具有整合泵送、调平和感测的模块化器官微生理系统的制作方法
本申请要求于2016年2月4日提交的美国临时申请号62/291,102以及于2016年7月7日提交的美国临时申请号62/359,567的优先权和权益,这些美国临时申请特此通过引用以其全部内容结合在本文中。关于联邦政府赞助的研究或开发的声明本发明是在以分别由美国国防部高级研究计划局微生理系统项目(defenseadvancedresearchprojectsagencymicrophysiologicalsystemsprogram)和美国国立卫生研究院(nationalinstitutesofhealth)授予的合同w911nf-12-2-0039和uh3tr000496为基础的政府支持下完成的。政府在本发明中具有某些权利。
背景技术:
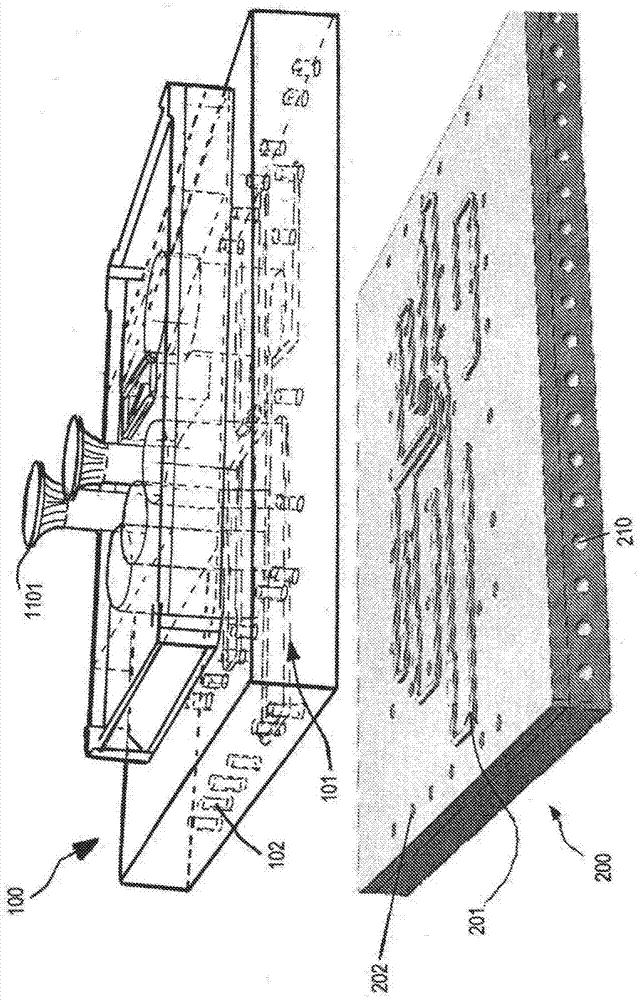
对于在临床试验中减少成本高昂的失败来说,提高对人药物反应的临床前预测的有效性至关重要。复杂疾病经常起因于由局部扰动和全身扰动的整合引起的全身性调控网络调控异常,包含跨多个器官。对组织间通信的不完全理解可能不利于对疾病状况的准确诊断和治疗。尽管对人病理生理学的研究已经依赖于如小鼠模型等基因易处理动物模型,但是这些动物模型对于概括具有各式各样临床表型的多基因和多因素人疾病来说可能并不充分。细胞生物学、微制造和微流体学的最新进展已经实现了对人器官的功能单元的微工程化模型——被称为器官芯片(organ-on-a-chip,occ)——的开发,所述微工程化模型可以以较强的预测能力提供临床前测定基础。例如,授权给griffith等人的美国专利号6,197,575描述了一种适合于复杂分层组织或器官结构的接种、附接和培养的微基质和灌注组合件。授权给inman等人的美国专利号8,318,479描述了一种促进以适合于在多井板格式中培养和测定的毛细管床的长度尺度进行的灌注的系统。被称为微生理系统(microphysiologicalsystems;mps)的这些平台被设计成通过将组织工程原理与微制造或微机械加工技术整合来模拟生理功能以便概括3d多细胞相互作用和对营养物转运和/或机械刺激的动态调控(huhd等人,《芯片实验室(labchip)》,第12卷,第12期,第2156到2164页(2012年);sungjh等人,《芯片实验室(labchip)》,第13卷,第7期,第1201到1212页(2013年);wikswojp等人,《实验生物学和医学(expbiolmed)(梅坞市)》,第239卷,第9期,第1061到1072页(2014年);livingstonca等人,《计算和结构生物技术杂志(computationalandstructuralbiotechnologyjournal)》,第14卷,第207到210页(2016年);yuj等人,《今日药物发现(drugdiscoverytoday)》,第19卷,第10期,第1587到1594页(2014年);zhul等人,《芯片实验室(labchip)》,第16卷,第20期,第3898到3908页(2016年))。虽然对单独mps(例如,心、肺、肝、脑)的开发已经取得了显著进展(rotha等人,《先进药物输送评论(advdrugdeliverrev)》),第69和70卷,第179到189页(2014年);huebschn等人,《科学报告(scientificreports)》,第6卷,第24726页(2016年);domanskyk等人,《芯片实验室(labchip)》,第10卷,第1期,第51到58页(2010年)),但是针对mps互连所做出的努力仍处于初级阶段,其中大部分研究主要集中于基本生存力和毒性展示(oleagac等人,《科学报告(scirep)》,第6卷,第20030页(2016年);eschmb等人,《芯片实验室(labchip)》,第14卷,第16期,第3081到3092页(2014年);maschmeyeri等人,《芯片实验室(labchip)》,第15卷,第12期,第2688到2699页(2015年);materneem等人,《生物技术杂志(jbiotechnol)》,第205卷,第36到46页(2015年);loskillp等人,《公共科学图书馆综合(plosone)》,第10卷,第10期,第e0139587页(2015年))。然而,缺少临床疗效而不是毒性被认为是ii阶段和iii阶段临床试验(成本最高的阶段)中药物损耗的主要原因(kubinyih等人,《自然综述:药物发现(natrevdrugdiscov)》,第2卷,第8期,第665到668页(2003年);cookd等人,《自然综述:药物发现(natrevdrugdiscov)》,第13卷,第6期,第419到431页(2014年);denayert等人,《转化医学新视野(newhorizonsintranslationalmedicine)》,第2卷,第1期,第5到11页(2014年))。主要促成因素包含不完全理解疾病机制、缺少预测性生物标记以及种间差异。由于需要用于靶识别/验证和生物标记发现的人源化模型系统,所以在药物开发时存在迫切的未满足的需要。对更具预测性的体外系统的不断增长的需要不限于单一mps技术。可以在多mps平台中在全身性层面更好地概述人体生理学的复杂性,在所述多mps平台中,多器官串扰和对治疗剂和毒素的生理反应通过替代信号(例如,趋化因子、细胞因子、生长因子)和循环细胞(例如,免疫细胞)而发生。shuler等人展示了对多隔室生物反应器系统的药理学应用(sweeneylm等人,《体外毒理学(toxicol.vitr.)》,第9卷,第307到316页(1995年))。sung等人展示可以将其中细胞被嵌入在单独室中的3d水凝胶中的微细胞培养模拟物(μcca)用于与mps系统相互作用(sungjh等人,《芯片实验室(labchip)》,第9卷,第1385页(2009年))。一些原型将重力流动用于mps间通信(sungjh等人,《芯片实验室(labchip)》,第10卷,第446到455页(2010年))。三mps系统的一些原型使用利用气泡捕捉器的平台外泵送(sungjh等人,《芯片实验室(labchip)》,第9卷,第1385页(2009年);eschmb等人,《芯片实验室(labchip)》,第14卷,第3081页(2014年))。虽然毒理学和药效学研究是常见应用,但是药代动力学研究在多mps平台方面一直受到限制。此外,当前多mps系统通常采用与用于与非常小的流体体积一起操作的传统微流体芯片相关的封闭格式(annasl,《流体力学综述年刊(annu.rev.fluidmech.)》,第48卷,第285到309页(2016年))。这些系统的当前制造工艺需要使用如pdms等可铸造弹性体聚合物以主要获得期望的光学性质,但是由于通常存在如药物和生长因子吸收等流体表面相互作用(halldorssons等人,《生物传感器与生物电子学(biosens.bioelectron.)》,第63卷,第218到231页(2015年))。硬件设计和制造方面的其它实际限制还显著降低了现有多mps装置定制化的鲁棒性、长期稳定性和相容性。不良硬件设计和构建体常常导致缺少对流体在井当中的方向性(井间方向性)和井内再循环的控制,从而由于流体流动的破裂、虹吸效应和/或蒸发而使一些井变干。以近生理尺度进行的培养基耗尽和废物移除经常需要单程培养基流动,从而使得难以或不可能研究缓慢清除药物、药物代谢物的作用以及mps间通信。在培养一些组织时,用于装配到多mps装置的井中的可移除插入物可能是令人期望的,但是一直难以实现所述插入物与流体流入的相容性以支持对培养物的灌注。因此,本发明的目的是提供具有用于长期组织培养以及对多个模块化器官模型的简单测定的整合流体控制装置的改进设备。本发明的另一目的是提供将流体泵送、调平和感测与生物反应器整合的方法。本发明的又另一目的是提供与敞开式流体生物反应器的动态调控相容的插入装置,所述插入装置支持灌注并且允许对细胞和生物材料的平台外接种、简单操纵以及容易从生物反应器移除而不引起损坏或污染。技术实现要素:多井细胞培养系统(或器官芯片装置、微生理生物反应器)设置有整合的泵送、自发流体调平以及可编程药物/培养基给药。多井培养系统、即芯片或生物反应器含有至少三层构建体,所述三层构建体从上到下为(1)多井细胞培养板构建体,其具有位于下面且连接到井的内置流体通道(例如,流体路径);(2)作为泵致动器的屏障膜;以及(3)用于呈现压力和真空的气动板。在不同实施例中,膜层结合在流体侧或气动侧上,或者是单独的部件。将膜层结合到气动侧或流体侧增强了可靠性并且减少了制造时间和成本。在优选实施例中,膜结合到气动侧,并且流体层是敞开的,从而使得更容易进行清洁和灭菌。在一些实施例中,在流体侧上无结合消除了分层。对真空或压力的气动控制使膜致动,所述膜的作用就像阀门以控制流体通道上的通过或阻塞,因此系统的流体侧上的流体流动。如细胞培养基等流体流入以填充所述井中的至少一个井,并且连接上部空间中的两个或更多个井的被动自调平溢流道允许将过多流体从一个井转移到另一个井。通过额外泵允许井内或两个井之间的主动再循环。所述系统组合以下特征中的一个或多个特征以提高所建模的器官芯片的可操作性和性能:具有用于促进单向流动和抗虹吸能力的、限定几何布置的溢流道。入口中的一个或多个特征、导管和/或溢流道的出口被设置成确保跨溢流道的自发毛细管流动以实现mps室中的流体量的单向自调平。一些实施例提供了消除用于使流体膜破坏最小化的阶梯式或v形切口的入口几何结构;并且包含源井周围的径向弯月面钉扎槽,所述槽能够“钉扎”流体弯月面,从而使得指定流体高度是大大有利的。一些实施例提供了溢流导管,其具有沿着导管的、位于底部的小宽度(例如,小于3mm)高纵横比槽,以用于允许自发毛细管流动、因此从源井到目的地井的过多流体调平。一些实施例提供了其中溢流导管末端的槽遇到放大的弯曲区域以使流体膜变薄的出口几何结构,由此将流体膜分裂成由于重力而合并且下落的小滴。在另一实施例中,在溢流道的出口处存在沿着壁且朝着目的地井的底部的垂直槽。另外,一些实施例在目的地井的壁中提供了底切,其中所述切口处于导管出口下面的某个距离处以防由于虹吸效应产生的回流。这些特征允许自调平溢流道进行单方向流动并且防止流动破裂和在源井或导管中的过度积累。任选地与内部湿度储库或抗蒸发壕沟耦合的多器官mps平台允许对功能器官样组织的长期培养,例如,持续至少1周、2周、3周、4周、5周、6周或至少1个月、2个月、3个月。板上泵送系统(例如,内置流体泵送通道)消除了对管的需要,但是模块化泵送可以被配置成驱动外部流动。可以使用套圈连接将内置泵与外部管介接,从而允许泵送歧管在紧凑包装中同时驱动大量流量。除了单个多室单元泵送系统之外的双泵送系统不仅允许脉动流动而且允许平滑的流动体积剖面。三泵送系统或更多平行通道可以进一步增加流动平滑性。提供了用于装配到平台上的井中的可移除且灌注使能的支架。与不允许可整合特征参与生物反应器中的灌注过程的常规可移除插入物不同,所述支架使得能够在平台上灌注细胞培养物并且在平台外处理细胞培养物。所述支架可以任选地含有用于非接触式氧气(o2)感测的流体聚集盖。提供了用于非接触式流体液位感测的一种或多种装置。具有对称的前和后电极设计的电容器从井壁内提供对井内流体液位的准确测量,从而避免直接接触电化学反应以及潜在污染。由于穿过生物反应器的气动板的主体的内部通道(例如,空气致动线路)的贯通式设计而可以将两个或更多个多器官生物反应器菊链式连接。还可以将两个或更多个生物反应器堆叠以节省空间。提供了堆叠配置的气动线路和流体连接布局。所述平台优选地由使由于吸附而产生的生物化学因子损失最小化的材料制造而成。在一些实施例中,顶部流体板由聚砜制造而成。在一些实施例中,顶部流体板由聚苯乙烯制造而成。在一些实施例中,顶部流体板由亚克力材料制造而成。在一些实施例中,致动膜由聚氨酯制造而成;可替代地,弹性体被分区段地置于多室泵送单元上以代替聚氨酯膜。器官芯片具有板上气动微流体泵送以便实现对如肝组织等功能组织的延伸3d培养。板上泵送技术使与过多管相关联的空间、辅助设备以及死体积最小化。多器官平台的特征在于针对0毫升每天到几百毫升每天的宽范围流速内的精确流速控制的、具有如0.1微升每行程与10微升每行程之间的受控体积通量、频率在约0.01hz与20hz之间的确定性泵送,以提供每个mps内的受控培养基再循环以及受控“全身性”循环。所述平台与具有被设计成容纳不同类型的微组织的典型多井板具有类似占用空间。单独的组织隔室配备有其自身的mps内泵以提供营养物再循环并且通过用于液位控制的被动溢流道流体连接到混合器。尽管单器官培养对于所述平台来说是可行的(例如,具有来自其它井的灌注和药物添加益处),但是硬件可以被重新配置成适应多种应用,包含双向、3向、4向和n向相互作用(n>=2),具有对混合室到不同组织的流速和流动分割的用户定义控制,从而概括生理相关循环。还提供了对多向mps相互作用的验证。“m-wmps”是指每个单独的微生理系统借此具有其自己的内部循环以独立于平台上的其它mps单元控制氧合和混合以及机械刺激的一种配置。每个mps流体通过中心循环流动回路或通过直接连接以受控方式流体连接到其它mps单元。例如,肠模块具有内部循环以混合转移井膜下面的流体并且收纳来自中心循环流动的流量,然后其流出物直接去往肝。肝模块具有其自己的内部循环流动,并且收纳来自肠、胰腺以及中心循环流动的流量。附图说明图1是具有板上泵送系统的多井装置中的部件的示例性图示。流体板100含有两个或更多个可以装配有如转移井(transwell)1101等插入物的井的在所述井中的至少两个井之间提供流体连接性的流体路径101以及用于与第二板200附接的销孔或插槽102。第二板200(例如,气动板)含有大量内部通道(即,空气致动线路),所述内部通道各自在第二板200的相反侧具有开口210(入口开口和出口开口)。第二板的表面上是与流体板100的流体路径101的形状、迂曲度和长度相对应的一个或多个凸起特征201。这些凸起特征具有与所述内部通道中的每一条内部通道连接的孔,使得压缩空气或真空通过内部通道分配到气动板的表面上的孔。气动板还具有用于与流体板附接的插槽202。不锈钢螺丝将所述层紧固成可以像传统n井板一样处理的单个单元。图2是示出了在气动板200的内部通道(即,空气致动线路)的开口210处菊链式连接的两个装置的示意图。流体板100(具有板盖1200)与气动板200组装。图3a示出了组装好的7向装置的示意图,所述装置具有用于细胞培养的井103和/或其中转移井插入物1101被装配到井中的混合培养基。与流体板的流体路径流体连接的两个端口105可以用于与外部流体容器连接以导入和/或导出流体。图3b是示出了与图3a相对应的7向平台上的待放置器官以及器官之间的流动方向性的绘图。图4是7向装置的气动板的俯视图的示意图。所述板具有用于对齐的对齐销203以及用于与流体板附接的插槽202。所述板在表面上具有凸起特征201,所述表面在多个位置中具有表示一组三室单元220a、220b和220c的一组三个孔。这三组三室单元与在气动板的相反侧具有入口开口210a和出口开口210b的三条内部通道(即,空气致动线路)气体/压力连接。这三组三室单元中的每一个三室单元的中间孔/室被定位成共享相同的内部通道(即,空气致动线路)。这三组三室单元中的每一个三室单元的中间孔/室的同一侧(即,左侧或右侧)上的孔/室被定位成共享另一相同的内部通道(即,空气致动线路),从而降低装置的气动致动流动控制的复杂性。此处在气动板上还示出了流体板的井和溢流导管121的相应位置。图5是示出了肠-肝-肺-子宫内膜4向平台的横截面侧视图的示意图。箭头表示流体流动的方向,其中流体通过井中的入口111a泵送到肠井103d中,超过某个高度的过量流体通过溢流导管121溢出到含有氧合尾部103c的肝井103b。肠井还在井中具有出口111b以便与入口111a进行可能的井内流体再循环。来自混合器/混合井103a的流体通过流体路径流到细胞培养井,包含子宫内膜井103e和肺井103f。所述板还具有用于抵抗蒸发的壕沟104。图6是示出了4向平台上以双向配置操作的每个井的流动方向性和细胞培养类型的图示。图7是示出了4向平台上以单器官配置操作的每个井的流动方向性和功能的图示。图8是示出了采用双向配置操作的每个井的流动方向性、流动分割和细胞培养类型的图示。图9是示出了另一4向平台上的每个井的流动方向性、流动分割和细胞培养类型的图示。图10是示出了7向平台上的每个井的流动方向性、流动分割和细胞培养类型的图示。图11是用于将药物添加到双向相互作用组的井定向的不同配置的图示。