水稻节律调控蛋白ELF3-1及其编码基因在抗病育种中的应用
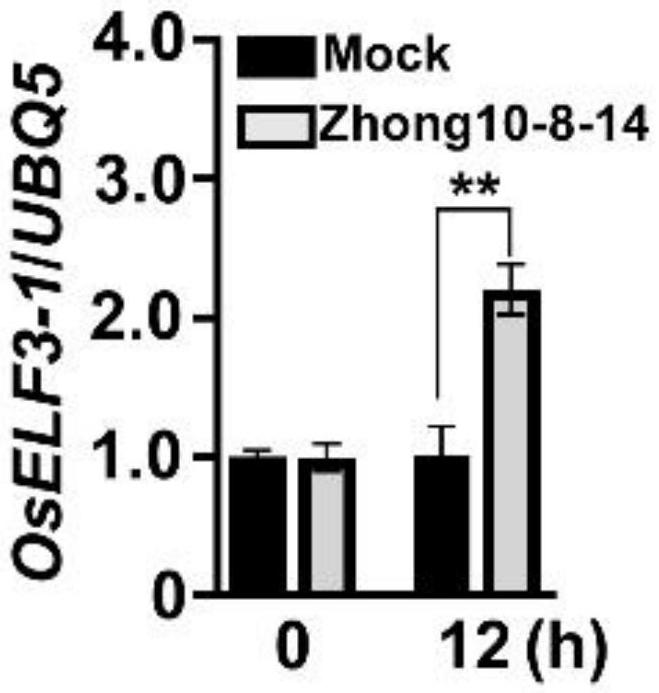
本发明涉及生物,特别是涉及水稻节律调控蛋白elf3-1及其编码基因在抗病育种中的应用。
背景技术:
1、真菌病害容易造成农作物的大规模减产,同时也在一定程度上影响了食品安全,因此培育抗病作物是种植业的主要目标之一。目前已报道了多种与植物抗病相关的基因,包括效应分子基因和调控基因,他们来源于水稻、小麦、玉米、大豆等农作物以及模式植物拟南芥等。其中一些已作为目的基因用于作物抗病基因工程,并成功培育出抗病水稻、小麦、玉米以及大豆等。但是由于遗传背景的差异,导致一些基因存在应用障碍,因此仍然需要新的相关基因来满足育种的需要。
2、在植物中,elf3蛋白通常被认为是一种光周期敏感的蛋白,它在调控植物的昼夜节律、开花时间、叶片形态和光合作用等方面起着关键作用。在水稻中,oself3同样参与到光周期信号途径中,通过与其他光周期相关基因相互作用,共同决定水稻的开花时间。例如,oself3基因的克隆和功能验证表明,它的正常功能对于水稻在适宜的时间抽穗至关重要。如果oself3基因发生突变或表达异常,可能会导致水稻过早或过晚抽穗,这将直接影响到水稻的产量和品质。然而,水稻elf3蛋白是否与抗病性有关,目前并未见相关报道。
技术实现思路
1、本发明的目的是提供水稻节律调控蛋白elf3-1及其编码基因在抗病育种中的应用,以解决上述现有技术存在的问题。本发明鉴定到一个表达受稻瘟菌诱导的水稻节律调控对蛋白elf3-1,经研究表明,与野生型水稻植株(zh11)相比,elf3-1功能缺失植株(elf3-1-kos)的稻瘟病抗性显著增强,说明elf3-1负调控水稻对稻瘟病的抗性,可作为目的基因用于提高植物抗病性的分子育种。
2、为实现上述目的,本发明提供了如下方案:
3、本发明提供水稻节律调控蛋白elf3-1及其编码基因elf3-1在调控水稻抗病性中的应用,敲除所述水稻节律调控蛋白elf3-1的编码基因elf3-1,能够提高水稻对稻瘟病的抗病性;
4、所述水稻节律调控蛋白elf3-1包括氨基酸序列如seq id no.4所示的蛋白或将seq id no.4所示的氨基酸序列经过一个以上氨基酸残基的取代和/或缺失和/或添加得到的与seq id no.4所示的蛋白质具有80%以上的同一性且具有负调控植物抗病性的蛋白质;
5、上述蛋白质中,同一性是指氨基酸序列的同一性。可使用国际互联网上的同源性检索站点测定氨基酸序列的同一性,如ncbi主页网站的blast网页。例如,可在高级blast2.1中,通过使用blastp作为程序,将expect值设置为10,将所有filter设置为off,使用blosum62作为matrix,将gap existence cost,per residue gap cost和lambda ratio分别设置为11,1和0.85(缺省值)并进行检索一对氨基酸序列的同一性进行计算,然后即可获得同一性的值(%)。
6、上述蛋白质中,所述80%以上的同一性可为至少81%、82%、85%、86%、88%、90%、91%、92%、95%、96%、98%、99%或100%的同一性。
7、所述编码基因elf3-1的核苷酸序列如seq id no.3所示。
8、本发明还提供一种调控水稻抗病性的方法,包括敲除水稻节律调控蛋白elf3-1的编码基因elf3-1,抑制或降低或下调所述水稻节律调控蛋白elf3-1的表达量,提高水稻对稻瘟病的抗病性;
