量子存储器和相关应用的制作方法
1.本发明涉及一种量子存储器。本发明还涉及一种相关的量子计算机。本发明还涉及一种用于在量子存储器中存储信息的方法。本发明还涉及一种相关的用途。本发明进一步涉及一种用于筛选多肽的方法。
背景技术:
2.经典计算依赖于对表现出多种状态的多个实体进行计算。在这样的经典背景下,这样的物理实体被命名为“位”、并且仅具有两种状态,通常命名为0和1。通过操作每个位的两种状态来进行计算。
3.相比之下,量子计算依赖于对被称为量子位(量子比特的简称)序列的一系列物理实体进行的操作。虽然每个量子位也像比特一样具有两种状态,但量子力学允许每个量子位同时处于两种状态的相干叠加。因此,通过为每个物理实体操作两种以上的状态来进行计算,使得依赖于此类计算的算法具有更高的潜力。
4.任意两级量子力学系统均可被用作比特。若能够单独操作两种状态,也可以使用多级系统。
5.作为量子力学系统的不完整列表,我们可以引用光子、离子、电子、原子核、约瑟夫森结或量子点。
6.作为特定示例,对于光子,多个向量可以被用作状态。值得注意的是,已经进行了基于作为状态(存在一个光子或不存在光子)的光子数量或基于光子的偏振状态的实验。
7.然而,量子退相干限制了此类量子力学系统的量子行为。量子退相干对应于量子力学系统与环境的相互作用,特别是环境残余噪声和与环境粒子的碰撞。
8.由于环境噪声和碰撞概率随温度而降低,此类量子力学系统常常在低温环境下使用。
9.这意味着这些系统均不适于在生理温度(即,35℃至40℃,尤其是36℃至38℃之间的温度)下运行。
技术实现要素:
10.因此,需要一种适用于在生理温度下操作的量子存储器。
11.最后,本说明书提出一种量子存储器,所述量子储存器包括存储空间所述存储空间包括量子位,至少一个所述量子位包括多肽或至少一种多肽、由多肽或至少一种多肽构成或为多肽或至少一种多肽,所述多肽或所述至少一种多肽包括至少一个阿尔法螺旋二级结构。
12.根据本发明的有利但非强制性的其他方面,所述量子存储器可以结合一个或多个如下特征,采用技术允许的任何组合:
13.作为包括至少一个阿尔法螺旋二级结构的多肽的量子位的数量与量子位总数之间的比率为大于或等于70%。
14.‑
每个量子位均为包括至少一个阿尔法螺旋二级结构的多肽。
15.‑
所述量子存储器还包括写入单元,所述写入单元适用于将数据写入所述存储空间中;以及读取单元,所述读取单元适用于读取存储在所述存储空间中的数据。
16.‑
所述写入单元为电子电路。
17.‑
所述读取单元为电子电路。
18.‑
所述多肽选自由普兰奈辛(plannexin)(序列号:1)、fgl(序列号:2)、肽p2(序列号:3)、双翅菌肽(diptericin)b(序列号:4)、pten
‑
pdz(序列号:5)、gnbp样3(序列号:6)、肽6(序列号:7)、ptd4
‑
pi3kac(序列号:8)、物质p(序列号:9)、生长抑素(序列号:10)、血管活性肠肽(序列号:11)、垂体腺苷酸环化酶激活多肽(序列号:12)、降钙素基因相关肽(序列号:13)、胃饥饿素(序列号:14)、肥胖抑制素(序列号:15)、神经肽s(序列号:16)、以及可卡因和安非他明调节转录肽(序列号:17)组成的组;
19.‑
所述存储空间包括少于100个量子位,优选为少于20个量子位;及
20.‑
所述存储空间包括多个相同的量子位对。
21.本说明书还涉及一种设备,特别是一种量子计算机,所述量子计算机包括至少一个如前所述的量子存储器。