图12是示出了溢流道(包含溢流导管121)的俯视图的示意图,所述溢流道提供了源井103i到汇井(或目的地井)103j的单向流体连接性。还示出了源井103i的入口111a和出口111b。图13是离开源井103i(包含例如用于主动泵送诱导的再循环的出口孔111b)的溢流道的入口几何结构的侧视图。径向弯月面钉扎槽122与溢流道的弯曲入口几何结构124对齐,并且弯曲的入口几何结构与溢流导管的导管槽125的底部对齐。转移井高度通过转移井的外缘搁置于其上的阶梯状搁架123的垂直位置设定。图14是进入目的地井103j的溢流道121的出口几何结构的侧视图。溢流道的出口几何结构在目的地井的壁中在溢流导管的边缘下方包含底切130以及用于沿着目的地井的壁引导的垂直槽131。图15是装置的灌注井中的可灌注支架的横截面侧视图,所述横截面侧视图示出了支架中的顶端体积1102以及井中的基部体积1103。图16a、图16b和图16c示出了(横截面侧视图)的溢流道v形入口几何结构的连续时程潜在发展,从跨溢流道的初始连续流体膜(图16a)到流体膜破裂(图16b)以及最终汇井中的变干和源中的过多积累(图16c)。图17是不具有图16a中的针对跨溢流道的连续流体膜的v形状的另一溢流道入口几何结构的横截面侧视图的示意图。图18是与图17相对应的溢流道入口几何结构、即在底部具有槽的u形导管的放大横截面侧视图的示意图。图19是溢流道几何结构的另一实施例的横截面侧视图的示意图。此溢流道具有允许敞开式流体流动(导管126上方的空间)的带有塔状导管128a入口和向上导管出口129a的导管127。图20是图19所示溢流道的俯视图的示意图。塔状导管在源井的壁中的阶梯的表面上具有开口、即孔128b,所述开口连接到采用敞开式流体配置126的溢流导管127。图21图是图19所示的溢流道的横截面侧视图的示意图,其中螺钉140插入塔状导管128a,从而阻止从源井中溢出。图22a、图22b和图22c示出了具有(横截面侧视图)的v形入口几何结构的溢流道的连续时程发展,从初始的流体前端进入导管(图22a)到流体前端沿着导管迁移(图22b)以及导管中的流体积累(图22c)。图23是用于计算以确定自发毛细管流动(spontaneouscapillaryflow,scf)的导管几何结构尺寸的示意图。wf表示液体-空气界面的尺寸。图24是用于计算以确定scf的矩形导管尺寸的示意图。导管具有深度b和宽度a,总计横截面导管周长pw,而液体-空气界面具有周长pf。图25是不具有用于支持scf的v形入口几何结构的溢流道的横截面侧面的示意图。图26a到图26d示出了倒圆底部溢流导管的不同视图:入口处视图(图26a)、对角视图(图26b)、截面视图(图26c)以及出口处视图(图26d)。图27a到图27d示出了具有刀刃几何结构的溢流导管的不同视图:入口处视图(图27a)、对角视图(图27b)、截面视图(图27c)以及出口处视图(图27d)。图28a到图28d示出了具有v形切口几何结构的溢流导管的不同视图:入口处视图(图28a)、对角视图(图28b)、截面视图(图28c)以及出口处视图(图28d)。图29是具有高深宽比(例如,大于3)的位于底部的矩形槽的溢流导管几何结构、即u形的横截面侧视图的示意图。图30a、图30b和图30c示出了具有v形入口几何结构(横截面侧视图)的溢流道的另一连续时程发展,从跨溢流道的初始连续流体膜(图30a)到导管中的流体积累(图30b)以及虹吸效应(图30c)。图31是溢流道出口几何结构的横截面侧视图的示意图,其中溢流导管127以斜坡132结束,并且在导管下面的距离d处,在目的地井的壁中存在底切130。位于斜坡132下面且被底切130中断的垂直槽131沿着目的地井的壁存在。图32是溢流道出口几何结构的俯视图的示意图,其中流自小宽度槽127的流体遇到放大的弯曲区域132以便离开。图33是在井的底面上具有引导槽151的氧合尾部150a的俯视图的示意图。图34是连接到之字形氧合尾部150b的井103的俯视图的示意图。图35是示出了图34所示的之字形氧合尾部的用于阶段引导的几何结构特征的图示。尾部具有最大宽度w1和最小宽度w2,所述最大宽度和最小宽度分别在长度l1和l2内以交替顺序出现。角α表示渐增宽度相对于氧合尾部中的流体流动方向的方向。图36是插入到平台上的井中的可移除灌注后支架160的横截面侧视图的示意图,所述示意图示出了用于固定(例如,通过螺纹转动)支架的坡道区域159、径向密封件161a和161b(例如,o型环)、支架中的细胞培养区域162以及可用于非接触式氧感测的流体聚集盖163。图37a是示出了气动板的表面上的三室单元的俯视图的示意图。图37b是示出了与图37a相对应的三室单元的侧视图的示意图。屏障膜300将流体板(含有流体路径101)与气动板分离。气动板具有凸起特征201,所述凸起特征上的孔产生连接到气动板(在此图中未示出)的内部通道(空气致动线路)的室空间。此处,室221充当阀门,室222充当泵,并且室223充当另一阀门。图38是由四条空气致动线路控制的双重三室单元顶部的分裂后流体流动的俯视图的示意图。图39是壁内流体液位感测电容器1200的横截面侧视图示意图,所述电容器包含位于板1202(例如,多氯联苯(pcb)板)的相反侧上的前电极1201和后电极1203。图40是图39所示的壁内流体液位电容传感器的电极的俯视图示意图,所述俯视图示意图示出了前感测电极1201b连同在一侧共面的前参考电极1201a和在另一侧共面的另一前参考电极1201c以及后感测电极1203b连同在一侧共面的后参考电极1203a和在另一侧共面的另一后参考电极1203c。图41是堆叠式平台的气动线路的三个层的示意图。图42是顶板150和底板250的横截面侧视图示意图,所述顶板和底板具有支持在所述两个板之间进行烧结的几何结构。底板250具有分别带有变窄顶点252a和252b的凸起柱251a和251b以及比凸起柱低高度d2的平坦表面凸出部253。图43是由图42的顶板150和底板250烧结而来的融合后一件式构建体350的横截面侧视图示意图。图42中的凸出柱的顶点在烧结(顶板与底板之间受热强制压缩)之后已经变形为烧结后表面252c和252d并且与顶板附接。在烧结之前的底板的凸出部之间的空间已经变成流体351的空间(例如,通道)。具体实施方式i.定义可互换地使用的术语“器官芯片(ooc)”、“生物反应器”和“微生理系统(mps)”是指在体外平台上提供单个或多个器官或其它组织类型之间的相互作用的平台,所述体外平台提供对这些组织的生长的维持。术语“气动”是指使用空气压力或真空压力进行操作的系统。术语“歧管”是指供气动或流体连接的互连装置。术语“溢流道”是指源井与目的地井之间用于自动维持源井中的流体液位的流体连接系统。术语“调平”是指维持流体液位。术语“自调平”是指使用无源装置、即在没有有源装置的情况下维持液位。术语“底切”是指与悬垂特征相关联的机械细部。术语“润湿”是指在气体环境中通过液体润湿固体表面,润湿由系统的吉布斯能(gibbsenergy)决定。在气体环境中通过液体润湿固体表面在固体/液体(sl)界面与液体/气体(lg)界面从接触线散发时跨所述界面之间的液相产生平衡接触角θ。总体而言,术语“润湿”表面和“非润湿”表面分别是指θ<90°和θ>90°的情况。接触角与所涉及的界面能之间的关系通过杨氏方程γsv=γsl-γcosθ表达,其中γsv、γsl和γ分别是固体与气体之间、固体与液体之间以及液体与蒸汽之间的吉布斯界面能,并且其中最后一个量被称为表面张力。为了满足热力学平衡要求,使用蒸汽使气相饱和。术语“弯月面”是指流体与固体材料和蒸汽相的交叉点处的流体边界。术语“弯月面钉扎”在本文中是指在垂直壁中的润湿液体的液位上升到顶部边缘的情况下,具有接触角θ的润湿线的端部保持(或者“钉扎”)在井的顶部边缘处,同时在液位进一步升高期间,在井侧壁的顶部边缘处,接触角θ将从<90°上升到>90°,直到液体积累导致溢出井的边缘,由此释放接触线(“未钉扎”)。对于非润湿液体来说,在垂直井的侧面的基部边缘和顶部边缘处发生弯月面钉扎,并且在顶部边缘处,接触线处的液体定向的角度从值θ改变为值θ+90°。在wijs等人,《分离:材料、装置和过程(separations:materials,devicesandprocesses)》,第62卷,第12期,第4453到4465页(2016年)中描述了所述术语的细节。术语“毛细管长度”是指两种流体之间经受重力加速度和由于表面处的表面张力而产生的表面力两者的界面的特性长度尺度。术语“插入物”是指可以机械地装配在mps的井中的元件。术语相关区段中的“支架”是井的为组织构建体提供支撑的插入物或部件。术语“横木(whippletree)”是指用于通过连杆均匀分配力或压力的机构。如本文中所使用的,其是指从一个方向施加在中心处或附近并且分配给尖端(通常为两个尖端)的力或压力,所述尖端各自充当用于对另外的尖端进行分配的中心。术语“程序”或“软件”是指可以用于对计算机或其它处理器进行编程以实施如以上讨论的实施例的各个方面的任何类型的计算机代码或计算机可执行指令集。另外,应当理解的是,一个或多个当被执行时执行本发明的方法的计算机程序无需驻留在单个计算机或处理器上,但可以以模块化方式分布于大量不同的计算机或处理器当中以实施本发明的各个方面。ii.设备和设备操作每个多井装置总体上是具有板上泵送系统的三部件构建体。流体板100含有多个井——一些井装配有如1101(康宁(corning),也由sigma-aldrich分销)等插入物——以及用于分配培养基的内置微机械加工流体路径101(图1)。气动板200通过小孔向气动板的表面分配压缩空气和真空。屏障膜300(总体上为半透明的)位于流体板100与气动板200之间,所述膜受到压力可能挠曲以扩展或收缩,由此阻挡或清除流体板的流体路径的相应部分。此屏障膜还提供无菌屏障,所述无菌屏障充当泵和阀门的致动层。可以将多个装置链接以实现同时同相操作/致动(图2)。每个装置都是生物反应器,所述生物反应器作为平台而支持对将不同器官、其互连以及相互作用模拟为在体内的多个mps的培养。敞开式井和通道允许用户容易接近细胞和培养基以执行需要直接流体接触的测量。如实例中所展示的,已经将这些mps中的高达七个mps耦合在一起,但是应当理解的是,所述系统允许混合同一类型的mps中的多于一个mps以及混合和整合各种不同的类型,而不限于总共七个。所述系统唯一地结合了被有效地配置成支持多器官培养的高自由度(dof)板上泵送系统。虽然现有装置具有通过单个泵被线性链接以驱动流过环(materneem等人,《视频实验期刊(j.vis.exp.)》,第1到11页(2015年),doi:10.3791/52526)或与施以预定义被动流速的通道直径并联链接(oleagac.等人,《科学报告(sci.rep.)》,第6卷,第20030页(2016年))的隔室,但是高dof控制使得易于重新配置平台以添加新的mps或排除某些隔室。在4向mps生物反应器的一些实施例中,所述平台可以以18个自由度(“dof”)或18条管通道进行操作。例如,在肝-肠-肺-子宫内膜4向mps中,可单独寻址的泵需要3个dof,而多个泵可以通过跨多个泵共享气动歧管而以相同速率运行。4向mps平台可以具有用于驱动9个泵的6个可独立重新编程流速。提供混合器到mps流动的所有四个泵都可以是可单独寻址的。共享了再循环泵速:混合器/肝再循环被链接,肠/肺/子宫内膜再循环也一样。链接泵速在经济上是有利的,因为这降低了气动阀门的数量以及平台所需的管连接。在7向mps生物反应器的一些实施例中,所述平台具有操作每平台17个注射泵的功能对等物的36个dof并且可以动态控制mps内或mps间混合。在此实例中,仅可以独立指定12个流速,因为每个流速需要3条气动线。a.多井生物反应器(1)流体流动方向综述图3a示出了7器官相互作用生物反应器的示意图,针对所述生物反应器,图3b示出了将在每个井中培养的组织以及流体流动方向的示例性绘图。在含有肺、子宫内膜、肠、肝、心、中枢神经系统(cns)以及胰腺的示例性7向生物反应器中,总体上主动的流体流动通过内置流体通道从混合器井(混合器)引导到肺(图3b中的箭头1)、从混合器引导到子宫内膜(子宫内膜,图3b中的箭头3)、从混合器引导到肠(图3b中的箭头4)、从混合器引导到肝(图3b中的箭头7)、从混合器引导到胰腺(图3b中的箭头9)、从混合器引导到cns(图3b中的箭头10)、从混合器引导到心(图3b中的箭头11);并且通过井内泵送引导以在肺、子宫内膜、肠、心、cns、肝、胰腺以及混合器中的每一个内再循环(图3b中的箭头2、6和12)。可以将外部供应导入到混合器(图3b中的箭头8),所述外部供应通过流体流动分配到每个器官井。可以将来自混合器的废物导出到外部收集器(图3b中的箭头5)。在一些实施例中,从混合器到器官的每种流出物都具有指定泵以实现单独控制的流速以及将外部供应导入到混合器并将废物从混合器导出到外部收集器。在一些实施例中,为了降低复杂性,肺、子宫内膜和肠中的每一者内的再循环可以共享一种泵控制以实现完全相同的再循环流速;心、cns和胰腺中的每一者内的再循环可以共享另一种泵控制以实现完全相同的再循环流速;并且混合器内和肝内的再循环可以共享又另一种泵控制以实现完全相同的再循环流速。通常在至少一对井之间、并且在7器官平台的一个实施例中在肺与混合器之间、在子宫内膜与混合器之间、在肠与肝之间、在肝与混合器之间、在心与混合器之间、在cns与混合器之间、并且在胰腺与肝之间设计有溢流道以将过多流体自动地从前一井转移到后一井。图4示出了与图3a中示出的用于如图3b中绘出的多器官培养的示例性7向设备相对应的气动底板的示意图。气动底板可以在气动板的侧面具有用于与顶板底部上的相应特征(例如,销孔或插槽)配合/对齐的对齐销203、在一些实施例中为关于中心对称的位置处的两个销。气动板还可以在多个位置(不阻挡空气传导致动线路)处具有大量贯穿所述板的深度的孔202以供顶部流体板底部上的相应凸出销特征与其对齐。在图4所示的气动板上,存在18条内部通道作为水平横跨气动板的内部的空气传导致动线路。例如,具有空气入口210a和空气出口210b(入口和出口相对于板的定向)的一组三条空气传导致动线路控制位于气动板的致动侧(即,通过致动膜与流体模板底部组装的侧面)的表面上的多个三室单元220a、220b和220c。每个三室单元(例如,括号为220a、220b和220c)都具有三个室,所述三个室各自具有到与下面的水平空气传导线路连接的表面的空气传导孔,并且一旦顶板被组装,三个室就作为整体通过引起膜的正偏转和负偏转的气动致动来控制顶板的流体通道中的行程或蠕动流体流动。气动板还可以具有与一个或多个三室单元连接的凸起弯曲线凸起特征201。这些凸起特征为流体板的底面中的相应流体通道提供匹配的密封表面,所述相应流体通道在与各种流体mps模块互连的限定流体回路中引导流体。一旦气动板与流体板组装,这些凸起特征201就可以视为概括了流体路径在流体板中的位置。元件121显示了一旦气动板与流体板组装就在流体模板中的mps模块之间承载流体的溢流道的位置。图5示出了示例性4向平台的横截面,所述横截面示出了从混合器到肠的流体流动的内置通道以及从肠到肝的总体溢流道位置。在多器官mps平台上用于细胞培养的所公开井总体上遵循这种“流入/溢出”操作原理。流体的主动流动和被动溢出的方向的操作总体上模拟体内系统中的循环路径,并且示例性7器官生物反应器中所示的原理适用于双向、3向、4向或其它数量的mps系统的平台。通过改变用于操作的软件代码可以排除在多井平台中使用一个或多个井,并且不需要硬件改变。还可重新配置每个井以实现多种用途。例如,还可以将混合室(混合器井)用作具有免疫能力的肠mps井或可以将其与一起使用。可以将肝mps井用作培养基储库或药物储库。在图6到图10中示出了多井平台的示例性重新配置后用途。流动分割通常通过改变泵送频率来实现。在图11中示出了多井平台的另一示例性配置,其中容纳于三个井中的三种药物被递送到肝井和肠井,同时这些井通过混合器井和肝与肠之间的溢流道被灌注并且相互作用。(2)用于控制流动方向和液位的装置自调平溢流道通过切割在板的顶侧中以将过多流体递送回到混合器的溢流通道系统,所述设备被动地实现mps井的自调平以及总体上到混合器的流体返回。总体上,溢流道在底部井中的某些底部井上方包含通道(例如,敞开式流体),所述通道将入口井连接到出口井(图12)。溢流道消除了对用于实施流入物与流出物之间的平衡的返回泵和液位传感器的需要,同时还允许回流横跨入口mps给送流动。在优选实施例中,溢流道在需要调平时避免破裂溢流道中的流体流动,并且避免虹吸效应以防井变干。