特异性生产类菌孢素氨基酸的MysDs酶及其制备方法与流程
本技术涉及类菌孢素氨基酸生产,特别是涉及一类特异性生产类菌孢素氨基酸的mysds酶及其制备方法。
背景技术:
1、紫外线(uv)是太阳光的一部分,它可以分为三种类型:uva、uvb和uvc。uvc被地球大气层中的臭氧层吸收,所以不会到达地面。而uva(315-400nm)和uvb(280-315nm)都能到达地表并对人体造成一系列危害,如皮肤癌,皮肤老化,免疫系统抑制和眼部损伤等。
2、类菌孢素氨基酸(mycosporine-like amino acids,maas)是一类具有紫外线吸收特性的天然化合物,由于其生物相容性和较低的环境影响,它们被视为比传统防晒剂如锌氧化物和氧苯酮更为理想的防晒成分。maas还表现出抗氧化、免疫调节、抗衰老和抗炎等多种生物活性。这些特性使得maas成为化妆品和护肤品行业的重要原料。
3、传统的maas制备方法,主要是通过从从红藻(如紫菜porphyra属)、绿藻以及某些种类的蓝细菌中提取获得。这种从天然资源中提取maas的制备方法存在产量低的问题,需要大量收获天然藻类;在过度采集的情况下,这种方法还可能会对生态环境造成压力。而且自然条件下生长的藻类会受到季节变化和地理位置的影响,导致maas含量的变化,从而影响maas提取的稳定性和一致性。
4、为了解决传统方法提取maas所造成的问题,大多数研究学者主要想要通过代谢工程改造微生物来生产maas,主要在酿酒酵母(saccharomyces cerevisiae)和耶氏解脂酵母(yarrowia lipolytica)等微生物中改造。maas的生物合成基因簇包含三个保守基因,即编码ddg合酶(2-demethyl 4-deoxygadusol synthase,ddgs)的mysa基因,编码sam依赖的o-甲基转移酶(o-methyltransferase,o-mt)的mysb基因,以及编码atp-grasp酶的mysc基因。mysa和mysb催化磷酸七糖(sedoheptulose 7-phosphate,s7p)从磷酸戊糖途径(ppp)逐步转化为所有maas的骨架4-脱氧鳕醇(4-deoxygadusol,4-dg),而mysc随后催化4-dg与甘氨酸,产生菌孢素-甘氨酸(mycosporine-glycine,mg)。专利号为202211236882.2的专利文件中,通过引入木糖代谢途径,敲除tal1和nqm1基因以增加前体s7p的量,再多拷贝插入ddgs-omt,从而增加了4-dg的量,进而为构建生产maas直接前体物质mg的菌株(sc.l26)提供基础,如图1所示。
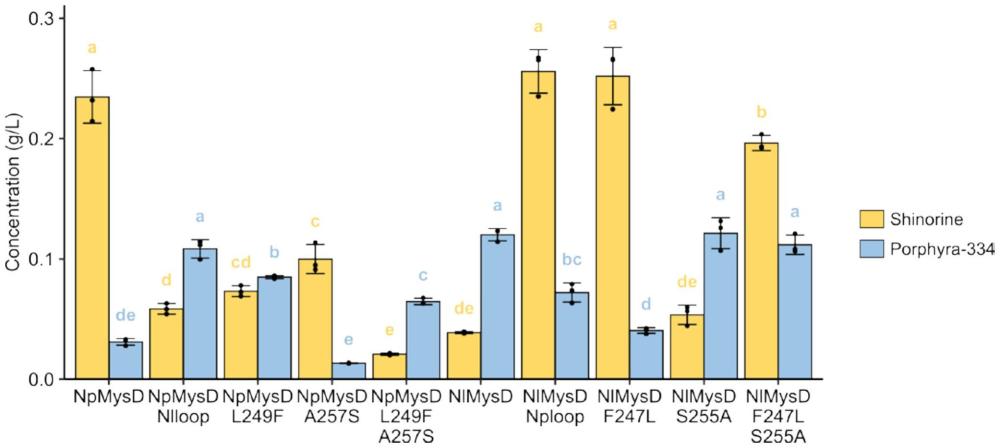
5、maas的基因簇还包括mysd,它编码d-丙氨酸-d-丙氨酸连接酶(d-ala-d-ala-ligase)的同源物,即mysds酶。mysds酶能够催化氨基酸与mg的结合,产生不同类型的二取代maas,包括shinorine(mg+l-丝氨酸)和porphyra-334(mg+l-苏氨酸)。因此通过代谢工程改造,在微生物以内引入mysa、mysb、mysc和mysd等基因可以生产多种类型的maas,如shinorine、porphyra-334和mycosporine-2-glycine等。目前已公开的专利文献中,au2019224876a1,us2021230610a1中sol kim,seoul(kr)通过微生物中引入2-脱氢-3-脱氧磷酸庚酸醛缩酶(2-dehydro-3-deoxyphosphoheptonate aldolase)、磷酸烯醇丙酮酸合成酶(2-dehydro-3-deoxyphosphoheptonate aldolase)和转酮醇酶蛋白(transketolase)的活性从而提高微生物生产maas的产量。