22.本说明书还描述了一种用于在量子存储器中存储数据的方法,所述量子存储器包括存储空间,所述存储空间包括量子位,至少一个量子位为包括至少一个阿尔法螺旋二级结构的多肽。所述方法包括将数据写入所述存储空间中,以及读取在所述存储空间中的所述数据。
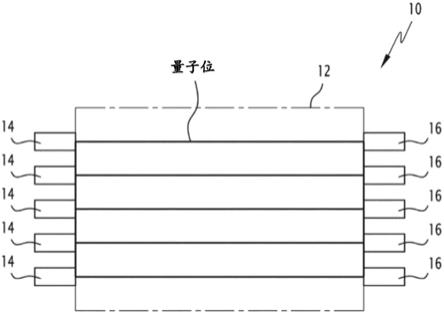
23.本说明书还涉及一种包括至少一个阿尔法螺旋二级结构的多肽的使用,所述多肽用于改善与其他器官协同运行的器官的量子存储器的性能,所述运行至少部分建模为量子存储器。
24.根据本发明的有利但非强制性的其他方面,所使用的所述多肽可以结合一个或多个如下特征,采用技术允许的任何组合:
25.‑
所述器官为大脑,以及
26.‑
通过评价所述器官的记忆能力、所述器官的学习能力以及所述器官的监察能力(reconnaissance)构成的列表中选择的至少一个标准来评价所述量子存储器的性能。
27.本说明书还描述了一种用于筛选多肽的方法,所述多肽对改善受试者的、与其他器官协同运行的器官的量子存储器的性能是有用的,所述运行至少部分建模为量子存储器,所述方法包括:
28.a)选择已知包括至少一个阿尔法螺旋二级结构的多肽,或者通过结构建模来确定包括至少一个阿尔法螺旋二级结构的多肽;
29.b)评估所述选择的、包括至少一个阿尔法螺旋二级结构的多肽是否改善了与其他器官协同运行的器官的量子存储器的性能,所述运行至少部分建模为量子存储器;以及
30.c)基于步骤b)的评估,选择对改善与其他器官协同运行的器官的量子存储器的性能有用的所述多肽。
附图说明
31.基于如下描述将更好地理解本发明,所述描述对应于附图且作为说明性示例给
出,但不限制本发明的目的。在附图中:
32.‑
图1示出了量子存储器的示例;
33.‑
图2示出了阿尔法螺旋二级结构;
34.‑
图3示出了构成阿尔法螺旋的分子网络的示意图,该示意图说明了所述分子网络可以分解为相互连接的氢键的三个一维子网;
35.‑
图4说明了视觉处理模型中涉及的途径:后鼻腔皮层(postrhinal cortex,por)通过平行的视觉途径从被称为上丘的进化上古老的脑干区域获取有关运动对象的信息。两条平行的视觉途径通过称为丘脑的大脑结构中的不同中继站进行路由:用于v1的dlgn(dorsolateral geniculate nucleus,侧膝状体背核),以及用于por的丘脑后结节(改编自加州大学旧金山分校的massimo scanziani实验室的图形)。
具体实施方式
36.图1示意性地说明了量子存储器10。
37.量子存储器是一种适用于存储各种数据的电子设备。
38.量子存储器10包括存储空间12、写入单元14以及读取单元16。
39.存储空间12为一组量子位。
40.在本示例中,量子位包括多肽或由多肽构成,该多肽包括至少一个阿尔法螺旋(α
‑
螺旋)二级结构。
41.多肽链由一系列通过肽键连接的氨基酸(aa)残基
‑
nh
‑
cα
‑
co
‑
形成。多肽中的氨基酸数量至少为两个,并且可以包括数百或数千个氨基酸。对于包括至少一个α
‑
螺旋的多肽,该多肽必须为至少为4个氨基酸长。优选地,该多肽至少为10、20、30、40或50个氨基酸长。在某些实施方式中,该多肽包括至多1000个氨基酸,例如,最多500、400、350、300、250、200、150、100或50个氨基酸。
42.基于多肽的一级结构(即,构成多肽链的氨基酸序列),本领域技术人员通过使用蛋白质结构预测软件,如pep
‑
fold3(http://bioserv.rpbs.univ
‑
paris
‑
diderot.fr/services/pep
‑