所述设备使用自发毛细管流动(自润湿)和阶段引导原理以引导流体通路中的流动和润湿从而允许敞开式流体器官芯片系统的更加鲁棒的操作。从源井到目的地井的单向流动是使用以下详述的弯月面控制特征以及其它特性来实现的,所述其它特性包含溢流导管的附加槽几何结构、受控表面粗糙度、表面张力以及溢流道的入口和出口中的附加特征。器官芯片设备的流体容器中的这些一个或多个几何特征允许以径向方式钉扎流体以限制通过表面张力产生的弯月面效应。这种构造可以允许更好的被动流体调平,所述流体调平然后可以在这些系统内转变成更具确定性的性能和测量。溢流道以以下方式实施被动调平。如果到入口井中的流体流动引起流体在入口井中纯粹地积累,则入口井中的液位将开始上升。随着液位开始上升,流体将在溢流道处上升,并且由此引起增加的流量流过溢流道到达出口井中。如果入口井中的液位降低,则入口井的溢流道处的流体液位将下降,由此降低通过溢流道的流量。以此方式,入口井中的液位被被动控制成大致上等于期望液位。这种调平是被动的,因为不存在感测液位并且响应于这种液位感测而改变某个泵送速率的主动过程。相反,重力和表面张力的效应组合以便以不需要明确感测和控制的被动方式调节流量。为了实现适当的溢出功能,在源中的所设计流体高度上方,溢流道在源到汇的方向上采用低阻流动路径。在一些实施例中,所述路径对于从汇到源和所述系统的流入物来说是不可渗透的,使得溢流道整体上可以抵抗由于倾斜而产生的流体高度变化。入口几何结构各种入口特征对于稳定源井弯月面来说是有用的,从而提供到溢流通道的入口或密封源井中的培养基体积的方式。图16a到图16c示出了在源井(入口井)处具有v形切口的溢流道如何经历流体膜中止(例如,流体膜破裂)并且因此溢流导管变干,从而使流体在源井和汇井中积累以变干直到变空的时程示意图。这种类型的溢流道以具有允许流体输送的相关流体剖面的亚稳方案开始操作。当流体膜破裂(具体地,在入口阶梯式和v形切口几何结构处),发现在很大程度上有利的是,流体在源井中积累由此增加高度,而不是在流体入口中前进并溢出到导管和汇井(出口井)中。当高度增加超过某个值时,流体最终溢出;但是对于如胰腺和肝等具有大表面积的器官来说,这种高度增加需要很大的体积量,已经发现,在使用这些几何结构来测试7向平台时,这是混合器在孵育器中的12小时之后变干的主要原因。已经确定了以下内容以提高效果:平坦弯月面的浅且平缓的入口到溢流道的较浅且较平缓的入口几何结构使得用于使流体溢出到导管槽中的能量最小化。源井中的径向槽引导弯月面并且利用高度增加以产生溢出事件。当存在流体膜并且溢流道正引导流体时,阶梯式特征和v形切口特征可能并不防止体积位移免于瞬时倾斜或虹吸。因此,对于一些实施例来说,消除了入口阶梯和v形切口以使在此液位下的流体膜破坏最小化。可以使用阶梯式屏障来防止另外的流体积累,如图17中使用不具有图18所示的v形切口的示例性入口的横截面视图所示。当重力占主导并且表面张力效应可忽略时,如在具有较大互连溢流道的大井中,v形切口有效地确定自调平的确切高度并且破坏连接。对于较小几何结构来说,更有效的是,具有到溢流道中的直接入口(并且在一个实施例中,具有弯月面钉扎槽)并且注意通过使用溢流道出口特征来破坏流体接触。流体钉扎槽在一些实施例中,到溢流道的入口另外包含“流体钉扎”槽,其可以是流体井中的20度、30度、40度、45度、50度或60度圆周槽122、优选地45度。这种槽捕获流体弯月面,这促进维持限定流体高度并且改进调平和溢流道操作动态。此径向弯月面钉扎槽的底部与溢流道流体流动通道的底部对齐,如图13所详述的。所钉扎弯月面不稳定并且因此将溢出,使得流体不会上升超过径向弯月面钉扎槽的高度。插入聚四氟乙烯环以实现确定性的流体液位。相对于溢流道将聚四氟乙烯环放置在不同的高度确定溢出前的最大流体高度。所插入聚四氟乙烯环捕获弯月面,因此固定液位以便不超过所述液位。所述环还有助于防止蒸发。实施例图13示出了溢流道的改进入口几何结构的一个实施例,其中浅且平缓的流体入口通过井周围的径向弯月面钉扎槽,其中弯月面钉扎槽的底部与槽形流体流动通道的底部对齐。图19和图20分别以横截面侧视图和俯视图示出了敞开式导管溢流道的改进入口几何结构的另一实施例。倾斜的导管塔128a将源井连接到敞开式导管127,所述敞开式导管可以在底部具有自发毛细管流动(scf)槽。入口几何结构利用壁中孔设计,其中孔128b产生于阶梯式表面上以连接到倾斜的导管塔128a。可以放置螺钉密封件140以插入导管128a的开口孔从而隔离mps相互作用(图21)。螺钉密封件通常在螺纹附近具有o型环以在插入到孔中后产生良好的密封。允许自发毛细管流动(scf)的导管图22a到图22c示出了来自具有v形入口几何结构的溢流道的流体跨溢流导管的时程发展。当还未对导管进行打底漆时或者当溢流导管由于蒸发或流体膜破坏而变干时,来自源井的迁移流体的前端在导管的壁内形成弯月面,所述弯月面缓慢前进并且积累流体超过导管的槽。此溢流导管问题首先发现于对7向α溢流道的染色实验中,其中溢流道通过流体前端润湿,但沿着导管的流体迁移很慢并且需要大量体积以润湿整个溢流道。以下表示了用于通过改变导管几何结构来提高流量的方式。用于允许自发毛细管流动确保通道中的鲁棒润湿的几何结构和尺寸。在相当的小尺寸圆底、v形以及矩形底部敞开式通道之间对沿着通道的流体移动效率进行了比较。在敞开式通道的一端添加了2μl的流体小滴以在不对通道打底漆的情况下测量润湿距离。显示的是,v形通道展现出103mm的润湿距离。矩形通道具有44mm的润湿距离,并且圆底通道具有7mm的润湿距离。v形通道和矩形底部通道两者支持concus-finn流动(berthierj等人,《aims生物物理学(aimsbiophysics)》,第1卷,第1期,第31到48页(2014年))。较大的润湿距离总体上表明在敞开式通道溢流道中维持连续流体流动的较好润湿性能。用于形成导管的材料的影响具有自发毛细管流动(scf)的导管在最小的体积需求的情况下并且在无任何特定打底漆或泵送速率的情况下保持流体膜以及因此流体连通。为了实现scf,导管的横截面应当满足以下关系式:其中pf=自由(与空气接触)周长pf=润湿(与壁接触)周长θ=广义卡西角(材料的平均接触角)。当通过润湿壁产生的能量降低超过扩展自由表面产生的能量增加时,产生scf。使用吉布斯热力学方程,复合壁和空气系统中的自发毛细管流动的一般标准是广义卡西角θ必须<90°。广义卡西角是材料的平均接触角。在流体板由聚砜制成的优选实施例中,对于具有水或培养基的聚砜来说,已经测得培养基-聚砜-空气的接触角为30°<θc<113°。此宽范围的接触角基于表面微拍动的效应并且在较小程度上基于很小的聚砜疏水性差异和培养环境的热效应。为了满足scf关系式,允许本文所述实施例中的scf的周长比的范围为0<pf/pw<0.866(cos30°≈0.886;负周长比是不可能的,因此未考虑)。这是示例性估计,并且应当理解的是,其它实施方式可以利用替代性比例。实际上,通道中的任何地方的接触角被合理地假设为≤80°,从而考虑弯月面效应和/或润湿性差的表面(可以对其进行机械加工以产生用于促成较高润湿性的光洁度)。因此,在具有明显弯月面效应或具有润湿性差的表面的场景中,为了使接触角为约80°,周长比变为0<pf/pw<0.18(cos80°≈0.174;arccosine0.1866≈80°)以便满足scf关系式。图23和图24提供了对任意形状的通道的横截面分析。此处,暴露于空气的液体的周长wf将是以上关系式中的自由周长pf;并且与三个壁接触的流体周长之和w1+w2+w3将是以上关系式的润湿周长pw。图24示出了具有宽度a和高度b的示例性矩形通道。为了满足scf关系式,周长比应当遵循:当限定纵横比λ=b/a因此b=λa时,所述关系式变为:可以对其进行计算以得出纵横比标准从而允许通过润湿性差的表面和/或具有明显弯月面效应的通道表面实现scf:因此,具有大于3的纵横比的小矩形通道总体上可以实现scf。在考虑制造能力的一些实施例中,纵横比范围为2.5<λ<5以支持scf设计原理。在一些实施例中,在具有约2的纵横比的三角形水平通道中实现自发毛细管流动,其中壁光滑度为使得接触角为约60o。相比于矩形通道,对三角形通道的pf/pw的计算将不同,但是相同的原理成立。在一些实施例中,溢流导管内的优选流体路径是具有大于3的纵横比的矩形或v形通道,所述纵横比处于微流体尺寸内以允许发生毛细管流动(图25示出了跨溢流道的连续流体膜)。在与导管通道进行初始流体接触时,具有支持scf的几何结构的通道中的最小流体体积将快速润湿整个几何结构并且产生能够高效地将流体从源输送到汇的流体膜。用于确保重力相关溢出的毛细管长度和溢流道宽度。根据brakke等人,《实验数学(expmath)》,第1卷,第2期,第141到165页(1992年),对于与亚克力(其具有与聚砜类似的疏水性)接触的水来说,毛细管长度[γ/(ρ*ɡ)]1/2(其中γ是表面张力;ρ是流体密度;并且ɡ是重力加速度)为2.7mm。如果通道的两个壁之间的距离(即,溢流通道的宽度)小于毛细管长度,则重力影响可忽略。因此,2.1mm的溢流道宽度使系统采用其中重力的主导地位不如毛细管的方案。在期望通过重力驱动溢出的一些实施例中(例如,在导管塔128a中),溢流道宽度大于3mm。实施例图29示出了具有用于允许自发毛细管流动的3:1纵横比矩形槽的示例性溢流导管几何结构。溢流道上方的u形通道是用于为钻头夹头流出空间的释放切口(reliefcut)。在优选实施例中,一旦源井中的液体调平,表面张力就自发传播,并且驱动流体移动穿过导管到达目标井。具有底切设计的出口几何结构图30a到图30c示出了来自具有v形入口几何结构并且不具有额外出口几何结构的溢流道的溢流道出口。当溢流道出口在垂直壁中不具有流体膜时,流体开始在导管中积累并且在出口几何结构处导致溢出爆发或甚至是稳定的弯月面。当导管处的流体弯月面与汇处的弯月面接触时,这种积累停止,并且流体膜被重新建立。当流体膜总是存在时,甚至是在源流体液位低于汇液位之后,不良出口设计可能遇到虹吸效应。可以使用以下选项中的一个或多个来避免或最小化这种问题:用于防止逆流的沿着高纵横比垂直槽的尖锐底切。图31示出了溢流导管127通过略微成锥形的浅斜坡(边缘)132离开以与沿着汇/目的地井的壁的垂直槽131连接。例如使用铣削机械形成的尖锐底切130将垂直槽131分裂成两部分。底切被切割到汇井的壁中在锥形浅斜坡132下面并且具有与垂线具有大于约30°的角度(例如,30°、35°、40°、45°、50°、55°、60°或更多,以及示例性数量之间的任何连续角度)。在一些实施例中,底切130与溢流导管出口之间的距离d处于溢流导管的宽度的约5倍到约10倍之间,以便防止虹吸效应。垂直槽131被设计成展现自发毛细管流动(scf)并且维持流体膜。除了存在底切的地方,垂直槽从上到下连续行进。只要存在前向流体方向性,这种几何结构就有助于维持连接导管和汇的稳定流体膜。在反向流动(例如,虹吸效应)的情况下,底切切断流体膜并且产生流体弯月面,所述流体弯月面将仅在重新建立前向流动时重新连接流体膜。在一些实施例中,使用导管中的scf槽中描述的相同设计参数、例如大于3的高纵横比将溢流道出口垂直槽配置成展现自发毛细管流动(scp)。已经在3x3α溢流道和机械加工的聚砜块中、在一系列实验中对底切和高纵横比垂直槽进行了测试,其产生受控的流体膜破裂和抗虹吸效应。改进的出口几何结构上的稳定垂直流体膜不会容易蒸发并且在重新获得溢流道出口处的前向流动时允许流体膜恢复和流动。用于将膜分裂成小滴的倒圆斜坡出口和小宽度槽。另一改进特征是在溢流导管的末端引入倒圆斜坡出口/边缘。当溢流导管的小宽度scf槽“遇到”放大的圆形弯曲区域(图32)时,小宽度scf槽中的稳定流体膜(由于表面张力)在放大的圆形弯曲出口区域处变得不稳定,所述稳定流体膜被有效地分裂成小滴并且将落(“掉”)到汇井中。以此方式,源井变得与汇井独立,并且实现流体流动的单向性。在一些实施例中,从源井到导管的入口几何结构不具有斜坡,即,其从尖锐边缘下降,而来自导管的出口几何结构在液体滴入汇井之前遇到放大的弯曲区域。来自导管的替代性向上出口在scf通道处于汇井的期望液位下面的一些实施例中,利用了具有出口孔的向上出口导管,如通过图19的元件129a所示。实施例图31示出了在出口之下具有底切的溢流道出口以及用于抗虹吸效应的垂直槽。钉扎在平面壁的边缘上的壁结合液滴总体上被称为壁边缘结合液滴。壁边缘结合液滴在大自然中通常作为露水而存在,露水悬挂于植物叶子上,直到达到相当大的体积并且液滴落下。当液滴被钉扎在有尖头的壁边缘上时,所述液滴被称为壁边缘顶点结合液滴。壁边缘顶点结合液滴模拟表明与高度润湿的固体壁(形成溢流道出口)接触的液体界面往往随着角度减小而滴下。这是因为由于润湿壁而引起的能量降低大于液体-气体界面的能量,使得接触区域想要在形成细流体丝的具有较小角度的角落中无限膨胀。在对于敞开式通道流体系统需要对流体调平和流动的实际控制的情形下,产生细流体丝是相关且令人期望的,因为这些细丝的亚稳定性可以提供允许或停止流体输送的手段。(3)再循环被动自调平可以促进回流,如以上详细描述的。通常,再循环用于确保浓度在井内良好分布且均匀。因此,再循环流速通常高于器官间流速。通过井内泵送驱动的主动再循环可以增加培养基的氧合。例如,在7向mps平台中,再循环可以在肺、子宫内膜、肠、心、cns、肝、胰腺和混合器中的每一个内发生。在一些实施例中,为了降低复杂性,肺、子宫内膜和肠中的每一者内的再循环可以共享一种泵控制以实现完全相同的再循环流速;心、cns和胰腺中的每一者内的再循环可以共享另一种泵控制以实现完全相同的再循环流速;并且混合器内和肝内的再循环可以共享又另一种泵控制以实现完全相同的再循环流速。(4)用于促进氧合的特征到通常含有成百上千到几百万个细胞的“介观尺度”组织的充分灌注速率对于细胞生存力来说是困难且关键的。基于具有高需氧量的肝的耗氧速率,使用细胞培养基作为循环流体,每百万个细胞需要约每秒6μl与每秒10μl之间的流速(powersmj等人,《生物技术与生物工程(biotechnolbioeng)》,第78卷,第257到269页(2002年);domanskyk等人,《芯片实验室(labonachip)》,第10卷,第51到58页(2010年);ebrahimkhanimr等人,《先进药物递送评论(advanceddrugdeliveryreviews)》,4月,第132到157页(2014年))。因为在所公开设备的敞开式流体系统中的空气-液体界面处发生气体交换,所以尽管平台材料本身是任选的,但是其无需是可透氧的。氧合尾部优选地,除了用于细胞培养的主井之外,设计了用于如肝等为了存活而需要较高氧合水平的器官的尾部。氧合尾部具有支持将氧较好地扩散和混合到培养基中的特征,如浅壁、较快再循环以及独立流入线路和流出线路。氧合尾部的示例性布局包含引导槽尾部(图33)、垂直倒圆(例如,且加深的)尾部、具有钉扎柱的平坦尾部以及具有弯月面钉扎槽尾部的平坦尾部。优选地,尾部包含倾斜表面,使得液体的深度可以如5mm、4mm、3mm、2mm或1mm一样薄以实现充分曝气/氧合。在优选实施例中,所述设备支持长达一个月、两个月或更长时间的细胞培养存活。除了尾部表面上的图案的表面粗糙度和几何结构之外,可以修改尾部的迂曲度以及尾部的宽度以增强氧合。例如,之字形迂曲尾部提供用于增强肝模块的需要减少的液体体积的氧合的手段。每个转弯环或转弯点是弯月面可以钉扎到的地方。图34示出了肝模块的之字形尾部布局。约225mm的总尾部长度可以支持约80μl的总尾部体积以实现增强的被动氧合,即增加暴露于空气的液体的表面积。图35示出了沿着氧合尾部重复存在的阶段引导几何结构。在这个之字形尾部内,此尾部以重复的方式具有分别为w1和w2的交替最大宽度和最小宽度(w1>w2)。一般而言,在尾部的处于两个u形环之间的每个段内,存在交替的最大宽度和最小宽度的两次、三次、四次或更多次重复。交替宽度各自具有其自身长度,例如,每个最大宽度w1具有l1的长度,并且每个最小宽度w2具有l2的长度。一般而言,l1大于l2以容纳更多体积。阶段引导特征归因于宽度的角度增大。如图35所示,角度α表示尾部宽度相对于尾部中的前向流体流动方向增大。尾部宽度的角度增大、即尾部宽度在前向流体流动方向上变窄提供对流体方向性的更好引导。w1可以处于0.1mm与10mm之间的范围内,例如处于约0.5mm与约1mm之间。w2可以处于0.05mm与5mm之间的范围内,例如处于约0.3mm与0.5mm之间。l1可以处于0.5mm与10mm之间的范围内。l2可以处于0.1mm与10mm之间的范围内。