一种植物温敏不育基因突变体tms11及其应用
本发明涉及基因学领域,具体涉及一种水稻温敏不育突变体tms11及其应用,其具有温敏不育特性,并且可以在特定条件下恢复育性。
背景技术:
1、水稻作为我国重要的粮食作物之一,全国有60%以上的人口以大米为食(庞乾林,2004)。利用杂种优势可以使水稻产量提高20%-50%(tester and langridge,2010)。水稻作为一种雌雄同花、自花授粉植物给水稻杂交育种造成了极大的麻烦,因此发掘雄性不育植株以及利用其生产水稻杂交种是水稻杂种优势利用的重点。
2、目前水稻杂交制种主要应用三系法和两系法。其中基于细胞质雄性不育(cms)的三系法包括:cms系、恢复系、保持系。cms系作为杂交母本,含有细胞质雄性不育基因,同时细胞核中缺少恢复基因(rf),表现为雄性不育。但是由于rf基因只在少数水稻品种中存在,因此限制了杂种优势的利用。环境敏感型细胞核雄性不育(egms)被认为是可以应用于杂交育种的(virmani and ilyas-ahmed,2001)。egms的育性会随着环境因素的变化而改变,由于egms是由细胞核基因控制,不再受到rf基因的限制,因此在两系法杂交育种中只需要egms系母本与任意一种有花粉的父本既可以实现配种,不仅节省了制种成本还可以更大程度的利用杂种优势,2018年我国杂交水稻推广面积中两系杂交稻已经达到了44%,且呈现逐年提高的态势(2019年中国水稻产业发展报告,2019)。目前我国的三系杂交种的推广面积仍然高于两系杂交种,要实现杂交水稻由三系向两系转变的战略部署,新的egms系的发掘与应用,以及相关egms系的育性转换机制的深入研究都至关重要。
3、第一个egms水稻品系是石明松先生1973年在粳稻中发现的光照敏感型核雄性不育系(pgms),表现为长日照下雄性不育,短日照下恢复育性(shi ms,1985)。随后在1988年发现的annong s-1是一个温度敏感型核雄性不育(tgms)突变体,导致该突变体表型的基因tms5由于其保守性较强,且完全敲除就会表现出良好且稳定的tgms表型而受到广泛应用(邓华凤等.,1999;jian-hang et al.,2003;zhou et al.,2014)。目前水稻两系育种中广泛应用的主要是光敏或温敏雄性不育,因此光温敏雄性不育基因的克隆以及相应育性转换的分子机制研究是至关重要的。
4、自两系法建立以来已有许多光温敏核不育基因相继被定位及克隆。农垦58s作为第一株被发现的pgms系,其pgms表型是由pms1 pms3两个位点决定的,只有当两个位点都是纯合时农垦58s才是长日照下完全不育的(mei,m.h et al.,1999)。这些phasirnas在农垦58s与野生型的积累再长日照下存在差异,研究者认为这些phasirnas在长日照下的过度积累是导致农垦58s不育的原因(ding,j et al.,2012;fan,y et al.,2016)。
