同源性,来源于大豆且编码上述任一所述融合蛋白gfp-gmcdf1-c17的dna分子;
28、c4)在严格条件下与c1)或c2)限定的核苷酸序列杂交,来源于大豆且编码上述任一所述融合蛋白gfp-gmcdf1-c17的dna分子。
29、其中,所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如mrna或hnrna等。
30、其中,seq id no:2由801个核苷酸组成,seq id no:2的核苷酸编码seq id no:1所示的氨基酸序列。
31、本领域普通技术人员可以很容易地采用已知的方法,例如定向进化和点突变的方法,对本发明的编码上述任一所述融合蛋白gfp-gmcdf1-c17的核苷酸序列进行突变。那些经过人工修饰的,具有与本发明分离得到的上述任一所述融合蛋白gfp-gmcdf1-c17的核苷酸序列80%或者更高同一性的核苷酸,只要编码上述任一所述融合蛋白gfp-gmcdf1-c17,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。
32、这里使用的术语“同一性”指与天然核酸序列的序列相似性。“同一性”包括与本发明的编码seq id no:1所示的氨基酸序列组成的融合蛋白gfp-gmcdf1-c17的核苷酸序列具有80%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或计算机软件进行评价。使用计算机软件,两个或多个序列之间的同一性可以用百分比(%)表示,其可以用来评价相关序列之间的同一性。
33、本发明还保护上述任一所述融合蛋白gfp-gmcdf1-c17在调控大豆开花时间或培育开花时间提前的转基因大豆中的应用。
34、本发明还保护上述任一所述核酸分子在调控大豆开花时间或培育开花时间提前的转基因大豆中的应用。
35、上述任一所述的应用中,所述调控大豆开花时间可为大豆开花时间提前。
36、上述任一所述的应用中,所述大豆的培养条件可为长日照(如(16h光照/8h黑暗)。
37、本发明还保护一种培育转基因大豆的方法,可包括如下步骤:提高出发大豆中上述任一所述融合蛋白gfp-gmcdf1-c17的表达量和/或活性,得到转基因大豆;与出发大豆相比,转基因大豆的开花时间提前。
38、上述方法中,所述提高出发大豆中上述任一所述融合蛋白gfp-gmcdf1-c17的表达量和/或活性可通过转基因、多拷贝、改变启动子、调控因子等本领域熟知的方法,达到提高出发大豆中上述任一所述融合蛋白gfp-gmcdf1-c17的表达量和/或活性的效果。
39、上述方法中,所述提高出发大豆中上述任一所述融合蛋白gfp-gmcdf1-c17的表达量和/或活性通过向出发大豆中导入编码上述任一所述融合蛋白gfp-gmcdf1-c17的核酸分子实现。
40、上述方法中,所述向出发大豆中导入编码上述任一所述融合蛋白gfp-gmcdf1-c17的核酸分子可通过向出发大豆中导入重组载体实现;所述重组载体可为向表达载体插入编码上述任一所述融合蛋白gfp-gmcdf1-c17的核酸分子,得到的重组质粒。所述重组载体具体可为实施例提及的重组质粒ptf101-gfp-c17。
41、上述方法中,所述编码上述任一所述融合蛋白gfp-gmcdf1-c17的核酸分子可为c1)或c2)或c3)或c4)所示的dna分子:
42、c1)编码区为seq id no:2所示的dna分子;
43、c2)核苷酸序列为seq id no:2所示的dna分子;
44、c3)与c1)或c2)限定的核苷酸序列具有75%或75%以上同源性,来源于大豆且编码上述任一所述融合蛋白gfp-gmcdf1-c17的dna分子;
45、c4)在严格条件下与c1)或c2)限定的核苷酸序列杂交,来源于大豆且编码上述任一所述融合蛋白gfp-gmcdf1-c17的dna分子。
46、其中,所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如mrna或hnrna等。
47、其中,seq id no:2由801个核苷酸组成,seq id no:2的核苷酸编码seq id no:1所示的氨基酸序列。
48、本发明还保护一种大豆育种方法,包括如下步骤:增加大豆中上述任一所述融合蛋白gfp-gmcdf1-c17的含量和/或活性,从而使大豆开花时间提前。
49、上述任一所述的方法中,所述大豆的培养条件可为长日照(如(16h光照/8h黑暗)。
50、实验证明,向大豆品种jack中导入重组质粒ptf101-gfp-c17,然后通过自交,获得t1代转gfp-gmcdf1-c17基因大豆。长日照条件下,与大豆品种jack相比,3个t1代转gfp-gmcdf1-c17基因大豆株系(gfp-c17-oe1、gfp-c17-oe2和gfp-c17-oe3)的开花时间均显著提前20天左右。由此可见,融合蛋白gfp-gmcdf1-c17可以调控大豆开花时间。本发明具有重要的应用价值。
- 还没有人留言评论。精彩留言会获得点赞!