角度α可以处于90°与180°之间的范围内。在一个实施例中,w1为0.8mm;w2为0.45mm;l1为1mm;l2为0.5mm;并且α为150°。之字形氧合尾部的深度可以是处于约0.05mm与5cm之间的范围内的固定深度或渐变深度,例如处于约0.1mm与约10mm之间。在一个实施例中,尾部的深度被固定为0.5mm。主动氧合泵送系统用于增强氧合的另一手段是在肝井与尾部之间双向利用主动氧合泵送系统。在一些实施例中,肝培养井在其自身内具有再循环泵送系统,使得其具有从下到上的氧合后培养基流动。通常含有浅深度的液体的氧合尾部在其自身内再循环,使得在稳定状态下达到所需氧浓度。主动泵送允许将来自具有支架的井的培养基(氧浓度通常由于代谢消耗而很低)泵送到富氧尾部部分。然后,来自尾部的氧合后培养基被泵送回到所述井。(5)可移除插入物可整合用于灌注的可移除支架可以将可移除支架用于所选mps、例如肝和胰腺,从而允许对所灌注组织进行平台外接种、操纵和测定。在不引起损坏和污染的情况下,由其他人提出的之前支架难以从平台移除。在一些实施例中,可移除支架固持标准尺寸的、例如与所公开平台和/或可商购获得的相容的过滤器和保持环。支架被配置成允许通过沿着倾斜的引导坡道旋转和滑动实现平缓插入和移除。被设计成与这些支架一起使用的一些mps隔室包含倾斜坡道以引导支架插入和/或移除。所述平台的径向密封件建立有低结合性o型环(例如,o型环),从而允许对整个可移除装置进行灌注。图36示出了允许从周围平台平缓移除所灌注细胞构建体的示例性模块化可移除灌注支架。所述装置包含杯状壳体,所述杯状壳体具有径向o型环密封件161a/161b以及其基部处的流动扩散支撑结构。具有或不具有生物材料的细胞可以在此支撑件162的顶部生长。在所述装置的位于支架的上部主体或延伸臂中的侧面,两个孔允许使用无菌镊子进行操纵并且小凸缘有助于沿着坡道螺纹对其进行引导。坡道159允许通过旋转而不是垂直力进行平缓插入和移除。将支架扭转到位使细胞在插入和移除期间经历的流体压力最小化。便携性允许改进多井生物反应器的可用性的多种功能。例如,可以在唯一的细胞培养基中对构建体进行隔离培养,并且然后在所述构建体被结合在一起以进行人芯片实验之前针对健康和生存力对其进行选择。可移除支架还允许将一个细胞群体与多井生物反应器完全分离,从而允许在共享培养基不被潜在有害的试剂污染的情况下执行外部细胞健康和代谢测定。支架从下面、即井的底面支撑流体流入以及到平台上的其它井的溢流道流出。在一些实施例中,流体聚集装置163被任选地添加在可移除的灌注后支架上以将流体收集到狭孔中。流体被混合,并且所聚集流体收集通过固定位置。在流出位置处,可以通过搁置在装置顶部之内的小探针探询氧张力或其它流体性质。以此方式,用于平均o2测量的传感器不需要浸入到培养基或培养物的一部分中。在一些实施例中,利用位于膜上的具有约0.05mm与约5mm之间、优选地约0.1mm与约1mm之间或约0.25mm的薄底部/壁厚的薄支架来接种与肝相关联的细胞以增强氧合,其中支架被穿孔有大量通道(例如,~0.3mm的直径)并且保持处于再循环流动多井板生物反应器中。接种到支架中的肝细胞形成3d组织状结构,所述结构以足以在没有过度剪切的情况下跨支架产生生理氧张力下降的流速进行灌注(yatesc等人,《癌症研究进展(adv.cancerres.)》,第97卷,第225到246页(2007年))并且可以在无菌培养基中保持处于功能状态持续几周。所述设备可以含有相容的井以容纳用于细胞培养的多种插入物载剂,如用于支撑细胞或类器官的可商购获得的插入物或定制生物材料支架。(6)用于减少蒸发的壕沟另外地或可替代地,所述设备的一些实施例包含湿度壕沟(图5中的元件104)以增加局部湿度并减少细胞培养基的蒸发。可以将壕沟连接到外部流体源或通过流体板中的内置流体通道泵入的流体。可能需要对进入壕沟的流体的监测和泵送以补偿由于蒸发产生的流体损失,这通常取决于器官培养井中的流量变化。可以将所述平台内壕沟或微蒸发室置于流体板的任何区域中以增大壕沟面积从而使井的蒸发最小化,进而允许在微生理井区域周围产生湿润微环境。还可以使用壕沟中的局部加热,以便用于在平台上方维持高相对湿度的蒸发的大部分来自于壕沟。(7)用于添加或取出药剂/样本的装置可以将所述设备连接到一个或多个自采样装置或将所述设备与一个或多个自采样装置一起使用。例如,可以将自采样装置流体连接到低润湿样本收集管。(8)气动致动板上泵送相比于商业注射器或蠕动泵显著节省空间和成本、更具可扩展性、并且允许以非常低的死体积进行闭环操作。对流速和方向性的动态控制能够实现对浓度剖面的精确调控,从而允许将实验操作调整成与临床/生理分布相匹配。通过在单独的微生理系统中强加特定泵送频率来控制流量分割,从而导致特定流速以及因此对流量的“分割”。气动歧管/板流体板中的气动控制的流体流动总体上通过三室单元、例如图4的220a、220b或220c实现。图37a和图37b示出了含有位于中心的泵以及各自位于侧面的两个阀门的示例性三室单元的细节。当被顺序地致动时,此阀门安排可以通过在允许前向流体位移的同时防止回流而提供流动方向性。良好表征的可靠阀门-泵-阀门单元提供固定的流体行程,所述固定的流体行程产生确定性的流体流动。这支持约1μl/天与约10ml/分钟之间的宽动态泵送范围。在一些实施例中,泵送通道中的一个或多个或所有通道具有可逆流动,从而支持打底漆、采样和/或培养基/药物递送配置。一般而言,气动层使用贯通设计,其中具有空气入口210a和空气出口210b(入口和出口相对于板的定向)的空气传导致动线路水平穿过气动板、优选地以直线路径。空气传导致动线路的直线路径占据较少的总平台占用空间,并且所述线路支持较快气动响应(例如,由于低体积产生的快速压力变化)。对称的空气入口和空气出口允许将平台菊链式连接成同时行进,从而使用短长度的管将一个板的出口直接连接到下一个。气动歧管总体上采用由允许产生内部气动通道的材料组成的单个结合层。气动致动器膜总体上是可以按压在气动板与流体板之间或者附接到所述板之一的单层聚合材料,例如聚氨酯。在这种情况下,流体侧含有具有允许流体引导和排出的微米范围分辨率几何结构的流体通道。流体通道的分辨率较高通常产生较慢的流体移动速度,但是其可能允许较小的死体积。4路线双通道泵除了阀门-泵-阀门(v-p-v)气动致动配置之外,可以添加泵-泵-泵(p-p-p)配置以允许流体蠕动。所述三室单元中的两组或三组可以共享一条或多条空气传导致动线路,如图38所示。当(流体板的)流体通道分裂成通过被放置成相隔一条致动线且180°异相的v-p-v泵组和p-p-p泵组两者气动调节的两个通道时,从这两个脉动行程组合而来的全体流体具有平滑的体积剖面。这两个泵组的四条致动线路具有四个自由度,相比v-p-v配置,这仅需要又一条气动线路。可以以与图38所示的原理类似的原理将一个或多个x室单元(x>=3)放置成相隔一条或多条空气传导致动线路以具有定制化的流量平滑度。模块化泵送独立泵送允许实现与共享泵提供的流速不同的流速,例如比所述流速更高的流速。将流体井结合到板中可以减少或消除对管的需要,但是也可以使泵设计适合于驱动外部流动。可以使用如套圈连接等连接将内置泵与外部管介接,从而允许泵送歧管同时且在紧凑包装中驱动大量流量。用于单程或再循环灌注的泵座顶部或流体层含有mps隔室以及将所述隔室互连的泵和通道。在流体层下面,如聚氨酯膜等薄膜为通道提供密封表面并且充当泵的致动层。底层是歧管(例如,亚克力歧管),所述歧管通过将压缩空气路由到每个泵室的基部提供对泵的气动致动。当施加真空时,膜被朝着气动层下拉,从而使泵填充有流体。相反,当施加压力时,膜被迫上升到流体板中,从而将流体驱动出泵。通过连续致动三个室,固定的位移蠕动泵形成,从而允许线性地且抵抗压头而移动流体,而不产生回流(domanskyk等人,《芯片实验室(labchip)》,第10卷,第1期,第51到58页(2010年);walkeri等人,《微型机械学与微型工程杂志(journalofmicromechanicsandmicroengineering)》,第17期,第5卷,第891页(2007年))。用于降低膜应力的几何结构可以使用除了图37所示的几何结构之外的不同泵几何结构。可替代形式包含这样的设计:将泵连接到阀门的水平通道已经移除,仅留下直接链接两个相邻室的v形连接。这些v形几何结构背后的基本原理是:这些特征在膜偏转时将一个室与另一室气动隔离,使得当一个阀门被致动时,其相邻阀门不响应。泵几何结构的可替代配置可以降低膜应力并且增加致动系统的寿命及其一致性。在一些实施例中,产生对泵腔几何结构的另外修改以在膜与不同的v形桥接件之间形成一个凹形触点和一个凸形触点,从而避免膜变形和破裂。泵送验证预期流速与实际流速之间的对等能够实现平台上的mps之间的充分混合且预期的分子生物分布。对硬件的验证可以包含使用毛细管流动测量工具进行的直接泵速测量。在一些实施例中,所述工具与每个mps隔室中的出口介接,使得可以根据填充固定长度的管所需的时间来测量流量。流体板之间的流速偏差可以归因于泵室的深度的轻微机械加工差异。然而,可以输入根据所述测量计算的软件校准系数以将泵送速率校正为处于目标流速的约±5%、±4%、±3%、±2%或±1%内从而对单独泵进行调整。一般而言,小误差范围仍允许可靠且确定性的操作以及准确的数据解释。(9)用于非接触式流体液位感测的装置使用三电极设计的电容式感测可以使用电容感测、以非接触方式测量mps井中的流体液位。电荷通过塑料,使得可以将探头放置在井附近而不与井的培养基/培养物接触以避免可能的污染。可以将电容式感测探头嵌入井(例如,由塑料制成)的壁材料中。电路在无流体接触的情况下感测通过壁的电容。电容式感测电极坐于与流体隔离的塑料层后面。如图39所示,前电极1201测量流体附近的电容,而后电极1203仅测量塑料的电容(作为参考)。可以将前电极和后电极建立在多氯联苯(pcb)板1202或挠性背撑件的两侧并且因此由所述pcb板或挠性背撑件背撑。前电极在中间具有一个感测电极和两个参考电极,一侧一个,所述参考电极与中心感测电极共面(图40)。参考电极和感测电极的这种组织允许良好匹配。镜面相反电极提供自保护。之前的设计将一个负(参考电极)导电板与一个正(感测电极)并排且共面放置,以试图以非接触方式从井壁内测量液位。这使参考电容器cref处于错误的位置并且导致不准确的流体液位测量结果。所述设备利用改进的设计,所述设计含有三个对称电极、即两个参考电极与中心感测电极共面且关于中心感测电极对称,所述三个对称电极与pcb板的背面上的一组镜面电极耦合。这引起cref处于正确的位置以进行自保护和更好的匹配。光学测量对流体表面的光照明可以指示井中的液体深度。压力感测在一些实施方式中,可能有利的是,使用压力传感器确定井中的流体高度。考虑到通过p=rho*g*h给出的压力与高度之间的已知关系,这是可能的,其中p是压力,rho是流体密度,g是重力加速度,并且h是流体高度。可以将所属领域中的已知类型的压力传感器结合成与井流体连接,使得压力传感器测量已知参考高度下的井中压力。针对体积和流速对泵进行的反馈控制可以将井中的流体液位测量结果传输到对泵送到一个或多个mps井中的流体的流速进行调节的控制单元。使用井中的流体高度的电容式感测测量结果,可以使用井的已知表面积计算所述井中的流体体积。因此,对流体液位的实时测量提供体积流速的信息(即,在时间段内的流体高度差)。反馈促进对设定点体积和到mps井的泵送流速的控制。(10)用于温度和压力感测和控制的装置温度是通过将这些平台置于孵育器中来控制的,所述孵育器使全局温度保持在37±0.5度内。可以使用所属领域中已知的静态压力感测传感器类型中的任何类型进行压力感测,并且压力感测可以给出对井中流体高度的指示。结合传感器以测量井流体高度、在反馈回路中使用电容式流体液位传感器或压力传感器可以有助于主动控制井流体体积。(11)对整合部件的组装将气动侧与流体侧固定在一些实施例中,可以将通过对齐销进行的螺栓连接用作用于将生物反应器的气动侧与流体侧组装的手段。不充分的密封可能导致流体泄露。可以将夹紧用作用于将生物反应器的气动侧与流体侧固定的替代性手段。可以使用机械夹具以及磁性夹具将流体板、致动膜以及气动板夹紧在一起。可以利用横木压力分配机构跨不同的空气致动线路或跨平台分配压力。多个生物反应器的菊链式连接在一些实施例中,两个或更多个多器官mps平台在空气入口和出口处相继链接。在跨气动板的空气传导致动线路的贯通式直线路径设计的情况下,两个或更多个平台共享气动件和同一组控制器。不需要额外硬件以放大群组中的平台数量。在每个生物反应器上的空气入口和出口对称的情况下,易于设置和拆卸菊链式连接。同时,这种特征节省操作若干生物反应器/mps平台的时间、成本和空间。多层器官芯片流体板和气动板图41示出了多层堆叠平台配置的泵送系统的交叉设计。可以通过借助于对独立层或3d打印的结合形成的内部通道组装多层器官芯片板。对于这种较高密度的流体板来说,泵之间的连接可以重叠。其还与不同的泵送和阀控配置(例如,如气动致动中描述的配置)相容。具有多层板的能力能够实现代替阀门中的v形切口的内部通道,这降低由于阀门操作产生的压力尖峰并且提高性能以实现更具确定性的泵送剖面。多层板相比于单层夹紧板可以具有若干益处。较高泵和通道密度总体上允许较好的密封、总体装置占用空间减少、无需清洁一次性歧管以及使用交叉通道进行多重控制的能力增加。而且,相比于单层链配置,在多层板配置中,更容易将通道转移到期望进行成像的区域或需要插入传感器以进行测量的区域周围。其提供更自由的培养井布局以及结合新泵配置的能力。外部连接可以将多层生物反应器设备连接到微控制器和外部气动螺线管歧管以从外部提供泵送源。对于生物反应器设备的孵育来说,连接有气动螺线管歧管,所述气动螺线管歧管控制行进穿过孵育器的背部到达中间连接器的36个或定制数量的管通道。在孵育器之内,管通过阀控分离式耦合件附接到平台/生物反应器以允许针对培养基变化和采样而容易地从孵育器移除。连接器和软件架构允许所述装置在对软件配置进行最少修改的情况下与双向、3向、4向、5向、6向、7向或可定制数量的多器官平台以及许多未来的平台变体相容。泵流速和校准系数是通过膝上型计算机上的图形用户界面来设置的并且可以通过usb或wifi发送到定制微控制器(例如,美国国家仪器公司(nationalinstruments)myrio-1900)。可根据实验需要获得对泵速的手动控制和预编程控制两者,并且微控制器可以独立于膝上型计算机运行。在一些实施例中,多器官mps平台连接到控制器与泵之间的局部储库。在其它实施例中,其连接到外部微流体装置以导入外部供应和导出废物。计算机化操作控制软件被配置成实例化/安装在如微控制器(例如,美国国家仪器公司myrio微控制器)等适当装置上或与其一起实例化/安装以允许在不需要专用膝上型计算机或台式计算机的情况下连续操作生理模拟平台,但是应当理解的是,一些实施例可以利用这种专用计算机。软件为了以下目的而操作容纳在平台中的气动泵:流体补给和混合(这向细胞提供营养物和氧气);从内部源或外部源引入新鲜培养基(给送);将培养基移除到内部收集容器或外部收集容器(采样和废物收集);和/或给予在实验上感兴趣的测试化合物、生长因子、或其它化学物质/蛋白质(给药)。通过提供图形用户界面以控制混合、给送、采样和给药,此软件促进执行意在复制隔室化器官的生理相互作用的复杂实验。所述组件和/或软件还提供来自整合到硬件中的压力和真空传感器的实时反馈,所述硬件可以含有微控制器、气动螺线管、压力传感器和/或电力分配电子装置。在一些实施例中,还存在添加针对可接受范围之外的压力和真空漂移的警报的能力,并且可以实施对这些值的长期数据记录。在一些实施例中,单独和全局校准系数结合在软件中以允许针对平台上的泵的制造变化性而进行校准。例如,如果以同一频率(例如,2hz)操作的两个泵中的一个泵被机械加工成稍微深于另一个泵,则所述两个泵将不总是以恰好相同的速率进行泵送。通过确定校准系数(迭代地和/或通过实验),可以将泵的速率调整为非常精确,其中泵在2hz下被测量、校准和重新测量为目标1μl/s。软件校准系数提高泵的性能并且使跨平台的制造变化最小化。在一些实施例中,微控制器是wifi兼容的。所述软件可以配置有webui和后台(例如,labview后台)以控制泵。这允许用户在不必物理连接到微控制器的情况下在web浏览器中访问软件的控制面板,从而允许远程地控制和监测来自整个房间或整个世界的实验。从用户到输出端的示例性信息流包含以下内容。用户通过局部网络或远程地通过vpn访问webui。控制变化从webui传递到后台,所述后台调整螺线管致动定时以满足期望流速(说明单独和全局泵校准系数)。