5、而annong s-1是第一个在籼稻中被发现的水稻温敏雄性不育突变体,其tgms表型由单基因隐型基因tms5控制(jian-hang et al.,2003)。水稻tms5编码了一个核糖核酸酶z(rnase z)称之为rnasezs1,ubl40是保守的泛素-60s核糖体蛋白l40家族成员,这些mrna的表达水平对小孢子母细胞的正常发育至关重要,转录组分析表明rnasezs1在高温下影响了小孢子母细胞的ubl40的剪切,导致ubl40高量表达,导致小孢子败育(zhou et al.,2014)。然而高低温下如何导致的ubl40的表达量差异还未可知。2011年,用含tms5的tgms选育的71个商业双系杂交稻品种至少占中国所有两系杂交稻品种的71%,占中国所有两系杂交稻种植土地的83.8%(zhou et al.,2014)。目前为止tms5仍然是两系杂交育种的主要应用位点,对于tms5所导致的光温敏机制仍然有待进一步挖掘。
6、综上所述,目前虽然报道了一些光温敏不育位点,但是一方面光温敏效应的共性机制还未被揭示,另一方面在育种过程中,基因资源的多样性至关重要。模式植物中的光温敏研究为光温敏的共性机制提供了方向,研究者认为低温下植株特别是小孢子的发育缓慢是光温敏不育突变体恢复育性的关键(zhu,j et al.,2020;zhang,c et al.,2022)。早有研究表明减数分裂对温度是极度敏感的(dowrick,g.j,1957;de jaeger-braet,j et al.,2022;endo,m et al.,2009;zinn,k.e et al.,2010),减数分裂作为植物有性生殖的核心过程,其相关基因具有较高的特异性,并且在许多作物中极其保守。
7、因此,找到更多具有植物温敏不育特性的基因资源对于水稻育种而言,具有重要意义。
技术实现思路
1、针对上述问题,本技术的发明人尝试不同的方式进行温敏不育基因的研究,并且意外获得了一种温敏不育突变体tms11。
2、具体而言,为了获得更多的新的光温敏不育位点,深入研究水稻光温敏不育的分子机制,本技术的发明人对于各种水稻品种的种子进行基因发掘研究。当发明人以粳稻中花11为材料,使用甲基磺酸乙酯(ems)处理中花11的种子,随后经过高低温下筛选的过程中,意外获得了温敏核不育系tms11。
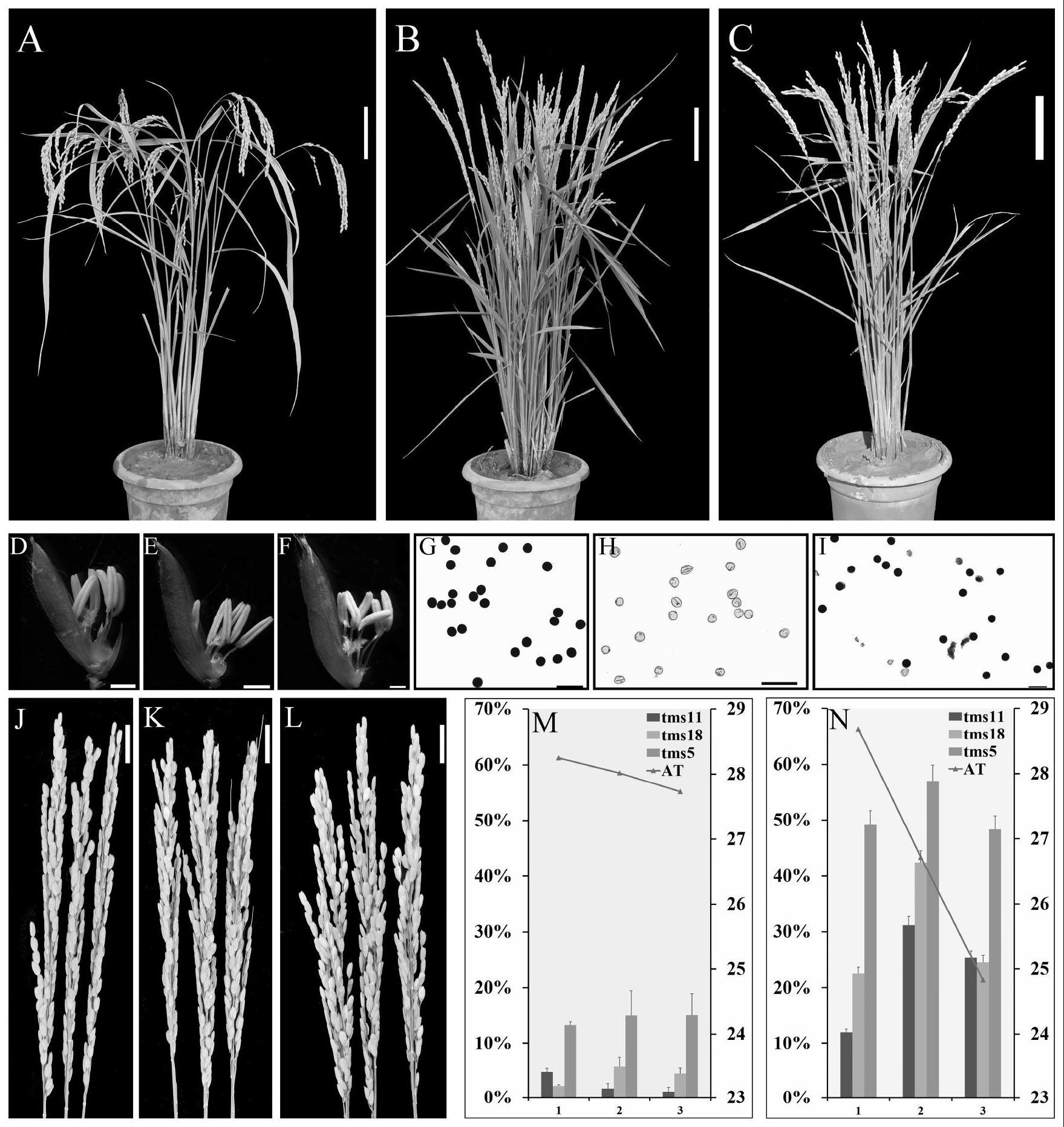
3、发明人发现,与野生型相比含有该突变体tms11的植株,营养生长没有差异。在高温下突变体tms11植株表现为完全不育。低温(<23℃)下突变体tms11植株恢复部分育性。使用碘-碘化钾染液对成熟时期的野生型和tms11的花粉进行染色发现高温下tms11几乎没有可以正常着色的花粉,而在低温下出现数量可观的正常着色的花粉。以上结果表明,水稻突变体tms11植株的育性是受温度影响的,高温使其育性受损不能产生正常有活力的花粉,而低温则可以恢复水稻tms11的育性。