fold3/)或psipred 4.0(预测二级结构,http://bioinf.cs.ucl.ac.uk/psipred_new/)等开源软件可以很容易地确定该多肽是否包括二级结构,即,α
‑
螺旋和/或β
‑
片层。
43.根据一实施方式,包括至少一个α
‑
螺旋二级结构的多肽进一步包括β
‑
片层。根据另一实施方式,包括至少一个α
‑
螺旋二级结构的多肽不包括β
‑
片层。在包括至少一个α
‑
螺旋二级结构的多肽中β
‑
片层的存在与量子位状态无关。
44.在这种情况下,量子位可以解释为自旋部分,并且作为自旋部分的支撑物或支架。换而言之,包括至少一个阿尔法螺旋(α
‑
螺旋)二级结构的多肽是对应于量子运行的量子效应的位置,所述多肽为量子位或为量子位的一部分。
45.在所描述的示例中,作为包括至少一个阿尔法螺旋二级结构的多肽的量子位的数量与量子位总数之间的比率为大于或等于70%。
46.在这样的示例中,量子位的每个状态都是对应于c=o键的拉伸的酰胺
‑
i模式的振动状态。
47.替代地或补充地,量子位的每个状态都是对应于n
‑
h键的拉伸的酰胺
‑
a模式的振
动状态。
48.包括至少一个阿尔法螺旋二级结构的多肽的示例包括普兰奈辛(序列号:1)、fgl(序列号:2)、肽p2(序列号:3)、双翅菌肽b(序列号:4)、pten
‑
pdz(序列号:5)、gnbp样3(序列号:6)、肽6(序列号:7)、ptd4
‑
pi3kac(序列号:8)、物质p(序列号:9)、生长抑素(序列号:10)、血管活性肠肽(序列号:11)、垂体腺苷酸环化酶激活肽(序列号:12)、降钙素基因相关肽(序列号:13)、胃饥饿素(序列号:14)、肥胖抑制素(序列号:15)、神经肽s(序列号:16)、以及可卡因和安非他明调节转录肽(序列号:17)。这些多肽与认知过程(特别是学习和记忆过程)的维持、恢复或改善有关。
49.一组量子位包括少于100个量子位,优选为少于20个量子位。这意味着存储空间12中的量子位总数低于或等于100个量子位,优选为低于或等于20个量子位。
50.在一具体实施方式中,该组量子位包括多于2个量子位,优选为多于10个量子位。
51.根据详细的示例,该组量子位包括若干个相同的量子位对。这为存储空间12提供了对称性。
52.写入单元14适用于在每个量子位上写入数据。
53.更确切地说,写入单元14适用于在至少一个量子位的状态组合中转换数据。
54.这特别意味着写入单元14适用于改变所述量子位的振动状态。
55.例如,写入单元14为一个电应用单元。
56.在这种情况下,所述电应用单元适用于以电脉冲的形式在一个或多个量子位中编码数据
57.电脉冲适用于改变至少一个量子位的振动状态,以使得至少一个量子位的振动状态对应于待存储的数据。
58.更普遍地,写入单元14为电子电路。
59.读取单元16适用于读取每个量子位的振动状态。
60.这特别意味着读取单元16适用于确定量子位的振动状态。
61.例如,读取单元16为电压表。
62.在这种情况下,电压表能够来检测量子位的电压。
63.每个量子位的电压能够来确定每个量子位的状态。
64.更普遍地,读取单元16为电子电路。
65.得知每个量子位的状态,能够来访问存储在存储空间12中的数据。
66.现在,参考在量子存储器10中存储数据的方法来描述量子存储器10的运行。
67.存储方法包括写入步骤和读取步骤。
68.在写入步骤中,通过写入单元14将数据写入所述量子空间12。
69.量子存储器10存储数据,更具体地,量子空间12存储数据。
70.附录中提供了将阿尔法螺旋构型中的多肽的信息从写入单元14转移到读取单元16的能力的解释。