然后,微控制器将3v数字接通/断开信号输出到控制板,所述控制板将所述信号放大为对期望螺线管的12v模拟致动。在一些实施例中,为了找到故障螺线管或手动打开/关闭平台的单独泵和阀门的目的,存在允许对每一个螺线管的手动操作的调试模式。根据不同平台硬件设计,软件以大量不同方式实施,所述方式包含:4向平台软件——控制泵送和校准系数、显示4器官平台的压力/真空数据;7向平台软件——控制泵送和校准系数、显示7器官平台的压力/真空数据,这是用于限定随着时间的自动化流速变化以及来自平台上的外部端口的自动化给送和采样控制的程序模式;3xgl平台软件——控制泵送和校准系数、显示3器官平台的压力/真空数据、包含用于限定随着时间的自动化速率改变的程序模式、控制到器官的自动化给送和药物给药(受控体积)。而且,计算机可以具有一个或多个输入和输出装置,包含本文所公开的一个或多个显示器。除了其它用途之外,这些装置可以用于呈现用户接口。可以用于提供用户界面的输出装置的实例包含用于对输出进行视觉呈现的打印机或显示屏,以及扬声器或用于对输出进行听觉呈现的其它声音生成装置。可以用于用户界面的输入装置的实例包含键盘和指示装置,如鼠标、触摸板及数字化平板。作为另一实例,计算机可以通过语音识别或以其它可听格式接收输入信息。这种计算机可以通过一个或多个网络以任何适当形式互连,所述网络包含局域网或广域网,如企业网络和智能网(in)或互联网。这种网络可以基于任何适当的技术并且可以根据任何适当协议操作,并且可以包含无线网、有线网或光纤网。本文概述的各种方法或过程可以被编码为可在采用各种操作系统或平台中的任何一个操作系统或平台的一个或多个处理器上执行的软件。另外,这种软件可以使用大量适当的编程语言和/或编程或脚本编写工具中的任何一种来编写,并且还可以被编译为在框架或虚拟机上执行的可执行机器语言代码或中间代码。在一些实施例中,用于操作多mps平台的所公开软件被具体化为使用一个或多个程序编码的计算机可读存储介质(或多个计算机可读存储介质)(例如,计算机存储器、一个或多个软盘、致密盘、光盘、磁带、闪存、现场可编程门阵列或其它半导体装置中的电路配置、或其它非暂时性介质或有形计算机存储介质),所述一个或多个程序当在一个或多个计算机或其它处理器上执行时执行实施以上讨论的本发明的各个实施例的方法。所述一个或多个计算机可读介质可以是可运输的,使得存储在其上的一个或多个程序可以加载到一个或多个不同的计算机或其它处理器上以实施如以上讨论的各个方面。计算机可执行指令可以采用由一个或多个计算机或其它装置执行的许多形式,如程序模块。通常,程序模块包含执行特定任务或实施特定抽象数据类型的例程、程序、对象、组件、数据结构等。通常,在各个实施例中,程序模块的功能可以根据期望而组合或分布。而且,数据结构可以以任何适当形式存储在计算机可读介质中。为了简化说明,可以将数据结构示出为具有通过在数据结构中的位置相关的字段。这种关系同样可以通过向在计算机可读介质中具有传达字段之间的关系的位置的字段分配存储装置来实现。然而,可以使用任何适当机制来建立数据结构的字段中的信息之间的关系,包含通过使用指针、标签或建立数据元素之间的关系的其它机制。作为所述方法的一部分而执行的动作可以以任何适当方式排序。因此,可以构造这样的实施例,在所述实施例中,动作以与所示出的顺序不同的顺序执行,其可以包含同时执行一些动作,即使所述动作在说明性实施例中被示出为连续动作。iii.设备制造以上所述设备可以通过模制过程、机械加工过程和灭菌过程制造。单片式表面微机械加工流体板是优选的。其提供可靠的性能,并且其易于清洁。所有流体接触表面都是可接近的,以便进行清洁。所有部件具有相对长的寿命,并且在如高压灭菌等灭菌过程中不发生分层。气动件可以容易脱离凝结。总体上,所述设备仅使用结合在一起的两个部件,使得所有启动通道占据板内的同一平面。可以通过使入口的通道交错并且使用钻孔特征将不通过垂直位置处的入口连接到通道层而堆叠入口,因而将所述入口更浓密地压紧在歧管的侧面。模块化计算机辅助设计(cad)和机械加工的转变周期相对较快。根据研究者的个人需要,其是容易且可快速定制的。a.材料器官芯片系统可以由聚二甲基硅氧烷(pdms)、聚砜和其它材料制作而成。pdms是易于模制(并且因此高度适合于原型制作)、具有良好光学性质并且可透氧的多用途弹性体。在一些实施例中,包含类固醇激素和许多药物的疏水化合物展现出强烈分割成pdms,由此妨碍定量分析和对药物暴露的控制(toepkemw等人,《芯片实验室(labchip)》,第6卷,第1484到1486页(2006年))。在优选实施例中,流体板由聚砜(psf)制作而成。psf是具有食品级fda批准(21cfr177.1655)和uspvi类生物相容性的刚性琥珀色可机械加工的热塑性塑料。其总体上耐受广泛范围的化学溶剂,可以经受高压灭菌,并且常用于器械和医疗装置。psf还对疏水性和亲脂性化合物具有显著较低的表面吸附性以及几乎不具有大量吸收性(ngsf等人,《药学(pharmaceutics)》,第2卷,第209到223页(2010年))。可以在每次实验之前使用血清白蛋白使所公开设备的所有流体表面钝化以进一步减少对平台中的生物分子或药物的结合。还可以对流体板进行清洁并且重复使用所需次数。顶部流体板可以由整块所选材料、例如聚砜(psf)塑料机械加工成包含用于容纳每个mps的隔室以及用于整合和混合表示体循环的回流的互连室(被称为混合器或混合室)。微流体通道和泵被机械加工到流体板的底面以将流体从混合室输送到每个mps。可单独寻址的微泵被制造成与内置流体通道成一条线并且可以基于3室蠕动泵-泵-泵设计或阀门-泵-阀门设计。每个井之下的额外泵提供再循环流动,从而减少每个隔室内的营养物和氧气梯度。b.用于组装和结合的技术通常将流体板、气动板和膜(灭菌袋中)组装在生物安全柜中。在组装之前,通常将无菌微板胶带连接到流体板上以保护细胞培养区域的无菌性。然后,可以将所述层倒置组装以辅助通过亚克力板进行视觉对齐。一旦对齐销与流体板配合,就可以小心地将平台从罩盖移除,从而保持用于维持板之间的密封的压力。只要板未分离,就可以将螺钉插入非无菌环境中并将其拧紧。可以通过使用直接连接到相应端口的短长度管来连接两个完全组装的平台而将所述平台菊链式连接。菊链式连接的平台最容易使用大型金属托盘运输。平台在兴趣细胞培养实验开始之前几天(例如,4天)被组装。可以按照适合于每个隔室的体积在pbs中使用1%bsa和盘尼西林-链霉素而进行对无菌平台的表面钝化(打底漆)。通常可以通过从混合器到每个干隔室进行泵送、然后通过逆向运行再循环泵以将所有空气从通道中清除来在视觉上确认泵功能和管连接。可以使用小体积手动润湿溢流道以确保溢流道操作。平台通常在孵育器中过夜运行以在添加细胞之前钝化和确认完整操作。在一些实施例中,在特定有尖性的塑料板之间使用烧结方法来结合流体板以产生封闭流体路径。图42和图43示出了用于结合多个部件的烧结方法,其中底板250具有与顶板150的一个或多个小(例如,有尖)接触区域251a/251b以及高度比小接触区域251a/251b的顶点矮d2差异的一些平坦表面区域253。在用于将顶板与底板烧结的力和处理之后,之前的小表面区域变形并且与顶板的平坦底面融合,到达通过所接触平面表面253的接点高度设置的高度,从而产生具有供流体通过的内部空间/体积351的融合部件350。在一些实施例中,可以在张力环上展放约20微米与200微米厚之间、优选地50微米与100微米厚之间、如50微米厚的聚氨酯(pu)膜以保持恒定张力。如果存在用于将顶板与气动板对齐的螺孔,则可以将所述膜激光切割成具有气动板上的相应模式的螺孔。可以在气动板与用于流体培养的板之间展放膜隔片(任选地在与气动件的泵与阀门相对应的区域中含有弹性体)并且可以按压所述膜隔片以粘附到气动板。在一些实施例中,使用自动化将膜附接到流体板。可替代地,在膜层上使用弹性体补片以在流体板中产生密封和气密通路。可以仅在膜或补片的与气动泵和阀门相对应的区域处使用弹性体材料。可以提前制备含有弹性体补片的膜并且使其无菌以组装芯片。这将促进器官芯片板的组装和操作,其中弹性体仅在板的非常局部区域中偏转以产生泵送动作。广泛范围的弹性体类型和厚度可能适用。c.用于控制润湿性的表面处理可以使用表面图案化控制器官芯片板中的敞开式流体通路的润湿性。机械加工图案包含斑纹(线性)表面、鲨鱼(shark)表面、同心表面和光滑表面。使用不同的机械加工工艺和微纹理化可能大大影响器官芯片的培养板的润湿性。表面抛光可以明显地将聚砜润湿性修改为高达与水或细胞培养基的接触角的40°变化。孵育器条件也可以在约2°到3°差异的轻微程度上增加润湿性。可能优选的是,介观流体装置具有增加的润湿性以提高性能。通常,深色聚砜比浅色聚砜更具疏水性。选择不同级别的聚砜提供了改变板的润湿性的另一手段。d.灭菌可以对细胞培养流体板、致动膜以及任选地气动板执行一个或多个灭菌程序。灭菌技术包含气体处理(例如,环氧己烷)、电离辐射、声处理、表面处理(例如,表面活性剂)以及高压灭菌。通常,在使用之前,对顶板(例如,聚砜顶板)进行清洁和灭菌。首先,可以将板浸没在约10%漂白剂中持续约30分钟到60分钟,随后是在蒸馏水中进行短暂冲洗。然后,使用无残留表面活性剂、通过浸没在约10%溶液(例如,7x溶液,mp生物医疗(mpbiomedicals)#mp0976680hp)中持续约15分钟进行声处理而移除任何剩余污染物。随后在新鲜的去离子水中进行两个随后的15分钟声处理循环以在最终去离子水冲洗之前移除所有表面活性剂。然后,可以使板风干,将其密封在灭菌袋中,并且对其进行高压灭菌。通常,气动板不需要正式灭菌,但是在组装之前,可以使用喷有70%乙醇的kimwipe对其进行彻底擦拭以将任何灰尘或颗粒从与膜接触的密封区域移除。可以在约10%7x溶液中并且使用过量去离子水对气动致动器膜进行冲洗。通常,在膜风干之后是环氧己烷气体灭菌步骤,并且允许膜在24小时内在化学通风橱中除气。e.细胞可以将如干细胞和潘氏细胞等分化细胞类型和特化细胞类型以及如肠mps等一些实施例的微生物组添加到平台。所述平台支持的微生理系统(mps)可以包括原代细胞、细胞系、多能干细胞、祖细胞、类器官、或哺乳动物或非哺乳动物细胞的任何组合。例如,在转移井插入物上由与ht29细胞混合的caco2细胞或caco-2细胞形成的上皮单层是肠的一个模型,其中基底隔室(转移井下方)中的循环用于改进混合并且因此将药物和其它药剂从上皮层的顶面输送到上皮层的基面;所述混合促进在基底配药物和其它化合物并且因此改进平台上的不同mps单元之间的整体混合。可以通过将免疫细胞源(例如,来自人外周血单核细胞或其它源的树突细胞或巨噬细胞)添加到膜的基面使此模型更复杂。可以通过在包封在细胞外基质胶中的基质顶部培养上皮使其更加复杂;类似布置可以与原代肠细胞一起使用。通过liverchip类型布置例示了第二类型的流动模块,其中流量被泵送通过含有3d组织的支架,所述3d组织在被设计成通过组织分配流量的支架上包括多种细胞类型。在另一配置中,在中心胶体区的任一侧上具有流动通道的封闭微流体装置可以支持如3d岛或淋巴结等组织,其中使用岛或淋巴结接种到凝胶中的内皮细胞形成允许通过所述通道对岛或淋巴结进行灌注的3d容器。还可以将岛或淋巴结保持在转移井插入物的胶体中,并且可以使转移井插入物的基面覆盖有内皮细胞。最后,可以将细胞添加到中心循环单元或单独的mps循环单元中的任何循环单元中以允许细胞转运。例如,添加到肠的基底隔室的pbmc可以通过炎症性信号流通到膜上的基质。在所公开设备中对肝中的三阴性乳腺癌(tnbc)(即,缺少雌激素、黄体酮和her2受体的表达)微转移进行建模的一些实施例中,可以培养mda-mb-231细胞连同原代人肝细胞和非实质细胞。在肠和肝mps被建模成评估对休眠微转移的炎症相关刺激的一些实施例中,可以在顶表面上、通常以20:1与5:1之间、更优选地13:1与7:1之间或约9:1的数量比接种吸收性肠细胞(例如,cc2bb/e1系)和粘蛋白分泌杯状细胞(例如,ht29-mtx系);而可以在设备的一个井中的的膜的下侧接种从人pbmc衍生的单核细胞的体外分化获得的树突细胞。可以根据用途添加其它细胞系或细胞类型。iv.应用体外到体内转移(ivivt)是将mps结果与临床相关结果进行比较并且对mps结果进行验证的解释性步骤。在预测药代动力学和药效学(pk和pd)时,可以将所公开的设备与ivivt方法一起应用以评估额外的因素,如内源生长因子、炎症信号和激素信号。相比于预测pk时的体内到体外相关(ivivc)和体内到体外外推(ivive)方法,ivivt进一步包含对这些额外因素的分析以及因此另外使用来自mps技术的信息预测pd、临床毒理学、生物标记以及患者分层。组合用于ivivt的基于生理学的pk(pbpk)模型,所公开设备提供关于人对测试剂的反应的改进定量预测,将缺失器官、器官与培养基大小失配以及药物暴露考虑在内。在一些实施例中,系统还可以用于例示疾病或障碍。例如,所述设备可以用于在相对较大量的肝细胞(≥1百万个细胞)的背景下建立微转移并且然后用于使用可以常规地在患者体内(在循环培养基上)进行的测量以及还无法在患者身体上进行的测量——肿瘤细胞生长和死亡的动力学——两者分析复杂的细胞-细胞通信网络特征。a.临床前药物发现mps支持一个或多个器官芯片在延长的时间段、如两周、三周、四周、五周、两个月、三个月或更长时间内的存活和功能培养。对于研究低清除率药物的药理学、支持重复药物暴露、分析药物-药物相互作用以及对慢性病进行建模来说,长期多器官培养特别有利。所述平台还可以用于靶识别和验证、靶相关筛选、表型筛选以及其它生物技术应用。细胞和培养基体积提供足够的信号以便进行如elisa等商业测定以及高含量多重测定。跨细胞中的信息流的尺度进行的多组学测量,从dna到rna到蛋白质、蛋白质活性状态和代谢物以及对患者衍生的免疫细胞功能的类似类型的分析。尽管标准培养系统对于大多数小分子药物pk测定来说是合理有效的,但是缺少充分治疗的大量疾病具有炎症含义并且未在标准培养系统中予以良好表示或建模。所述设备可以特别适合于通常涉及免疫系统的药物开发后期。已经显示,所述设备概括了抗il6受体抗体、托珠单抗与辛伐他汀的代谢之间的复杂的基于免疫学的药物-药物相互作用,这是在标准培养中无法复制的现象(longt等人,《药物代谢与处置(drugmetabdispos)》,第44卷,第1940到1948页(2016年))。可以针对药用应用、化妆品应用或科学应用而在所公开设备中对广泛范围的药剂(小分子、肽、蛋白质、核酸等)进行测试。通常,对混合井进行添加模拟静脉内给药,并且对肠井进行添加模拟口服给药。药剂是基于待治疗或预防的疾病或障碍来选择的。b.待建模的疾病或障碍多器官设备是用于疾病建模和药物开发的有用工具,特别是在识别和定义用于表示疾病状态的相互作用器官系统的适当“最小集”时有用的工具。可以利用所公开的多器官芯片设备、通过培养用于全身性研究的相关组织或细胞类型改进针对各种疾病和/或障碍的药物开发。优选地,捕获疾病的局部特征、特别是炎症的复杂的单独器官芯片适用于对全身性疾病或与多个器官相关联或者涉及多种类型的细胞的疾病进行建模。可以在所公开的生物反应器中建模的疾病和/或障碍包含但不限于癌症/肿瘤(例如,乳房、骨、肝、肺和脑中的肿瘤)、慢性炎症性疾病(例如,糖尿病、关节炎、子宫内膜异位症以及阿尔兹海默病)、子宫内膜非恶性生长在子宫外部(子宫内膜异位症)或移位到子宫肌中(子宫腺肌症)、肝功能异常,如非酒精脂肪肝病引起的肝功能异常。所述系统提供用于使细胞暴露于药剂以确定药剂对细胞的作用的装置,按不同剂量、按不同给药方案、或者和组合一种或多种其它药剂施用所述药剂,并且确定其对细胞的作用,以及其中药剂被施用给不同细胞类型或与一种或多种疾病或障碍相关联的细胞类型。这允许人们施用人细胞、在模拟人的条件下、至少部分地在受控条件下在体外对药剂进行测试,从而寻找对其它细胞类型的作用以及人们想要监测以发现作用的细胞的作用。这是更快速的、更受控的并且又不受限于单一类别的细胞或组织。实例将参考以下非限制性实例进一步理解本发明。实例1.所灌注单器官微生理系统(mps)芯片(1)肝:对所灌注的肝细胞-库普弗细胞共培养物进行培养持续三周。材料和方法对具有代谢和免疫能力的3d冷冻保存的人肝细胞和库普弗细胞进行共培养。已经在mps中对多个肝细胞和库普弗细胞供体进行定性。共培养物对下至0.01μg/ml的脂多糖(lps)刺激物有反应。结果表1示出了在所灌注mps平台中在7天内对仅肝细胞与比例为10:1的肝细胞和库普弗细胞的共培养物的比较。表1肝细胞对具有免疫能力的肝mps的生物功能。