9、所述敲除水稻节律调控蛋白elf3-1的编码基因elf3-1的物质包括敲除所述编码基因elf3-1的重组载体或重组微生物,所述重组载体包含靶向所述编码基因elf3-1的grna,所述grna的靶点序列如seq id no.5所示。
10、所述敲除水稻节律调控蛋白elf3-1的编码基因elf3-1的物质还包括含有编码所述水稻节律调控蛋白elf3-1的核酸分子或含有所述核酸分子的表达盒,所述的含有核酸分子的表达盒,是指能够在宿主细胞中表达上述应用中所述蛋白质的dna,该dna不但可包括启动蛋白编码基因转录的启动子,还可包括终止蛋白编码基因转录的终止子。
11、进一步的,所述表达盒还包括增强子序列。可用于本发明的启动子包括但不限于:组成型启动子,组织、器官和发育特异的启动子,和诱导型启动子。
12、可用现有的植物表达载体构建敲除所述编码基因elf3-1的重组载体。所述植物表达载体包括双元农杆菌载体和可用于植物微弹轰击的载体等。如pahc25、pwmb123、pbin438、pcambia1302、pcambia2301、pcambia2300、pcambia1301、pcambia1300、pbi121、pcambia1391-xa或pcambia1391-xb(cambia公司)等。所述植物表达载体还可包含外源基因的3’端非翻译区域,即包含聚腺苷酸信号和任何其它参与mrna加工或基因表达的dna片段。所述聚腺苷酸信号可引导聚腺苷酸加入到mrna前体的3’端,如农杆菌冠瘿瘤诱导(ti)质粒基因(如胭脂碱合成酶基因nos)、植物基因(如水稻淀粉合成酶基因)3’端转录的非翻译区均具有类似功能。构建敲除所述编码基因elf3-1的重组载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。
13、本发明还提供水稻节律调控蛋白elf3-1及其编码基因elf3-1在培育高抗病性水稻中的应用,敲除所述水稻节律调控蛋白elf3-1的编码基因elf3-1,获得的水稻植株对稻瘟病的抗病性增强。
14、本发明还提供水稻节律调控蛋白elf3-1及其编码基因elf3-1在对水稻进行基因编辑辅助育种中的应用。
15、进一步地,所述基因编辑为利用crispr基因编辑技术敲除所述水稻节律调控蛋白elf3-1的编码基因elf3-1。
16、进一步地,敲除所述水稻节律调控蛋白elf3-1的编码基因elf3-1具体包括构建敲除所述编码基因elf3-1的重组载体,利用所述重组载体对所述编码基因elf3-1进行敲除,所述重组载体包含靶向所述编码基因elf3-1的grna,所述grna的靶点序列如seq id no.5所示。
17、本发明公开了以下技术效果:
18、本发明鉴定到一个表达受稻瘟菌诱导的水稻节律调控蛋白elf3-1,其编码基因位于6号染色体的loc_os06g05060(elf3-1)。以往该基因的功能研究主要集中于水稻节律和开花时间的调控。本发明研究表明,与野生型水稻植株(zh11)相比,elf3-1功能缺失植株(elf3-1-kos)的稻瘟病抗性显著增强。这些结果说明elf3-1负调控水稻对稻瘟病的抗性,可作为目的基因用于提高植物抗病性的分子育种。
19、本发明为培育高抗病性水稻品种提供了一个新的基因,对改善水稻抗病性具有重要价值。
- 还没有人留言评论。精彩留言会获得点赞!