71.当期望访问数据时,通过使用读取单元16执行读取步骤。
72.读取单元16测量量子空间12的量子位数值。
73.在当前情况下,该数值以伏特表示。
74.数值能够访问每个量子位的状态。
75.每个量子位的状态随后由读取单元16转换为待存储的数据。
76.因此,量子存储器适用于在量子存储器10中存储数据,而无需冷却外壳。
77.这种量子存储器10,更确切地说,包括至少一个阿尔法螺旋构造的多肽的量子特性可被用于多种应用。
78.作为第一种应用量子存储器10用于量子计算机。
79.作为第二种应用,建议将包括至少一个阿尔法螺旋二级结构的多肽用于改善与其他器官协同运行的器官的量子存储器的性能,所述运行至少部分建模为量子存储器。
80.作为一具体示例,所述器官为大脑,尤其为视觉皮层,更具体为后鼻腔皮层。
81.通过评价器官的记忆容量来评价量子存储器的性能。
82.替代地,将使用另一性能标准,例如器官的学习能力或器官的监察能力。
83.对于性能标准,也可以考虑任一种性能标准的组合。
84.因此,第二种应用还涉及包括至少一个阿尔法螺旋二级结构的多肽的使用以用于改善实验受试者的记忆能力、学习能力和/或监察能力。所述第二种应用还涉及一种用于改善有需求的受试者的记忆能力、学习能力和/或监察能力的方法,包括向受试者施用包括至少一个阿尔法螺旋二级结构的多肽。
85.受试者特别为哺乳动物或非哺乳动物,更具体为啮齿动物、猫科动物或灵长类动物,例如猴子或人。
86.根据一实施方式,通过结构建模,例如通过使用蛋白质结构预测软件,多肽先前已经被表征为包括一个阿尔法螺旋二级结构,并且基于这种表征而被选择。
87.根据另一实施方式,所述多肽先前已经被表征为具有与大脑的一个或多个特定区域(例如,视觉皮层或后鼻腔皮层)结合的能力,并且基于这种表征而被选择。
88.根据另一实施方式,通过常规用于评价认知功能的测试,所述多肽先前已经被表征为具有刺激认知功能的能力,详情如下。
89.在所述第二种应用中,包括至少一个阿尔法螺旋二级结构的所述多肽优选不同于普兰奈辛(序列号:1)、fgl(序列号:2)、肽p2(序列号:3)、双翅菌肽b(序列号:4)、pten
‑
pdz(序列号:5)、gnbp样3(序列号:6)、肽6(序列号:7)、ptd4
‑
pi3kac(序列号:8)、物质p(序列号:9)、生长抑素(序列号:10)、血管活性肠肽(序列号:11)、垂体腺苷酸环化酶激活多肽(序列号:12)、降钙素基因相关肽(序列号:13)、胃饥饿素(序列号:14)、肥胖抑制素(序列号:15)、神经肽s(序列号:16)、以及可卡因和安非他明调节转录肽(序列号:17)。
90.第三种应用涉及一种用于筛选多肽的方法,该多肽对改善受试者的、与其他器官协同运行的器官的量子存储器的性能是有用的,所述运行至少部分建模为量子存储器,所述方法包括:
91.a)选择已知包括至少一个阿尔法螺旋二级结构的多肽,或者通过结构建模来确定包括至少一个阿尔法螺旋二级结构的多肽;
92.b)评估选择的、包括至少一个阿尔法螺旋二级结构的多肽是否改善了与其他器官协同运行的器官的量子存储器的性能,所述运行至少部分建模为量子存储器;以及
93.c)基于步骤b)的评估,选择改善与其他器官协同运行的器官的量子存储器的性能有用的多肽。
94.所述受试者特别为哺乳动物或非哺乳动物,更具体为啮齿动物、猫科动物或灵长
类动物,例如猴子或人。
95.如上所述,通过使用蛋白质结构预测软件(例如,pep
‑
fold3或psipred4.0)可以确定多肽包括至少一个阿尔法螺旋二级结构。