第7天的功能(n=3)仅肝细胞肝细胞+库普弗细胞(10:1)白蛋白(μg/天/mg)35±1153±32尿素(μg/天/mg)175±75184±25cyp3a(pmol/分钟/mg)2.9±0.52.0±0.7测量了共培养的肝mps的白介素6(il-6)和肿瘤坏死因子α(tnfα)的分泌物。确定了il-6对lps刺激物的反应的再现性。在共同培养基中使用了生理相关(相对较低)水平的皮质醇。氢化可的松(皮质醇)增强了分化功能,但是抑制了炎症反应。灌注后mps平台上的冷冻保存的人肝细胞和库普弗细胞共培养物的持续时间延长到了3周。对84个肝基因的表达在第7天与第21天之间保持稳定。库普弗细胞保持灭活21天,直到使用lps进行刺激。细胞死亡标记ldh在接种之后衰落并且保持处于很低的恒定水平。肝表型活性、包含白蛋白和cyp450保持可测量并且优于2d培养物持续21天。cyp450活性对培养物中的氢化可的松水平敏感。(2)肺:原代人气管支气管上皮分化。材料和方法在mps平台上在transwell中开发了气管支气管模块。原代基底上皮细胞(所有ck5+)在气体-液体界面处分化成上皮细胞类型的完整子集。对不同的度量进行了评估,包含跨上皮电阻(teer)、粘液产生、分化后细胞群以及基底il-8产生。结果取得了荧光显微图像并且荧光显微图像确认了对分化和功能标记的表达:微管蛋白(有纤毛的)、ck5(基底)、muc5ac(杯状)以及鬼笔环肽(肌动蛋白)。表2对肺mps平台上的分化后细胞的估计和测量百分比的比较表2确认肺mps模型支持细胞分化,到达与生理估计良好一致的程度,这指示原代人气管支气管上皮模型的功能。(3)子宫内膜:子宫内膜-基质的半原代共培养物稳定且在功能上分泌糖蛋白。材料和方法在月经周期内,人子宫内膜经历由雌激素水平升高标记的增生期和由黄体酮水平升高标记的分泌期。在分泌期内,子宫内膜分泌特有蛋白,如免疫抑制性糖蛋白、催乳素、胰岛素样生长因子结合蛋白1(igf-bp1)。mps平台上的中的细胞培养系统的示例性子宫内膜模型包含包封基质细胞的水凝胶,并且在的顶面对水凝胶的顶面上的上皮细胞进行了培养。上皮细胞源是原代人子宫内膜上皮细胞,所述细胞容易从子宫内膜活检中获得,在培养时具有受限的扩增和寿命,在增生期和分泌期内的产量之间展现出功能差异,并且支持鲁棒的免疫抑制性糖蛋白(分泌期细胞)。所使用的细胞系是石川人阶段1adenocarincoma细胞系,所述细胞系是雌激素和黄体酮受体阳性的,在基质胶中(chitcholtan等人,《实验细胞研究(expcellresearch)》(2013年))或功能化peg凝胶中偏振,并且具有低/可变的免疫抑制性糖蛋白分泌。基质细胞源是原代人子宫内膜基质细胞,所述细胞容易从人活检中获得,具有公认的体外扩增方案,并且在增生期与分泌期中的产量之间显示出功能差异。所使用的细胞系是人第三永生化细胞系(thesc),所述细胞系可高度增生且高度稳定,具有低/可变催乳素或igf-bp1分泌而不具有强烈的蜕膜化线索,并且将在peg凝胶中静默。结果在石川上皮细胞的情况下,顶端培养基含有雌二醇和黄体酮。石川免疫抑制性糖蛋白分泌低于检测限制。对thesc的平台外培养已经由原代细胞产生了刚好处于检测限制的igf-bp1以及边界可检测量的催乳素(prl)(由于稀释效应而可能)。这些“半原代”细胞系的平台上共培养物稳定且具有可根据顶端采样检测的功能。(4)肠/免疫:两周共培养形成完好屏障,并且药物诱导的泄露触发免疫反应。材料和方法生理肠系统以以下各项为特征:吸收和代谢、肠免疫系统、微生物组之间的相互作用和粘膜相互作用、细胞与微生物组之间的免疫相互作用以及药物免疫相互作用。具有mps平台上的中的细胞培养物的示例性的具有免疫能力的肠模型包含肠细胞,并且在transwell的顶面上对杯状细胞进行了共培养并且在transwell膜的基面对免疫细胞进行了共培养。所使用的细胞系为caco2(肠细胞)、ht29-mtx(杯状细胞)以及树突细胞(免疫细胞),其中在平台外以9:1的比例对肠细胞:杯状细胞进行了培养(模拟小肠)持续2周直到成熟并且将其被转移到transwell膜的基面上具有所添加免疫细胞的灌注平台。结果相比在具有基底外侧流量的情况下在平台上培养的情况,当在平台外培养(静态培养基)时,免疫细胞在第14天具有低得多的存活率,如通过免疫荧光显微术确认的。平台上培养物在第14天支持肠屏障细胞的功能及其分化。实例2。在单独或双向mps芯片中评估药物毒性。(1)肝/免疫:双氯芬酸和托卡朋的毒性。制备且研究了具有免疫能力的肝mps模型。双氯芬酸损伤肝功能,而细胞死亡最低。托卡朋降低线粒体活性并且引起细胞死亡。(2)肠/免疫:双氯芬酸和托卡朋的毒性。制备且研究了具有免疫能力的肠mps模型。双氯芬酸降低上皮屏障完整性,从而引起具有最低细胞死亡的漏肠。托卡朋引起炎症的细胞死亡,因此完全失去上皮功能。(3)子宫内膜mps:双氯芬酸和托卡朋的毒性。制备且研究了子宫内膜mps模型。双氯芬酸诱导与细胞死亡相关的功能丧失。托卡朋诱导与细胞死亡相关的功能丧失。(4)肠-肝双向:对肠施用(“经口”)托卡朋导致肠特异性毒性。材料和方法制备(同样在实例3中示出了细节)并研究了具有免疫能力的肠-肝相互作用的mps模型。托卡朋被添加到肠mps中以模拟“经口”施用。度量为来自3个培养基隔室的体积加权平均值:信号全身=信号顶端,肠*v顶端,肠+信号基底,肠*v基底,肠+信号肝*v肝。结果在存在托卡朋的情况下,肠和肝遭受mps特异性功能损失,这指示托卡朋的mps特异性毒性,甚至是在托卡朋“经口”递送的情况下。肠和肝还遭受mps特异性细胞死亡标记,而通用标记对托卡朋不敏感,所述标记指示托卡朋的毒性位点。小肠型脂肪酸结合蛋白(i-fabp)被用作各种疾病状态的肠上皮细胞损害的临床生物标记。实例3。肠-肝双向mps中的炎症细胞因子/趋化因子串扰。在此体外平台中研究了具有免疫能力的人肝(干细胞和库普弗细胞)组合肠(肠上皮细胞、杯状细胞、和树突细胞)微生理系统,以在检查正常和炎症背景下的肠-肝相互作用。肝位于肠的下游;如此,肝始终暴露于肠衍生因子,包含代谢物、微生物抗原以及炎症介体。然而,对这些多细胞组织如何通信并且贡献于整体(病理)生理的定量理解有限。背景肠-肝串扰是正常生理的整体部分,并且其调节异常是许多疾病状况中的共同特征(marshalljc,《创伤、休克和败血症中的宿主防御功能障碍:机制和治疗方式(hostdefensedysfunctionintrauma,shockandsepsis:mechanismsandtherapeuticapproaches)》,编者faiste、meakinsjl和schildbergfw(海德堡柏林的施普林格出版社(springerberlinheidelberg),第243到252页(1993年))。此外,肠和肝是涉及药物吸收和代谢的主要器官;其功能相互作用的变化可能影响其对治疗介入的反应(morganet,《药物代谢与处置(drugmetabdispos)》,第29卷,第3期,第207到212页(2001年);dengx等人,《药理学综述(pharmacologicalreviews)》,第61卷,第3期,第262到282页(2009年);longtj等人,《药物代谢与处置(drugmetabolismanddisposition)》,第44卷,第12期,第1940到1948页(2016年))。肠和肝功能由于其在解剖学上的相近关系而密切相关。肝通过端口循环从肠接收其70%的血液供应。如此,肝始终暴露于肠衍生因子,包含代谢物、微生物抗原以及炎症介体。肠-肝轴线显著贡献于身体的整体免疫状态,其中肠是最大的免疫器官,并且肝含有身体中总巨噬细胞群体的超过70%。种间差异常常阻碍在动物模型中准确预测人反应;在涉及免疫系统的过程中,差异特别明显(mestasj等人,《免疫学杂志(thejournalofimmunology)》,第172卷,第5期,第2731到2738页(2004年);giesec等人,《先进药物输送评论(advdrugdeliverrev),第69卷,第103到122页(2014年))。例如,针对败血症治疗的少量临床试验已经引起药物批准(seokj等人,《美国国家科学院院刊(procnatlacadsciusa)》,第110卷,第9期,第3507到3512页(2013年));finkmp,《毒力(virulence)》,第5卷,第1期,第143到153页(2014年))。在败血症中,胃肠损伤和肝损伤已经与疾病严重性增加相关联(rowlandsbj等人,《英国医学公报(britishmedicalbulletin)》,第55卷,第1期,第196到211页(1999年);nesselern等人,《重症护理(critcare)》,第16卷,第5期,第235页(2012年))。在败血症开始后的前72小时内的急性肝衰竭与败血症患者体内的不良预后高度相关。然而,缺少特异性生物标记和预测性生物标记妨碍用于进行有效介入的早期诊断和患者分层(pierrakosc等人,《重症护理(critcare)》,第14卷,第1期,第r15页(2010年))。尽管已经在败血症反应升级中暗示了肠-肝轴线,但是所涉及的机制和分子参与者定义不清。因此,对肠-肝串扰的基本理解不仅对于预测药物处置、疗效和毒性来说,而且对于说明(病理)生理机制来说是关键的。材料和方法在体内,肝从肝动脉和门静脉接收双血液供应(liaskoue等人,《炎症介体(mediatorsinflamm)》,第2012卷,第949157页(2012年);brownrp等人,《毒理学和工业卫生(toxicolindhealth)》,第13卷,第4期,第407到484页(1997年))。相应地,来自混合器井的流量被分割成肠隔室和肝隔室以便分别按75%和25%缩放成与生理心输出量和肝血流量成比例,如下表3和表4所示。来自肠模块的输出给送到肝中,从而表示门脉循环。使用了15ml/天的全身再循环流速以确保外源因子和内源因子的高效分配。表3肠-肝mps中的示例性受控流速表4肠-肝mps中的示例性受控体积此研究中的肝和肠组织构建体是多细胞的且(先天)具有免疫性的,被设计成涵盖多种关键功能,包含代谢、屏障和免疫功能。肝微组织包括生理比例为10:1的人原代冷冻保存肝细胞和库普弗细胞的共培养物,保持在允许保留炎症反应的培养基中,如之前所描述的(longtj等人,《药物代谢与处置(drugmetabolismanddisposition)》,第44卷,第12期,第1940到1948页(2016年);sarkaru等人,《药物代谢与处置(drugmetabolismanddisposition)》,第43卷,第7期,第1091到1099页(2015年))。肠组织被工程化为模拟小肠,其中上皮单层源自于吸收性肠细胞(caco2-bbe)和产生粘液的杯状细胞(ht29-mtx)的9:1比例,并且免疫隔室含有原代单核细胞衍生的树突细胞。从美国生命技术公司(lifetechnologies)(hmcpms、hukccs)购买了人原代肝细胞和库普夫细胞。在70%etoh中对支架进行洗涤15分钟,在psb中对其冲洗两次,在pbs中在30ng/ml鼠尾胶原蛋白中在室温下对其进行孵育1小时,使其在室温下过夜干燥,并且将其推进平台中(保持环下面的支架下面的过滤器)。在实验开始前第3天,将10:1比例的肝细胞和库普弗细胞解冻到温热的冷冻保存肝细胞恢复培养基(cryopreservedhepatocyterecoverymedium,chrm,英杰公司(invitrogen))中,使其在100g下旋转8分钟,并且分别以6*105和6*104细胞/井将其接种在冷的肝细胞接种培养基(250ml高级dmem+9mlgibco混合物a+12.5mlfbs)。一天之后,培养基被改变为倒数第2天培养基(250ml高级dmem+10ml混合物b)。再过两天之后,培养基被改变为共同培养基,持续相互作用实验持续时间。本研究中使用的共同培养基由500mlwilliamse培养基+20mlgibco混合物b+100nm氢化可的松+1%盘尼西林-链霉素(p/s)组成。caco2(克隆:c2bbe1,atcc,48到58代)和ht29-mtx(sigma,20到30代)细胞系用于肠上皮培养。两个细胞系在解冻后在用于transwell接种之前传代两次。细胞系保持在补充有10%胎牛血清(atlantabiologicals,s11150,在57℃下热灭活(hi)30分钟)、1xglutamax(gibcotm35050-061)、1x非必需氨基酸(gibcotm15140-148)以及1%盘尼西林-链霉素(gibcotm15140-148)的dmem(gibcotm11965-092)中。使用0.25%的胰蛋白酶-edta(gibcotm25200-056)收获了~70%到80%汇合率的caco2以及~80%到90%汇合率的ht29-mtx,并且将其机械分裂成单一细胞以供transwell接种。在将细胞接种到中时,在4℃下过夜使用50μg/ml的i型胶原蛋白(康宁公司(corning),354236)来涂覆transwell膜的顶面和基面。使用pbs-/-来洗涤插入物以移除非结合蛋白质。以105细胞/cm2的密度将9:1比例的c2bbe1和ht29-mtx接种到孔径为0.4μm的12井插入物(costar3460)中。接种培养基含有10%热灭活的fbs、1xglutamax、高级dmem(gibcotm12491-015)中的1%p/s。顶面培养基在接种后第1天被替换以移除任何未附接细胞。transwell板的顶部隔室和底部隔室每2天到3天给送有0.5ml和1.5ml的接种培养基。7天之后,通过使用胰岛素(5μg/ml)-转铁蛋白(5μg/ml)-亚硒酸钠(5ng/ml)(roche11074547001)来代替fbs而将培养基切换为无菌肠培养基。为了在肠-肝相互作用中评估长期功能生存力,使用完全相同的培养基体积、流速和流量分割对平台上的相应单一组织对照物进行了测定。在支持保持肠功能和肝功能的限定的无菌共同培养基中对所有状况进行了测试。肝细胞(10:1肝细胞:库普弗细胞)在相互作用实验开始前3天被接种在平台上以允许组织形成和从接种相关应激反应中恢复。肠mps在相互作用实验开始前在平台外分化3周。在长期操作期间,系统中的共同培养基每3天替换。为了评估肝的健康,在每次培养基改变(每72小时)时取得来自所有隔室的样本并且通过elisa对白蛋白进行测定(bethyllaboratories,e80-129)。使用发达的cyp混合物测定测量了各种细胞色素p450(cyp)酶活性(pillaivc等人,《药物与生物医学分析杂志(jpharmbiomedanal)》,第74卷,第126到132页(2013年))。简言之,cyp基底的混合物被添加到肝隔室中进行一小时孵育,并且收集了上清液以供下游处理。使用质谱法分析了基底特异性代谢物产量。单核细胞衍生的树突细胞被用作肠mps的免疫组分。简言之,对leukopak(stemcelltechnologies,70500)进行处理而得到外周血单个核细胞(pbmc),并且将其储存在液氮中。对于每项实验来说,解冻pbmc,并且使用easysep人单核细胞富集试剂盒(stemcelltechnologies,19058)分离单核细胞。单核细胞在补充有1xglutamax、1%p/s、50ng/mlgm-csf(biolegend572903)、35ng/mlil4(biolegend574004)以及10nm视黄酸(sigmar2625)的高级rpmi培养基(gibcotm12633-012)中分化成树突细胞。在分化7天后(在肠上皮细胞成熟的第19到20天),使用accutase(gibcotma11105-01)收获了免疫树突细胞并且将其播种到倒置肠的基面上持续2小时。2小时后,将细胞返回到培养板并且给送以肠培养基。在树突细胞接种后一天,对肠屏障功能进行了评估。具有至少250ohm*cm2的跨上皮电阻值的肠mps被视为可接受用于实验。对于平台上的所有相互作用实验来说,肠mps保持在共同培养基中。使用endohm-12室、利用evom2仪(世界精密仪器(worldprecisioninstruments))执行了teer测量。样本和endohm室在热板上在~37℃下保温。在teer测量期间严格保持温度以使变化性最小化。在顶端肠隔室中、使用阿尔新蓝(alcianblue)测定对所分泌的粘液进行了测量。从(5)起改变粘液定量方案。简言之,来自顶端的培养基被收集在低结合管中,并且在10,000g下向下旋转5分钟,并且上清液被收集且在-80℃下储存以供随后分析。