此外,在专利文献us2023064780a1中ji-sook hahn,seoul(kr)通过引入木糖途径,敲除tal基因来增加前体物质s7p,进而提高maas的产量。但是目前生物工程改造生产maas的微生物,都会产生多种二取代maas,主要原因是mysds酶对底物特异性差,造成副产物的形成,其中maas主要是shinorine和porphyra-334的混合物。
6、因此,如何提高工程改造微生物生产maas的特异性,仍然是maas生产技术领域的研究重点和难点。
技术实现思路
1、本技术的目的是提供一类新的特异性生产类菌孢素氨基酸的mysds酶及其制备方法。
2、本技术采用了以下技术方案:
3、本技术的第一方面公开了特异性生产类菌孢素氨基酸的mysds酶,该mysds酶含有26个氨基酸的loop区域,并且,loop区域为以下两类情况;
4、第一类mysds酶:loop区域的第25个氨基酸为a,且第17个氨基酸为除f以外的任意氨基酸;或者,loop区域的第25个氨基酸为s,且第17个氨基酸为l;第一类mysds酶用于特异性的生产shinorine;
5、第二类mysds酶:loop区域的第25个氨基酸为s,且第17个氨基酸为除l以外的任意氨基酸;或者,loop区域的第25个氨基酸为a,且第17个氨基酸为f;第二类mysds酶用于特异性的生产porphyra-334。
6、需要说明的是,本技术的关键在于,研究发现mysds酶的26个氨基酸的loop区域能够有效的决定类菌孢素氨基酸生产的特异性,即特异性的生产shinorine或porphyra-334;该发现与ji-sook hahn,seoul(kr)团队发现的43-45个氨基酸序列的loop区域相比,本技术的loop区域更简短,更容易编辑和操作。并且,进一步的,本技术研究发现,本技术的26个氨基酸的loop区域中,第一类mysds酶能够特异性的生产shinorine,第二类mysds酶能够特异性的生产porphyra-334。
7、也就是说,将本技术的26个氨基酸的loop区域的第25个氨基酸突变为a,且第17个氨基酸为除f以外的任意氨基酸;或者,将loop区域的第25个氨基酸突变为s,且第17个氨基酸突变为l;这样的mysds酶或突变体,即第一类mysds酶,能够特异性的生产shinorine。将本技术的26个氨基酸的loop区域的第25个氨基酸突变为s,且第17个氨基酸为除l以外的任意氨基酸;或者,将loop区域的第25个氨基酸突变为a,且第17个氨基酸突变为f;这样的mysds酶或突变体,即第二类mysds酶,能够特异性的生产porphyra-334。
8、本技术中,特异性的生产shinorine或porphyra-334是指,mysds酶能够偏向性的生产shinorine或porphyra-334;例如,第一类mysds酶能够特异性的生产shinorine是指,与现有技术相比,采用第一类mysds酶获得的产物中大部分或者超过50%的产物为shinorine,其余仍然有少部分为porphyra-334,因此说第一类mysds酶具有shinorine偏向性,能够特异性的生产shinorine;同样的,第二类mysds酶能够特异性的生产porphyra-334是指,采用第二类mysds酶获得的产物中大部分或者超过50%的产物为porphyra-334,其余仍然有少部分为shinorine,因此说第二类mysds酶具有porphyra-334偏向性,能够特异性的生产porphyra-334。
