一种黑暗诱导的增强型发光表达载体及其构建方法和应用与流程
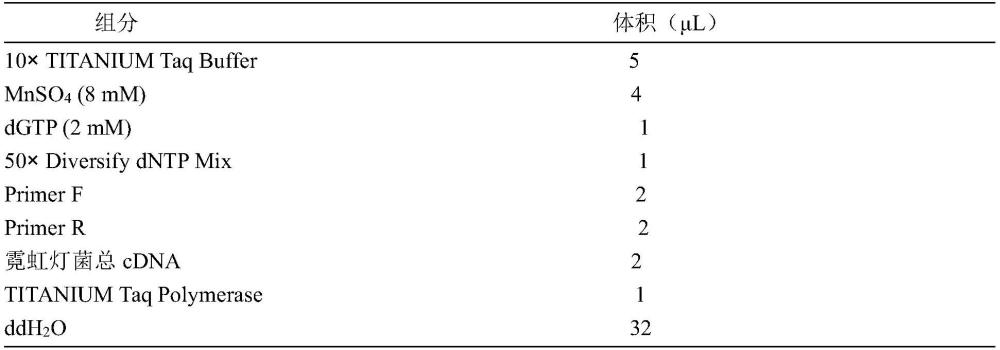
本发明涉及生物,具体涉及一种黑暗诱导的增强型发光表达载体及其构建方法和应用。
背景技术:
1、生物发光是生物体中由荧光素和荧光素酶发生生化反应产生的冷光(“coldlight”),是几个世纪以来一直令人着迷的自然现象。生物发光广泛存在于各种生物中,从低等的原核生物到高等的真核生物都存在生物发光现象,目前世界上已报道的发光类群约有800属约1万种。已知的不会发光的谱系有陆生植物和脊椎动物,如鸟类、两栖动物和哺乳动物。为利用自然界中的生物发光现象,科学家们从发光生物中解析了生物发光基因以及发光原理,并将其工具化应用于科学研究中。
2、利用生物发光系统实现植物自发光是其重要应用之一。在自然界中存在至少40种不同的生物发光系统,目前仅解析了7种生物发光系统发光所需的荧光素酶和荧光素,其中只有2个生物发光系统的完整生化过程被解析,即细菌和真菌的生物发光系统。细菌生物发光系统的发现已历经40余年,由于其细胞毒性和发光效率低,目前仍无法利用该发光系统创制自发光植物。真菌发光系统自2018年解析以来,由于其发光底物咖啡酸在各种生物中广泛存在,因此仅需转入发光反应相关的基因而不需要额外添加外源发光底物即可实现生物自发光,为创制自发光植物打开了新的大门。利用该发光系统已在烟草、牵牛花等植物中实现肉眼可见的自发光。然而,利用真菌生物发光系统创制的发光植物发光颜色单一,发光时间不可控,发光亮度不够强。
技术实现思路
1、本发明的目的是针对上述问题,提供一种黑暗诱导的增强型发光表达载体及其构建方法和应用。
2、本发明为了实现其目的,采用的技术方案是:
3、本发明的第一方面提供一种黑暗诱导的增强型发光表达载体,是以载体pdes为骨架载体构建而成,是在载体pdes的lr clonase重组酶识别的特异重组位点attr1和attr2元件之间按照从5’到3’端方向插入由依次连接的启动子psdin6,突变的nnluz基因mnnluz,霓虹灯菌(neonothopanus nambi)的hisps基因、h3h基因、cph基因,以及构巢曲霉(aspergillusnidulans)的npga基因构成的核心元件获得;
4、所述启动子psdin6的序列如seq id no.1所示,突变的nnluz基因mnnluz的序列如seq id no.4所示;
5、所述载体pdes是将seq id no.12所示的序列与用asci线性化的pyltac7载体进行同源重组得到的。
6、所述核心元件是将如seq id no.5所示的表达框psdin6-mnnluz-tnos、如seq idno.6所示的表达框p35s-hisps-t35s、如seq id no.7所示的表达框p35s-h3h-cph-npga依次串联得到。
7、所述表达框是通过gateway克隆技术构建入载体pdes中的;
8、所述attr1的序列为(seq id no.13):
9、5’-acaagtttgtacaaaaaagctgaacgagaaacgtaaaatgatataaatatcaatatattaaattagattttgcataaaaaacagactacataatactgtaaaacacaacatatccagtcatattg-3’,所述attr2的序列为(seq id no.14):
10、5’-catagtgactggatatgttgtgttttacagcattatgtagtctgttttttatgcaaaatctaatttaatatattgatatttatatcattttacgtttctcgttcagctttcttgtacaaagtggt-3’。
11、本发明的第二方面提供上述任一项所述的黑暗诱导的增强型发光表达载体的构建方法,包括如下步骤:
12、s1、构建质粒pdonor1-psdin6-mnnluz-tnos:将如seq id no.5所示的psdin6-mnnluz-tnos表达框通过酶切位点asisi和asci亚克隆到载体pdonor1中得到质粒pdonor1-psdin6-mnnluz-tnos;