针对溶入培养基中的粘液标准品(sigma-aldrichm3895)对粘液分泌物进行了定量。在96井板中、在样本和阿尔新蓝溶液(richardallenscientific)的3:1混合物中对样本和标准品进行孵育两小时。在孵育之后,在室温下、在1640g下对板进行离心30分钟。通过将板倒置移除上清液。使用洗涤缓冲液(40%(v/v)的乙醇和含有ph为5.8的25mmmgcl2的60%(v/v)的0.1m乙酸钠缓冲液)将样本冲洗两次,其中在每次冲洗之后进行10分钟离心步骤。在第二次旋转之后,将上清液移除,并且将赝本与10%sds一起溶于蒸馏水中。板通常需要摇动或移液以使样本完全重新悬浮。如果在重新悬浮期间形成气泡,则在620nm的spectramaxm3/m2e上进行吸收率测量之前再次对板进行离心约5分钟。使用多重细胞因子测定、37丛人炎症面板和40从面板趋化因子面板(美国加尼福尼亚州赫拉克勒斯bio-radlaboratories,inc.)测量细胞因子水平。简言之,培养基样本被收集在低结合管中,在10,000g下向下旋转5分钟以移除细胞碎片,并且在-80℃下储存上清液。以多个稀释度对样本进行测量以确保测量结果处于测定的线性动态范围内。为了使与珠的非特异性结合最小化,添加了牛血清蛋白(bsa)以实现在所有样本中实现最终5mg/ml的浓度。在同一培养基中重构蛋白标准品,并且连续稀释蛋白储备液以产生8点标准曲线。在bio-plex3d悬浮阵列系统(bio-radlaboratories,inc.)上运行了测定。使用flexmap3d软件第4.2版的xponent(美国德克萨斯州奥斯汀路明克斯公司(luminexcorporation))收集了数据。根据标准曲线确定每种分析物的浓度,所述标准曲线是通过将平均荧光强度的5参数逻辑回归拟合到每种分析物的已知浓度而产生的(bio-plexmanager软件)。为了获得每平台的总产量,通过隔室体积使浓度归一化并且跨每个平台的所有隔室(混合器、肠、肝)相加。对于基线状况和发炎状况(在第3天,n=4)两者来说,从平台中取出肠组织和肝组织,并且使用purelinkrna迷你试剂盒(thermofisher,12183018a)提取mrna。使用片段分析仪(advancedanalytical)对总rna进行分析和定量,并且使用smart-seqv3试剂盒(clontech)产生cdna。在cdna片段化(covariss2)之后,使用spri工厂片段库系统i(beckmancoultergenomics)对cdna进行末端修复和衔接子连接。然后在pcr扩增期间对衔接子连接的cdna进行索引,并且在illuminahiseq2000上测序之前使用片段分析仪和qpcr对所产生的库进行定量。分别针对基线状况和发炎状况而对具有每样本1500到2000万或500万读数的平均深度的40-50nt单端读数进行测序。根据测序运行产生fastq文件。使用tophat(版本2.0.12)将所产生的读数与人参考基因组(grch37/hg19)对齐(kimd等人,《基因组生物学(genomebiol)》,第14卷,第4期,第r36页(2013年))以识别映射到已知转录本的读数,从而解释剪接点。使用htseq来确定与已知基因组特征唯一重叠的读数计数的数量(anderss等人,《生物信息学(bioinformatics)》,第31卷,第2期,第166到169页(2015年))。为了识别隔离状况和相互作用状况下的显著变化基因,在r中使用deseq2(版本1.12.3)执行对计数数据的差异基因分析(lovemi等人,《基因组生物学(genomebiol),第15卷,第12期,第550页(2014年))。仅在至少4个重复中具有大于1cpm(每百万计数)的基因包含在分析中。使用benjamini和hochberg过程执行多测试校正。具有低于fdr截止值0.05的调整后p值的基因被视为是显著的。使用goseqr包(youngmd等人,《基因组生物学(genomebiol)》,第11卷,第2期,第r14页(2010年))来确定差异表达的基因(fdr调整后p值<0.05)的过表达生物制剂。执行gsea(版本2.2.3)以识别隔离和相互作用的差异调控的基因集合,如(subramaniana等人(2005年),《美国科学院院报(proceedingsofthenationalacademyofsciences)》,第102卷,第43期,第15545到15550页)中所述。为了稳定方差,在deseq2中使用正则化对数变换对归一化计数数据进行处理。使用信噪比度量来产生基因排序表。使用了来自分子特征数据库(c2.cp.v5.2)的经典途径基因集合,所述分子数据特征数据库是来自多个数据库(例如,reactome、kegg、biocarta、pid)的一批精选基因集合。相对于富集分数的零分布计算了每个富集分数的经验p值,所述零分布是通过1000个基因集合排列来计算的。具有标称p值<0.01和q值<0.05的基因集合被视为是显著的。使用富集图(11)、即cytoscape插件来使显著基因集合之间的重叠可视化并且促进对不同生物过程支架的相关性的系统解释。结果1.在肠-肝相互作用组中,基线肝特异性功能和肠特异性功能保持相对较长时间(>2周)。在两周培养内评估的肝功能和肠功能对于保持通信或隔离的mps来说是相当的,如通过测量白蛋白产量、肠屏障完整性以及肠粘液产量而评估的。为了在2周实验结束时评估肝代谢功能,对来自隔离状况和相互作用状况(不存在肠)的肝组织给予靶向特异性cyp450酶的药物底物混合物。使用质谱法对培养基中的药物特异性代谢物产量进行了测量以确定不同亚型的细胞色素p450活性。总体来说,肝代谢功能在很大程度上得以保持,其中在肠-肝相互作用中观察到对所选细胞色素p450的调节。具体地说,cyp2c9活性得到显著增强,而cyp3a4/5活性得到抑制。肠特异性功能、包含屏障完整性和粘液产量在相互作用对照物与隔离对照物之间未被显著改变。在2周相互作用之后观察到对细胞素色p450活性(例如,cyp3a4和cyp2c9)的细微但显著调节。2.胆汁酸合成途径在双向肠-肝串扰中得到调节。在与相应的隔离对照物(即,仅肠和仅肝)的3天相互作用之后执行了rna测序以概括肠组织和肝组织的总体转录组变化。在相对于隔离对照物的相互作用期间,肝中有105个基因显著(fdr调整后的p<0.05)变化,其中70个基因上调并且35个下调。对于肠来说,6个基因显著差异表达,其中2个上调并且4个下调。为了理解这些分子变化的功能含义,执行了基因本体(go)分析以识别在相互作用下改变的过表达生物过程。仅将显著变化的基因(fdr调整后的p<0.05)用于go分析。肝中的上调生物过程主要涉及细胞分裂相关过程(表5)。表5肠-肝相互作用下肝中上调的生物过程在肝中诱导细胞周期细胞可以表明对肠衍生信号的适应性反应,但是所涉及的可溶性因子是未知的。另一方面,肝中的下调生物过程主要是代谢过程,包含胆汁酸生物合成、脂质代谢以及异生物质代谢(表6)。表6基线肠-肝相互作用下肝中下调的生物过程具体地,胆汁酸代谢的介体cyp7a1下调,这指示肠-肝功能、例如视黄酸介导的肠肝串扰的生理偶联。cyp7a1是对于胆汁酸合成来说具有中心作用的酶;并且其通过fgf19肠肝通信进行的反馈抑制是公认的(dingl等人,《药学学报b(actapharmsinb)》,第5卷,第2期,第135到144页(2015年))。cyp7a1上的结果与之前的发现一致,所述之前的发现即将精密切割小鼠肠组织和肝组织灌注在微流体装置中持续7小时导致胆汁酸介导的cyp7a1抑制(vanmidwoudpm等人,《芯片实验室(labchip)》,第10卷,第20期,第2778到2786页(2010年))。尽管肠样本中的显著基因的数量对于go分析来说是不充分的,但是发现差异表达的基因之一pcsk9在胆固醇和脂质体内平衡中起到关键作用。事实上,已经显示,胆固醇和各种类型的胆汁酸在caco2肠培养物中抑制pcsk9mrna表达(leblondf等人,《美国生理学杂志:胃肠与肝生理学(amjphysiolgastrointestliverphysiol)》,第296卷,第4期,第g805到g815页(2009年))。研究表明,在胆固醇和胆汁酸代谢途径上汇聚指示由于mps间通信而产生的转录重新布线。3.在炎症性肠-肝串扰中观察到协调的转录组变化和组织特异性转录组变化。在体内平衡期间,大量免疫细胞驻留在肠和肝中,并且所述免疫细胞在疾病中活化可能有助于系统病理生理学。与特质药物不良反应相关联的肝功能异常已经与不适当的免疫活化关联(cosgrovebd等人,《毒理学及应用制药学(toxicologyandappliedpharmacology)》,第237卷,第3期,第317到330页(2009年))。此研究使用肠和肝中的免疫细胞来补充实质组织模型以便为疾病建模和药物测试提供更具生理相关性的培养平台。往复免疫-上皮细胞通信驱动全身性炎症。在模拟内毒素血症的炎症性环境中,从第0天(平台上的肠mps)到第3天(执行了rnaseq),在循环培养基中添加了2ng/ml脂多糖(lps),同时以与上述基线研究类似的方式对系统和隔离对照物的操作进行了测试。基于在患有炎症性疾病的患者中报告的血浆内毒素的临床相关范围(2ng/ml到10ng/ml)选择了lps浓度(guos等人,《美国病理学杂志(amjpathol)》,第182卷,第2期,第375到387页(2013年))。执行了rna测序以评估与炎症性肠-肝串扰相关联的总体分子变化。对于肝来说,2548个基因在相互作用中显著变化,其中1137个基因上调,并且1411个基因下调。对差异表达的基因的go分析显示了对细胞因子反应和抗原加工和呈递途径的上调以及对脂质和异生物质代谢途径的下调(表7)。表7炎症性肠-肝相互作用下肝中上调的生物过程表8炎症性肠-肝相互作用下肝中下调的生物过程对于肠来说,780个基因在相互作用期间显著变化,其中290个基因上调,并且490个基因下调。类似地,go分析揭示对防御反应、抗原加工和呈递途径以及蛋白质转译的上调;下调途径包含乙醇生物合成、类固醇和脂质代谢(表9)。表9炎症性肠-肝相互作用下肠中上调的生物过程表10炎症性肠-肝相互作用下肠中下调的生物过程除了仅集中于通过任意统计截止确定的显著变化基因的基于基因的go分析之外,还执行了基因集合富集分析(gsea)以发现功能上相关的基因群组的协调变化。gsea可以揭示可能已经被基于基因的方式中的严格截止掩蔽的更微妙的途径调控。总体上,gsea结果在很大程度上与go分析结果一致,但是具有更大的可解释性和普遍性。获得了来自不同数据库的基因集合的共识集群,所述共识集群含有定义主要生物过程的重叠但不同的基因群组。具体地,在相互作用下,在肠和肝两者中,以ifnα/β/γ信号传递为中心的炎症相关途径上调,而涉及胆固醇和脂质代谢的代谢过程下调(表11)。炎症性过程和脂质代谢的显著变化以败血症反应为特征。表11在肠-肝mps中的肠和肝两者中共同上调的基因集合。途径肝:q值肠:q值表12在肠-肝mps中的肠和肝两者中共同下调的基因集合。途径肝:q值肠:q值除了共同调节的途径之外,还识别到了组织特异性调控。缺氧和tgfβ/smad信号传递中涉及的途径在相互作用下的肝中唯一上调,这暗示促纤维化反应。尽管当前研究集中于急性炎症,但是慢性肝炎症已经与肝纤维化关联。在肠中,pi3k介导的erbb2和erbb4信号传递上调,这指示伤口愈合或抗凋亡反应,有可能充当保护机制。之前,已经在体外和体内显示了erbb2(yamaokat等人,《美国国家科学院院刊(procnatlacadsciusa)》,第105卷,第33期,第11772到11777页(2008年);zhangy等人,《实验室调查(labinvest)》,第92卷,第3期,第437到450页(2012年))和erbb4(freymr等人,《胃肠学(gastroenterology)》,第136卷,第1期,第217到226页(2009年))以防范肠上皮细胞中的tnf诱导的细胞凋亡并且在肠损伤之后提供促生存和促愈合效果。以下示出了组织特异性调节中涉及的完整基因集合列表:在炎症性肠-肝串扰期间在肝中唯一上调的基因集合包含:biocartatnfr2途径、st肿瘤坏死因子途径、pidtnf途径、reactome趋化因子受体结合趋化因子、kegg细胞因子细胞因子受体相互作用、keggrigi样受体信号传递途径、kegg胞质dna感测途径、reactomerigimda5信号传递的负调控物、naba分泌因子、pidcd40途径、pidhif1tf途径、pidhif2途径、pidil23途径、kegg原发性免疫缺陷、reactome根据干扰素刺激的基因的抗病毒机制、reactome粘蛋白的)o.o连接糖基化、reactome通过氧气对低氧诱导因子hif的调控、reactome对干扰素αβ途径的rigimda5介导的诱导、reactome通过tgfβ受体复合体进行的信号传递、reactomesmad2smad3smad4异三聚体调控转录、reactometraf6介导的irf7活化、reactomesmad2smad3smad4异三聚体的转录活性以及stfas信号传递途径。在炎症性肠-肝串扰期间在肠中唯一上调的基因集合包含:pidil12_2途径、keggabc转运体、reactome氨基酸合成和互换转氨作用、kegg氨酰trna生物合成、reactome胞质trna氨酰化、reactometrna氨酰化、kegg细胞粘附分子cam、kegg组氨酸代谢、reactome通过atf4进行的基因活化、reactomeperk调控的基因表达、reactomeerbb2信号传递中的pi3k事件以及reactomeerbb4信号传递中的pi3k事件。在炎症性肠-肝串扰期间在肝中唯一下调的基因集合包含:biocartaami途径、biocarta内源性途径、kegg丙氨酸天冬氨酸盐和谷氨酸盐代谢、kegg花生四烯酸代谢、kegg精氨酸和脯氨酸代谢、kegg丙胺酸代谢、kegg不饱和脂肪酸的生物合成、kegg丁酸甲酯代谢、kegg柠檬酸盐循环tca循环、kegg补体系统、kegg药物代谢细胞色素p450、kegg药物代谢其它酶、kegg脂肪酸代谢、kegg甘氨酸丝氨酸和苏氨酸代谢、kegg糖酵解糖异生、kegg丙酸代谢、kegg乙醛酸盐和二甲酸酯代谢、kegg组氨酸代谢、kegg亚油酸代谢、kegg赖氨酸降解、kegg氧化磷酸化、kegg帕金森氏病、kegg过氧物酶体、kegg近端小管碳酸氢盐回收、kegg丙酮酸盐代谢、kegg视黄醇代谢、kegg色氨酸代谢、kegg酪氨酸代谢、kegg缬氨酸亮氨酸和异亮氨酸降解、pidhnf3b途径、reactome生物氧化、reactome支链氨基酸分解代谢、reactome三羧酸循环tca循环、reactome脂肪酸三酰甘油和酮体代谢、reactome形成纤维蛋白凝块凝血瀑布、reactome氨基酸和衍生物代谢、reactome过氧化物酶体类脂代谢、reactome阶段ii缀合、reactome丙酮酸酯代谢和柠檬酸tca循环、reactome呼吸电子转运、通过化学渗透耦联的atp合成以及通过解偶联蛋白质的热产生、reactome胆汁酸和胆汁盐合成、reactome通过7α羟基胆固醇的胆汁酸和胆汁盐合成以及reactometca循环和呼吸电子转运。在炎症性肠-肝串扰期间在肠中唯一下调的基因集合包含:biocartatnfr2途径、keggdna复制、kegg泛酸盐和辅酶a生物合成、kegg戊糖和葡萄糖醛酸酯互换、kegg类固醇生物合成、kegg萜类主链生物合成、pid极光b途径、pidhif1tf途径、reactome响应于复制应力而活化、reactome对前复制复合体的活化、reactome胆固醇生物合成、reactome在着丝粒处含有核小体的新cenpa的沉积、reactomedna链伸长、reactome对dna复制的e2f介导的调控、reactome脂酰辅酶a生物合成、reactome通过ccttric形成微管蛋白折叠中间体、reactomeg1s特异性转录、reactomeg2m检查点、reactome对维生素核苷酸和相关分子的转运以及reactome甘油三脂生物合成。4.全身性炎症抑制肝解毒功能。内源化合物和异生物质化合物的肝清除由两种机制介导,即代谢和胆汁消除。结果解释炎症性串扰负面影响这些途径两者并且可能导致有毒副产物积累。