SNARE蛋白在调控溶瘤新城疫病毒复制水平中的应用
本发明属于生物,具体涉及snare蛋白在调控溶瘤新城疫病毒复制水平中的应用。
背景技术:
1、新城疫病毒(ndv)是单股、负链、不分节段的rna病毒,是副粘病毒科、禽副粘病毒属的唯一成员。ndv是对家禽养殖业具有重要意义的病原,同时也是一种强效的溶瘤病毒。ndv可感染多种肿瘤细胞,直接诱导肿瘤细胞的死亡,并调控肿瘤微环境与免疫微环境,促进机体的抗肿瘤反应。
2、ndv的溶瘤活性已在多种肿瘤细胞、肿瘤动物模型以及人的临床治疗试验中得到了验证。但是,研究显示,某些类型的肿瘤细胞可产生较强的抗病毒反应,抑制ndv的复制,减弱了病毒的溶瘤活性。目前,现有技术主要通过以下三个途径增强ndv的溶瘤活性:(1)改变ndv融合蛋白(f)的裂解位点,增强病毒的融合活性。(2)在ndv基因中表达免疫调控性的细胞因子。(3)在ndv基因组中表达免疫检查点抑制剂。但是,改变ndv f蛋白裂解位点,导致病毒的毒力增强,增加了病毒的环境释放和对鸟类的致病风险;在ndv基因组中表达细胞基因或免疫检查点抑制剂,主要通过肿瘤免疫的调控增强病毒的溶瘤活性,但是并不能增强病毒本身的复制能力。
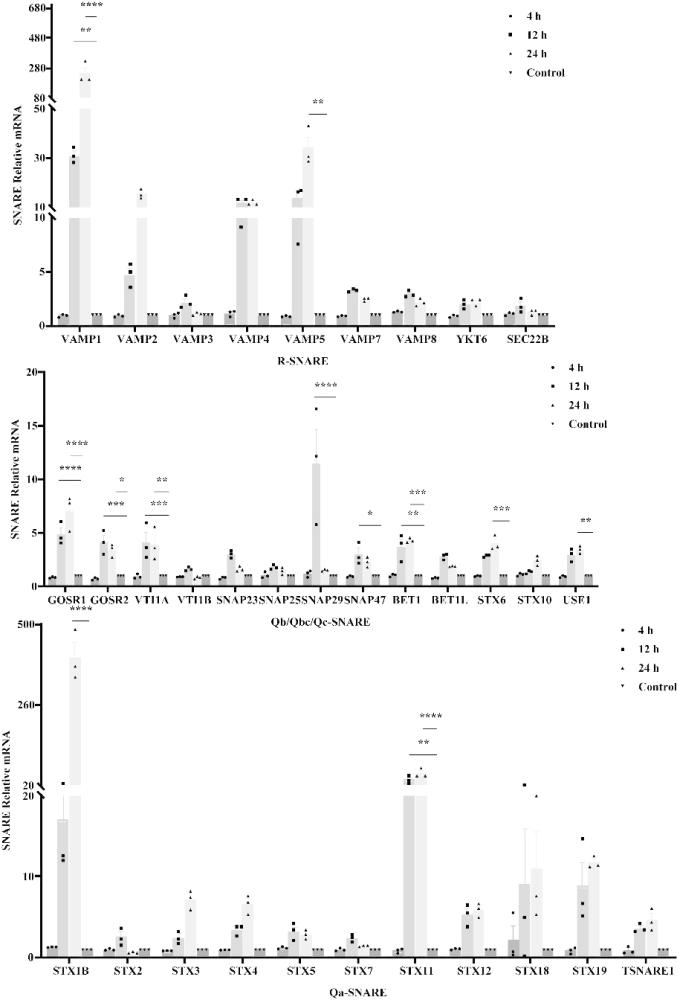
3、ndv进入宿主细胞后,病毒基因组在宿主细胞内进行转录,翻译的病毒蛋白最终被转运至细胞膜上,组装成病毒粒子并通过出芽的方式完成释放。新合成的病毒蛋白必须劫持宿主细胞的物质运输系统而被转运至细胞膜出芽位点。囊泡运输是细胞内大分子物质运输的主要路径,而膜融合是囊泡运输的最后阶段,是将货物卸载到目的地的关键步骤。可溶性n-乙基马来酰亚胺敏感因子连接蛋白受体(snare)是介导膜融合的最小核心机器。snare对多种病毒的蛋白胞内转运、组装以及复制有重要的调控作用。因此,snare蛋白可能在ndv复制过程中发挥重要作用,而snare蛋白是增强ndv组装与复制的潜在靶点。
技术实现思路
1、针对现有技术在增强溶瘤ndv复制能力上的不足和缺陷,本发明提供snare蛋白在调控溶瘤新城疫病毒复制水平中的应用,以ndv与snare蛋白的相互作用为切入点,首先筛选了ndv感染人非小细胞肺癌细胞(a549)、人宫颈癌上皮细胞(hela)中表达发生显著变化的snare蛋白基因;接着用不同策略评价了snare蛋白对ndv在hela细胞中复制水平的影响;最后评价了snare家族成员ykt6蛋白对ndv f蛋白转运与病毒粒子组装的影响。本发明证明ndv感染a549与hela细胞可显著上调大量sanre基因的表达;选择代表性的sanre蛋白ykt6为研究对象,证明ykt6蛋白功能突变体过表达以及ykt6蛋白的敲降均导致ndv复制水平的下降,而过表达ykt6蛋白可显著增强病毒复制。ykt6蛋白通过促进ndv f蛋白的膜转运,增强病毒粒子的组装与病毒复制。本发明提供了ykt6蛋白在增强ndv在肿瘤细胞中复制的新作用,以及ykt6蛋白可作为促进ndv溶瘤活性的新的靶标,在人与动物肿瘤的治疗方面有潜在的应用前景。
2、为达到上述目的,本发明采用的技术方案如下:
3、snare蛋白在调控溶瘤新城疫病毒复制水平中的应用,所述snare蛋白的序列为genebank accession no. nm_001297438.2、nm_001145149.3、nm_001286210.1、nm_018467.3、nm_003764.4、nm_017919.3、nm_001318203.2所示序列中的一种,所述snare蛋白不包含跨膜区。
4、进一步的,所述snare蛋白为ykt6蛋白,所述ykt6蛋白的序列如genebankaccession no. nm_006555.4所示。
5、进一步的,所述应用包括降低溶瘤新城疫病毒囊膜糖蛋白在细胞质中的表达,降低细胞质中病毒np、m、f蛋白的表达。
6、进一步的,敲降ykt6蛋白,使ykt6蛋白的表达水平降低。
7、进一步的,靶向ykt6的sirna序列如seq id no. 95与seq id no. 96所示。
8、进一步的,用于ykt6基因表达测定的荧光定量pcr如引物seq id no. 1与seq idno. 2所示。