9、还需要说明的是,本技术的mysds酶含有26个氨基酸的loop区域,是因为本技术研究发现这26个氨基酸的loop区域即可有效的决定mysds酶生产类菌孢素氨基酸的特异性;并且,限定26个氨基酸的loop区域是为了更好的对其中的第17个氨基酸和第25个氨基酸进行定位。原则上,在本技术的26个氨基酸的基础上,还可以根据需求或设计增加若干个氨基酸,从而形成更长的loop区域,只要符合本技术限定的第17个氨基酸和第25个氨基酸的突变类型,即可实现特异性的生产shinorine或porphyra-334。
10、本技术的一种实现方式中,loop区域为seq id no.1至seq id no.22中的至少一种;其中,第一类mysds酶的loop区域为seq id no.1至seq id no.10所示序列中的至少一种;第二类mysds酶的loop区域为seq id no.11至seq id no.22所示序列中的至少一种。
11、需要说明的是,seq id no.1至seq id no.22所示序列的loop区域,仅仅是本技术的实施例中具体采用的能够特异性生产类菌孢素氨基酸的22种loop区域。并且通过这22种loop区域,有效的证实和验证了loop区域的第25个氨基酸为a,且第17个氨基酸为除f以外的任意氨基酸,或loop区域的第25个氨基酸为s,且第17个氨基酸为l,能特异性的生产shinorine;loop区域的第25个氨基酸为s,且第17个氨基酸为除l以外的任意氨基酸,或loop区域的第25个氨基酸为a,且第17个氨基酸为f,能特异性的生产porphyra-334。
12、本技术的一种实现方式中,第一类mysds酶为seq id no.23至seq id no.32所示序列中的至少一种;第二类mysds酶为seq id no.33至seq id no.44所示序列中的至少一种。
13、需要说明的是,seq id no.23至seq id no.32所示序列的第一类mysds酶,seq idno.33至seq id no.44所示序列的第二类mysds酶,是本技术的实施例中具体采用的与seqid no.1至seq id no.22所示序列的loop区域对应的mysds酶,通过这些mysds酶有效的证实和验证了第17个氨基酸和第25个氨基酸的突变对mysds酶特异性生产类菌孢素氨基酸的影响,即第25个氨基酸为a,且第17个氨基酸为除f以外的任意氨基酸,或loop区域的第25个氨基酸为s,且第17个氨基酸为l,能特异性的生产shinorine;loop区域的第25个氨基酸为s,且第17个氨基酸为除l以外的任意氨基酸,或loop区域的第25个氨基酸为a,且第17个氨基酸为f,能特异性的生产porphyra-334。
14、可以理解,只要符合以上第17个氨基酸和第25个氨基酸的突变规律,都能够实现特异性的生产shinorine或porphyra-334,不仅限于本技术限定的seq id no.1至seq idno.22所示序列的22种loop区域,也不仅限于seq id no.23至seq id no.44所示序列的22种mysds酶。
15、本技术的第二方面公开了编码本技术的mysds酶的核酸。
16、本技术的第三方面公开了含有本技术的核酸的重组质粒。
17、需要说明的是,含有本技术核酸的重组质粒,能够转化到工程菌中表达本技术的mysds酶,从而实现特异性的生产shinorine或porphyra-334。
18、本技术的第四方面公开了转化有本技术的重组质粒的微生物。