4、因此,一方面本发明提供了一种水稻温敏不育基因突变体tms11,所述水稻温敏不育基因突变体tms11的核苷酸序列包含序列表中seq id no.1所示的核苷酸序列。
5、进一步地,所述水稻温敏不育基因突变体tms11的核苷酸序列由序列表中seq idno.1所示的核苷酸序列构成。
6、另一方面,本发明提供一种水稻温敏不育基因突变体tms11,所述水稻温敏不育基因突变体tms11的氨基酸序列如序列表中seq id no.2所示。
7、另一方面,本发明提供一种含有所述的水稻温敏不育基因突变体tms11或tms11的表达载体。
8、另一方面,本发明提供一种所述的水稻温敏不育基因突变体tms11或tms11的应用,其特征在于,所述水稻温敏不育基因突变体tms11或tms11用于制备隐性雄性核不育转基因植物。
9、另一方面,本发明提供一种的水稻温敏不育基因突变体tms11的应用,其用于水稻育种,所述应用包括(1)将含有水稻温敏不育基因突变体tms11的氨基酸序列或者核苷酸序列导入水稻;
10、(2)或将其通过杂交导入到水稻其他品种中,f2代中得到的不育植株同样也能显示温敏不育表型的;
11、(3)或者以(1)和(2)中获得的不育植株为母本,用不同的水稻品种作为父本进行杂交,培育杂交目标水稻,获得相应杂交种子。
12、另一方面,本发明提供一种培育在水稻花粉发育受温度影响的育性可恢复水稻的方法,所述方法包括将所述水稻温敏不育基因突变体tms11导入到水稻种子细胞中,利用导入了所述温敏不育基因突变体tms11的水稻种子进行相应水稻培育。(将该基因导入水稻细胞的方法可以采用现有方法,比如cripsr方法,abe或者cbe方法)
13、另一方面,本发明提供一种所述的水稻温敏不育基因突变体tms11或tms11的用途,其用于调节或提供植物的温敏育性性状,或被用作转基因植物的选择标记。
14、进一步地,所述选择标记的标记性状为温敏育性性状的可逆性变化。
15、进一步地,所述温敏育性性状的可逆性变化指在低温条件下,植物表现为育性恢复的性状;在高温条件下,植物表现为不育性状,所述低温条件为低于等于23℃,所述高温条件为高于等于28℃。
16、其中,tms11的核苷酸序列为
17、cds:
18、
19、tms11的氨基酸序列为:
20、
21、
22、技术效果
23、本发明的温敏不育突变体能够给植物,尤其是水稻带来良好的温敏不育特性,其相应植株在高温下表现为完全不育,而在低温(<23℃)下恢复部分育性。这种可以使育性完全丧失之后,又在特定条件下可以恢复的突变体对育种过程中的品种筛选以及纯度保证至关重要,是非常优质的基因资源,对我国水稻育种具有重要意义。
- 还没有人留言评论。精彩留言会获得点赞!