9、进一步的,用于ykt6基因干扰效率测定的荧光定量pcr引物序列如seq id no. 97和seq id no. 98所示。
10、进一步的,将ykt6蛋白第194或195位的半胱氨酸位点突变成丝氨酸,以pcdna3.1为载体构建功能突变体。
11、进一步的,将ykt6蛋白第194位的半胱氨酸位点突变成丝氨酸,构建得到ykt6-c194s突变体,构建ykt6-c194s突变体的引物如seq idno.89和seq idno.90所示;将ykt6蛋白第195位的半胱氨酸位点突变成丝氨酸,构建得到ykt6-c195s突变体,构建ykt6-c195s突变体的引物如seq id no.91和seq id no.92所示。
12、本发明还提供一种降低溶瘤新城疫病毒复制水平的方法,敲降ykt6蛋白,使ykt6蛋白的表达水平降低,所述ykt6蛋白的序列如genebank accession no. nm_006555.4所示。
13、本发明设计了针对36个人源sanre基因以及β-actin的引物,序列见seq id no. 1至seq id no. 74。利用荧光定量pcr技术测定了不同ndv毒株(herts/33、f48e8、la sota)感染a549或hela细胞中snare基因的转录水平。
14、进一步的,选择snare家族成员ykt6蛋白为代表,用western blot方法测定了ndvf48e8与la sota感染hela细胞中ykt6蛋白的表达水平。
15、进一步的,将vamp1、vamp7、stx6、use1、stx11、stx17以及vti1a蛋白的跨膜区删除,使用pegfp-c3载体构建了以上蛋白的功能突变体,所用引物序列如seq id no. 75至seq id no. 88所示;将ykt6蛋白第194、195位的半胱氨酸位点突变成丝氨酸,使用了pcdna3.1载体构建其功能突变体,所用引物序列如seq id no.89至seq id no.92所示。
16、进一步的,在hela细胞中过表达vamp1、vamp7、stx6、use1、stx11、stx17、vti1a与ykt6蛋白的功能突变体,再用ndv感染,测定病毒的复制水平。
17、进一步的,合成了针对ykt6蛋白的小干扰rna(sirna),序列如seq id no. 95与seq id no. 96所示。将sirna转染细胞,用荧光定量pcr方法验证sirna对ykt6基因转录水平的影响,所用荧光定量pcr的引物序列如seq id no. 97与seq id no. 98所示;用western blot方法验证sirna对ykt6蛋白表达的影响。用ndv感染ykt6-sirna转染的hela细胞,评价病毒复制水平。
18、进一步的,构建ykt6表达质粒,并用overlap pcr方法将其中sirna的识别位点突变,使其对sirna具有抗性,所用突变引物序列如seq id no. 99、seq id no. 100、seq idno. 101、seq id no. 102所示。在ykt6敲降的基础上再回补ykt6蛋白,之后进行ndv感染,测定病毒的复制水平。
19、进一步的,先在hela细胞中转染ykt6-sirna,再用表达ndv基质蛋白(m)、核衣壳蛋白(np)、f蛋白与血凝素-神经氨酸酶(hn)的质粒共转染细胞,收集培养上清,用超速离心法纯化上清中的病毒样颗粒;用western blot方法测定ndv病毒样颗粒(vlp)的组装效率。
20、进一步的,在ykt6-sirna转染的hela细胞中共转染ndv的m、np、f与hn表达质粒,分离细胞膜与细胞质蛋白,用western blot方法测定病毒蛋白转运至细胞膜的效率。
21、进一步的,构建表达ykt6蛋白的表达质粒,所用引物序列见seq id no. 99与seqid no. 102。通过质粒转染的方法,在hela细胞中过表达ykt6蛋白,再感染ndv,测定病毒的复制水平。
22、有益效果
23、本发明提供了snare家族成员ykt6蛋白在促进溶瘤ndv组装与复制中的新的应用,以及ykt6蛋白作为促进ndv溶瘤活性中的新的靶标,为利用溶瘤ndv进行人与动物肿瘤的治疗提供了新策略和新靶标。
- 还没有人留言评论。精彩留言会获得点赞!