19、需要说明的是,转化有本技术的重组质粒的微生物,实际上就是用于表达本技术的mysds酶的工程菌,以实现特异性的生产shinorine或porphyra-334。
20、本技术的一种实现方式中,微生物为重组生物体,重组生物体为敲除内源基因后,将木糖代谢基因、ddgs基因、omt基因整合至生物体的基因组中获得的可使用木糖生产4dg的生物体;其中,敲除的内源基因包括tal1基因、nqm1基因,木糖代谢基因包括xyl1基因、xyl2基因、xyl3基因,xyl1基因、xyl2基因、xyl3基因中的至少一种的插入位点为his3位点。
21、优选的,木糖代谢基因中,使用ptdh3启动子启动xyl1基因的表达,使用ppgk1启动子启动xyl2基因的表达,使用ptef2启动子启动xyl3基因的表达。
22、优选的,本技术的生物体或工程菌为酵母。
23、需要说明的是,本技术的重组生物体实际上就是本技术发明人之前的专利号为202211236882.2中构建的重组生物体;可以理解,在专利202211236882.2中构建的重组生物体的基础上,结合本技术的第一类mysds酶或者第二类mysds酶,即可实现特异性的生产shinorine或porphyra-334。并且,同样能够获得专利202211236882.2中重组生物体的优势效果,即在提高类菌孢素氨基酸生产特异性的同时,提高类菌孢素氨基酸的产量。因此,专利202211236882.2中所有关于重组生物体构建的技术内容援引至本技术。
24、本技术的第五方面公开了本技术的mysds酶、本技术的核酸、本技术的重组质粒或者本技术的微生物在特异性生产类菌孢素氨基酸中的应用。
25、需要说明的是,本技术的微生物即能够表达本技术的mysds酶的工程菌,因此,能够用于特异性的生产shinorine或porphyra-334;当然,根据不同的设计或生产需求,还可以对工程菌进行一系列的改造,例如采用专利202211236882.2中改进的重组生物体。至于本技术的核酸和重组质粒可以作为原材料制备表达本技术mysds酶的工程菌。
26、本技术的第六方面公开了一种特异性生产类菌孢素氨基酸的方法,包括采用本技术的mysds酶、本技术的核酸、本技术的重组质粒或者本技术的微生物,特异性的生产shinorine或porphyra-334。
27、本技术的一种实现方式中,具体包括采用本技术的mysds酶中的第一类mysds酶,或者本技术的编码第一类mysds酶的核酸,或者本技术的含有编码第一类mysds酶的核酸的重组质粒,或者本技术的转化有含有编码第一类mysds酶的核酸的重组质粒的微生物,特异性的生产shinorine;或者,采用本技术的mysds酶中的第二类mysds酶,或者本技术的编码第二类mysds酶的核酸,或者本技术的含有编码所述第二类mysds酶的核酸的重组质粒,或者本技术的转化有含有编码所述第二类mysds酶的核酸的重组质粒的微生物,特异性的生产porphyra-334。
28、需要说明的是,本技术的类菌孢素氨基酸的生产方法,能够特异性的生产shinorine或porphyra-334,解决了工程改造微生物生产类菌孢素氨基酸的特异性差的问题;并且,结合专利202211236882.2中改进的重组生物体,能够进一步的提高类菌孢素氨基酸的产量。
29、本技术的有益效果在于:
30、本技术的mysds酶,底物特异性好,能够有效的提高shinorine或porphyra-334的生产特异性,提高最终产品中单一类菌孢素氨基酸的纯度和质量,解决了工程改造微生物生产类菌孢素氨基酸的特异性问题。
- 还没有人留言评论。精彩留言会获得点赞!