13、s2、构建植物表达载体pdes-psdin6-mnnluz-tnos:将质粒pdonor1-psdin6-mnnluz-tnos与载体pdes混合,通过同源重组获得pdes-psdin6-mnnluz-tnos;
14、s3、构建质粒pdonor2-p35s-hisps-t35s、pdonor1-p35s-h3h-cph-npga
15、将如seq id no.6所示的p35s-hisps-t35s表达框通过酶切位点asisi和asci亚克隆到载体pdonor2中得到质粒pdonor2-p35s-hisps-t35s;
16、将如seq id no.7所示的p35s-h3h-cph-npga表达框通过酶切位点asisi和asci亚克隆到载体pdonor1中得到质粒pdonor1-p35s-h3h-cph-npga;
17、s4、构建载体pdes-psdin6-mnnlu-hisps:将质粒pdonor2-p35s-hisps-t35s与质粒pdes-psdin6-mnnluz-tnos混合,通过同源重组得到质粒pdes-psdin6-mnnlu-hisps;
18、s5、构建载体pdes-psdin6-mnnlu-hisps-h3h-cph-npga:将质粒pdonor1-p35s-h3h-cph-npga与pdes-psdin6-mnnlu-hisps混合,通过同源重组得到表达载体
19、pdes-psdin6-mnnlu-hisps-h3h-cph-npga。
20、步骤s2、s4、s5中进行同源重组前,将质粒先用i-scei进行线性化。
21、载体pdonor1是将合成序列i与用ecor i线性化的pcrtm8/gw/topotm载体进行同源重组得到的,所述合成序列i的核苷酸序列是在seq id no.8所示的序列的5’端加上seq idno.9所示的序列,在3’端加上seq id no.10所示的序列得到的序列;
22、载体pdonor2是将合成序列ii与用sal i线性化的puc19载体进行同源重组得到的,所述合成序列ii的核苷酸序列是在seq id no.11所示的序列的5’端加上seq id no.9所示的序列,在3’端加上seq id no.10所示的序列得到的序列。
23、本发明的第三方面提供上述任一项所述的黑暗诱导的增强型发光表达载体在培育转基因植物中的应用。
24、所述转基因植物在黑暗条件下发荧光。
25、所述的应用,将上述的表达载体导入受体植物中,得到转基因植物;与所述受体植物相比,所述转基因植物在夜晚发出明亮的荧光,而白天不发光。
26、所述受体植物选自烟草、拟南芥、矮牵牛花;采用农杆菌介导,优选采用gv3101农杆菌。
27、本发明的有益效果是:
28、针对目前创制的自发光植物发光时间不可控、发光亮度不够强的问题,设计构建了黑暗诱导的增强型发光载体,将该载体转入植物后可以使植物在夜晚发出明亮的荧光,而白天不发光。通过高通量筛选来自拟南芥中受黑暗诱导的启动子,获得黑暗诱导表达水平最高的dark inducible 6(din6)启动子(pdin6)。为减少启动子长度,提高载体构建效率和稳定性,对启动子pdin6进行了优化。优化后的启动子psdin6的片段大小远小于pdin6,活性却与pdin6相当。使用小片段的psdin6,减少启动子长度更有利于表达载体的组装,提高载体构建成功率,避免因重组的片段太长导致无法成功构建重组载体或者重组载体无法发挥预期功能;同时也能避免因重组片段太长导致的重组载体在细菌或者农杆菌中扩繁时,载体容易突变或者缺失片段的问题,提高了载体的稳定性。
29、使用定向进化技术随机突变发光真菌霓虹灯菌neonothopanus nambi的荧光素酶luz因,筛选获得比原有发光效率高5.7倍的荧光素酶突变基因mluz。
30、利用优化后的黑暗诱导型启动子psdin6驱动mluz基因的表达,同时,该载体插入了植物发光所必需的霓虹灯菌hisps基因、h3h基因、cph基因,以及构巢曲霉(aspergillusnidulans)npga基因,成功构建了黑暗诱导的增强型发光载体。将本发明构建的表达载体在烟草叶片上瞬时表达,实现了烟草叶片白天不发光而黑暗下发出明亮的荧光,为发光植物白天积累能量晚上释放光芒提供了新的路径。
- 还没有人留言评论。精彩留言会获得点赞!