cyp1a2、cyp2c9、cyp2c19、cyp2d6、、cyp3a4以及cyp3a5共同负责超过90%的已知药物的代谢(jacoba等人,《国际临床和实验医学杂志(intjclinexpmed)》,第2卷,第3期,第203到211页(2009年);ebrahimkhanimr等人,《先进药物输送评论(advdrugdelivrev)》,第69和70卷,第132到157页(2014年))。所有这些在整合系统中的肝中可能由于如il6、tnfα和/或i型干扰素等炎症性介体的积累而受到抑制(longtj等人,《药物代谢与处置(drugmetabolismanddisposition)》,第44卷,第12期,第1940到1948页(2016年);huangsm等人,《临床药理与治疗(clinpharmacolther)》,第87卷,第4期,第497到503页(2010年))。简言之,脂质代谢和验证是在肠-肝相互作用期间改变的显性途径。脂蛋白与lps结合可以将从库普弗细胞中摄取的lps重新引导到肝细胞,由此减弱免疫活化并促进对lps的胆汁清除(khovidhunkitw等人,《脂质研究杂志(jlipidres)》,第45卷,第7期,第1169到1196页(2004年))。已经显示,脂质代谢的主要调控因子过氧化物酶体增殖物激活受体(ppar)发挥抗炎症作用(vargat等人,《生物化学生物物理学学报(biochimbiophysacta)》,第1812卷,第8期,第1007到1022页(2011年))。合在一起,在炎症性肠-肝串扰期间观察到的对载脂蛋白合成和ppar信号传递的抑制指示潜在的保护机制缺失,由此加剧免疫细胞和上皮细胞中的炎症。对扰动的全身性反应的复杂性促生了对多细胞和多器官实验模型的需要。败血症患者易患由于对肝代谢功能、特别是细胞色素p450酶系统的活性的炎症诱导抑制而产生的不良药物反应(kimth等人,《febs杂志(febsj)》,第278卷,第13期,第2307到2317页(2011年))。结果展示,在炎症性肠-肝串扰中对i阶段和ii阶段代谢酶的mrna表达改变。因此,对药物药代动力学和药效学的准确预测需要考虑多器官相互作用以及生理情境(即,健康对疾病)。对于具有窄治疗窗的药物来说,这是特别相关的,因为甚至是细胞色素p450活性的重度变化都可能促生毒性。5.肠-肝整合系统中的细胞因子水平偏离单独的隔离系统的线性和。在刺激后的6小时、24小时和72小时在培养基中对所分泌细胞因子和趋化因子的水平进行了测量以检查炎症性反应的时间演化。对72小时细胞因子测量执行了成对分级聚类以探索分析物与状况之间的细胞因子反应的相关性。无监督主成分分析(pca)揭示,细胞因子数据集中超过96%的协方差可以被前2个主成分捕获。pc1占数据变化性的76.5%,分离相互作用对隔离对照物;pc2占总变化性的19.8%并且区别仅肠状况和仅肝状况。加载绘图描绘了每个分析物对地1和第2主成分的相对贡献。所有分析物都被主动加载到pc1上并且贡献于整合系统中的细胞因子水平,而对pc2的加载可以有助于推断整合系统中的循环细胞因子/趋化因子源的原代组织。虽然没有任何可溶因子对于肠或肝来说是唯一的,但是多元细胞因子模式可以揭示组织特异性特征。为了准确评估mps间串扰对整合炎症性反应的贡献,将相互作用系统中的所测量细胞因子水平与隔离状况的理论线性和进行比较。在隔离情况下观察到的细胞因子水平说明了由于直接tlr4活化和mps内旁分泌信号传递而产生的细胞因子输出。整合系统中的实际(所测量)细胞因子水平显著偏离隔离系统的线性和,从而揭示由于mps间通信而产生的对细胞因子产生的非线性调节。约58%的分析物是线性可加的,23%是负可加的,并且19%是正可加的,因此一些非常明显。有趣的是,若干细胞因子展现出与在高达24小时内是线性可加的cxcl6类似的时间动态,并且然后背离线性和并且变为正可加。这可能暗示阈值相关的调控,其中细胞因子产生取决于器官串扰期间的上游诱导物分子的积累。6.炎症相关cxcr3配体在肠-肝相互作用中大大扩增。表示出了cxcr3配体的显著正添加扩增,其中cxcl10(ip10)和cxcl11(i-tac)为最显著正添加并且cxcl9(mig)为边界显著。相比于单独的肠和单独的肝的水平的线性和,就肠-肝mps中的水平来说,总分析物中具有可加性、次可加性和正可加性的部分分别为58%、23%和19%。已经在自身免疫、移植排斥、感染、和癌症中蕴含了cxcr3信号传递(groomjr等人,《免疫细胞生物学(immunolcellbiol)》,第89卷,第2期,第207到215页(2011年);singhup等人,《内分泌代谢和免疫失调药物靶标(endocrmetabimmunedisorddrugtargets)》,第7卷,第2期,第111到123页(2007年))。表13统计上不同于线性和(调整后p值<0.05)的细胞因子/趋化因子以及相应受体。这些结果显示,对于评估全身性炎症过程及其对疾病发展的潜在影响来说,考虑肠-肝串扰可能很重要。rna测序数据显示出在器官串扰期间在肠和肝两者中对ifnα/β/γ信号传递途径的活化。tnfα可以扩大cxcr3配体的ifn相关产量。pca加载揭示,tnfα主要由肠衍生,并且ifnγ由肠和肝两者以相当水平产生。似是而非的是,肠(树突细胞)衍生的tnfα与组织特异性ifnγ信号传递相互作用以驱动肠和肝两者中的cxcr3配体产生。然而,难以确定上皮和免疫隔室对整合反应的相对贡献。尽管免疫细胞由于如表所示的tlr4的较高表达而是内毒素的主要响应者,所以上皮细胞还通过借助于免疫细胞衍生的细胞因子如tnfα和il-1进行的活化而对炎症有间接贡献(nguyentv等人,《药物代谢与处置(drugmetabdispos)》,第43卷,第5期,第774到785页(2015年);yeruvas等人,《国际结肠直肠疾病杂志(intjcolorectaldis)》,第23卷,第3期,第305到317页(2008年);dwinellmb等人,《胃肠学(gastroenterology)》,第120卷,第1期,第49到59页(2001年))。表14tlr表达(log10,归一化到gapdh)使鼠肝细胞在体外暴露于tnfα和ifnγ促进cxcl10mrna和蛋白质表达(hassanshahig等人,《伊朗过敏、哮喘和免疫学杂志(iranjallergyasthmaimmunol)》,第6卷,第3期,第115到121页(2007年))。已经显示,il-1α/β、tnfα和ifnγ的组合在肠细胞系和人肠异种移植物中诱导cxcr3配体基因表达和蛋白质分泌。为了评估上皮对细胞因子反应的贡献,添加了5ng/mltnfα、5ng/mlifnγ或两者使其存在24小时以在基底刺激肠上皮(caco2-bbe/ht29-mtx)。在不存在免疫细胞的情况下,tnfα和ifnγ对肠上皮的共同治疗引起在整合系统中识别的8个趋化因子中的4个趋化因子显著扩增,所述四个趋化因子包含cxcl9、cxcl10、cxcl11和cx3cl1(表11)。这些结果证实rnaseq发现并且显示ifnγ和tnfα信号传递串扰对于整合系统中的趋化因子产生起到中心作用。这些结果显示,上皮细胞在炎症性肠-肝串扰期间不是被动旁观者,而是通过与免疫细胞的旁分泌相互作用显著贡献于整体免疫环境。在炎症性肠-肝相互作用下,从所公开的整合肠-肝mps中检测到趋化因子产量的正可加扩增。这种扩增部分地通过tnfα和ifnγ信号传递介导。尽管免疫细胞通常被视为内毒素的主传感器,但是此处的结果显示,上皮细胞对免疫细胞衍生的信号进行响应以影响cxcl9/10/11和cx3cl1趋化因子产量。暴露于tnfα和ifnγ并不产生肠上皮细胞中的ccl19、ccl20、cxcl6和ccl2扩增,这指示涉及可能在不同的细胞类型中涉及额外机制。在整合系统中观察到的趋化因子产量可以靶向先天和适应性免疫系统(表)。可以基于趋化因子和相应受体剖面推断潜在免疫细胞募集。尽管在系统中未表示适应性免疫,但是在器官串扰期间,对连接先天和适应性免疫的途径的调控是明显的。例如,识别了cd40共刺激过程的富集。cd40是在免疫细胞以及非免疫细胞上泛表达的表面受体。cd40l主要由cd4+t细胞表达,并且cd40-cd40l接合介导异源性细胞通信(daneses等人,《肠(gut)》,第53卷,第7期,第1035到1043页(2004年))。合在一起,cxcr3趋化因子产生和cd40-cd40l调控暗示朝着th1信号传递的偏置。实例4。用于药代动力学/药效学(pk-pd)预测的4向mps芯片。(1)4向mps存活且具功能性持续至少2周。材料和方法验证:十三个4向mps平台(每个平台n=9个泵)中的流速在没有校准的情况下从0.82μl/s到1.12μl/s求平均,并且具有0.07μl/s的平均标准偏差。根据流速测量结果计算的软件校准系数并将其输入以将泵送速率校正为处于目标流速的±5%内。在实验期间使用qmix=5ml/天的系统相互作用流速。基于到人的每种组织类型的心输出量的相对百分比将流量从混合器分割到每个mps;这些数字可以容易地在用于不同缩放策略和mps模块的平台上修改。另外,mps内基部再循环速率0.25μl/s(肠mps、肺mps和子宫内膜mps)和1μl/s(肝mps和混合器)用于将良好混合的基部培养基提供到每个隔室中并氧化肝组织。每48小时进行完全培养基改变。在培养基改变期间,在两周相互作用研究内,从每个隔室中采集样本以评估mps功能。在4向mps的2周共培养期间测量健康细胞功能的生物标记度量:在流量分割情况下,肝、肠、肺和子宫内膜。每两周,从条件培养基中测量分泌的白蛋白和igfbp-1。用使用商业endohm系统在平台下测量的跨上皮细胞电阻(teer)量化肠mps和肺mps的屏障完整性。同时,监测每个隔离的mps的功能。结果1.4向mps在至少两周内支持细胞生存力和功能。来自4向mps平台研究的连续功能度量指示2周培养期间的多器官mps生存力。通过实验得出结论,观察短暂的白蛋白分泌动力学,最初白蛋白分泌增加,之后逐渐减少。用跨上皮细胞电阻(teer)量化肠mps和肺mps的屏障完整性。来自肠mps的teer值在实验的剩余部分沉降到150ω·cm2到250ω·cm2范围之前在相互作用研究的早期波动。肺mpsteer值在前几天跟随高teer的类似趋势,但是最终建立600ω·cm2到800ω·cm2范围内的稳定值。由胰岛素样生长因子-结合蛋白1(igfbp-1)的分泌评估的子宫内膜mps功能在整个研究期间保持在20pg/天到30pg/天范围内。观察隔离研究中每个表型度量的类似趋势,然而隔离子宫内膜mps(平台下)中的igfbp-1分泌速率低于相互作用研究的igfbp-1分泌速率。2.来自一个器官的内源产生的白蛋白以受控的系统流速均匀地分布到每个隔室。在4向mps平台中,通过从每个隔室中采集样本来表征系统流速(qmix)对白蛋白(由肝mps内源地产生)分泌和分布动力学的影响,并且然后从计算上对所述结果进行建模以评估分布的准确性。每个隔室和混合室中的白蛋白浓度在第2天(qmix=5ml/天),在第4天(qmix=15ml/天)以及在第6天(qmix=30ml/天)。随着系统流速增加,如由实验测量结果证明的,白蛋白被更均匀地分布,其中在更高流速的情况下,mps之间的偏差相当低。类似地,所计算的白蛋白分泌速率示出更小的标准偏差。然而,在第2天到第4天内一个平台示出了在所有隔室中相当低的白蛋白。此外,将计算上产生的白蛋白分布轮廓与实验上测量的白蛋白浓度进行比较。两个值之比指示在4向mps平台中更高的流速导致更确定的分子生物分布。(2)肠-肝/肺/子宫内膜4向平台:独立的流速控制改进用于复杂生理学的pk-pd预测。背景在人现代医学研究中,出于人治疗前景,对动物研究结果的解释通常采用异速比例;并且出于体内疗效的前景,对体外结果的解释通常被称为iviv相关性和外推法。肝mps药代动力学(pk)的体外研究通过考虑结合培养基、药物摄取和消除来表征。体内研究使用已知的临床数据和基于生理学的pk(pbpk)/吸收/结合模型来计算可比较参数。将来自manvelian等人,2012年;shimamoto等人,2000年;yilmaz等人,2011年;willis等人,1979年的七种药物的临床pk数据与从liverchiptm处收集的体外肝数据进行比较以评估来自liverchiptm的体外结果的预测准确性。尽管每个细胞的用于游离双氯芬酸消除的体内pbpk具有速率常数1.76x10-10(细胞*分钟)-1,但是来自liverchiptm的缩放的肝mps被研究以示出双氯芬酸消除速率常数5.66x10-9(细胞*分钟)-1。因此,来自liverchiptm的体外药物pk数据过高估计了体内药物消除速率。材料和方法在图示和流量分割之后(总qmixing=1μl/s;肝/混合器再循环=1μl/s;肠/肺/子宫内膜再循环速率=0.5μl/s)(9个泵送的流量:5个自循环,4个混合;6个独立的流速:5个自循环流量坍塌到2个独立气动工作周期;6个泵组=18dof),4向mps相互作用组被研究,其中向混合室添加说明静脉内计量的药剂,同时向肠室添加说明口服量的药剂。在实验中将药物添加到肠室的顶面。结果均匀药物分布被计算为下游(子宫内膜)mps达到混合室中的浓度的90%的时间。药物暴露被计算为下游(子宫内膜)mps中0到48小时的曲线下面积(auc)。药物暴露和分布能够强烈驱动选择有用的操作范围:对于药物渗透大于10-6cm/s而言,qmixing>15ml/天;并且对于0到48小时的auc大于2*104ng/l*hr而言,qmixing>40ml/天。实例5。7向mps芯片的操作材料和方法验证:十个7向平台(每个平台n=17个泵)的流速平均为1.12±0.10μl/s。根据流速测量结果计算的软件校准系数并将其输入以将泵送速率校正为处于目标流速的±5%内(0.99±0.056μl/s)。7向mps平台以与实例4中描述的4向平台相同的方式使用和操作。7向平台包含肠(免疫活性)mps、肝(免疫活性)mps、肺mps、子宫内膜mps、心mps、脑mps和胰腺mps,并且在3周时期内对存活性和功能进行评估。在相互作用研究之前,每个mps被隔离地分化或熟化。在流量分割情况下,平台以系统流速qmix=10ml/天运行。在培养基改变期间,基部共同培养基用于肠mps、肺mps、肝mps和子宫内膜mps,而新mps被供应有其优选维持培养基。然后允许每个基部培养基在整个相互作用过程中混合,培养基以48小时间隔改变。每2到4天,最多3周评估每个mps的功能,与隔离的mps进行比较以基准化无相互作用mps的功能。由于隔离的胰腺mps的功能急剧下降,对于相互作用研究和隔离研究两者,在第12天用新岛代替岛。结果1.7向mps在至少三周内支持细胞生存力和功能。建立短暂的白蛋白分泌动力学、持续的肠和肺teer值以及igfbp-1分泌轮廓。在研究期间良好保持通过拍频监测的心mps的功能。n-乙酰天冬氨酸(naa)和c-肽释放轮廓揭示了在多达3周脑mps和胰腺两者也是功能性的。相互作用结果与隔离结果的比较示出了相互作用对mps功能没有负面影响。在相互作用期间观察到增加的naa分泌和更持久的c肽分泌。在7向mps平台中可以维持长期mps生存力和功能持续至少三周延长的培养时期。2.“口服”施用的药物及其代谢物以与模型药代动力学预测一致的浓度跨mps分布。使用临床上相关的浓度进行的外源药物研究对将体外结果转化成临床成果来说是重要的。在7向mps平台中分析双氯芬酸、即非类固醇抗炎药的药代动力学。体内的口服双氯芬酸的最大测量血浆浓度cmax在2μm与6μm之间变化(daviesnm等人,《临床药代动力学(clin.pharmacokinet.)》,第33卷,第184页到213页(1997年))。4'-羟基-dcf(4-oh-dcf)是dcf的常见代谢物。为了从平台中的口服给药中概括临床上观察到的cmax,将双氯芬酸添加到肠mps的顶面。dcf的测量浓度以及跨不同mps隔室的4'-羟基-dcf培养基符合对应药代动力学模型预测。dcf剂量跨肠上皮屏障被吸收,分配到肝mps并且随后分配到混合室和所有其它mps隔室。代谢物4-oh-dcf在肝mps中产生,跨7向mps平台循环,并且在所有其它隔室中被检测到。对dcf和4-oh-dcf浓度的基于生理学的药代动力学(pbpk)模型预测与测量的数据良好的一致,这指示平台功能以确定性方式与生物学预测一致。未结合内在清除率(cl_int(u);即,在缺乏流动时肝移除药物的能力)被估计为13.90μl/分钟,并且这种清除的大约19%被估计为朝向形成4-oh-dcf代谢物。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1