96.例如,可以通过使用通常用于评价认知功能的测试来评估所选择的、包括至少一个阿尔法螺旋二级结构的多肽是否改善了与其他器官协同运行的器官的量子存储器的性能,所述运行至少部分建模为量子存储器。这些测试包括用于测定海马体依存性认知功能的测试,例如评价视觉空间记忆的morris水迷宫(morris r.j neurosci methods.1984;11:47
‑
60)、评价空间记忆的t迷宫(gentry g,brown wl,lee fi.,j comp physiol psychol.1948;41:312
‑
318)、评价工作记忆的径向迷宫(olton ds,samuelson rj.j exp psychol anim behav process.1976;2:97
‑
116)和评价联想记忆的被动回避测试(van der poel am.acta physiol pharmacol neerl.1967;14:503
‑
505;azami ns,piri m,oryan s等,neurobiol learn mem.2010;93:455
‑
162)。这些测试还包括用于测定海马体独立认知功能的测试,例如评价辨别记忆的物体识别(rothblat la,kromer lf.behav brain res.1991;42:25
‑
32)和评价联想记忆的条件味觉厌恶(krai pa,beggerly hd.physiol behav.1973;10:145
‑
147;buresova o,bures j.acta neurobiol exp(wars).1973;33:689
‑
698;martinez
‑
moreno a,rodriguez
‑
duran lf,escobar ml.front behav neurosci.2011;5:61)。其他可用的测试包括评价焦虑状态的高架十字迷宫(pellow s,chopin p,file se等,j neurosci methods.1985;14:149
‑
167)和评价应激状态的自发活动、唤醒、用于应激状态线索的开放式现场测试(denenberg vh,whimby ae.science.1963;142:1192
‑
1193;gonzalez
‑
burgos i,cuevas
‑
alvarez l.physiol behav.1992;52:1207
‑
1209)。
97.若在步骤b)中,多肽被显示改善了与其他器官协同运行的器官的量子存储器的性能,则多肽被认定或选择为对用于改善与其他器官协同运行的器官的量子存储器的性能有用的。
98.在一具体示例中,器官为大脑,尤其为视觉皮层,更具体为后鼻腔皮层。
99.通过评价器官的记忆容量来评价量子存储器的性能。
100.替代地,可以使用另一性能标准,例如器官的学习能力或器官的监察能力。
101.也可以考虑任一种性能标准的组合作为性能标准。
102.当技术相关时,可以将先前所描述的实施方式结合,以形成另一实施方式。
103.附录
104.多肽链由一系列通过肽键连接的氨基酸残基
‑
nh
‑
c
α
‑
co
‑
构成,其中c
α
是侧链的碳载体。由于肽键的刚性,链的结构表征为co
‑
nh酰胺基团的不同平面的连续取向。
105.在链中氢键的建立对其结构的稳定性起决定性作用,并且当同一蛋白质残基之间建立的键数最大时,会出现两个主要的二级结构:β
‑
片层和α
‑
螺旋。
106.α
‑
螺旋二级结构对应于链的周期性缠绕而成为直螺旋,使得α
‑
螺旋是准一维(1d)结构。
107.β
‑
片层由几个β
‑
链构成,β
‑
链为通过氢键网络保持在一起的多肽链的拉伸片段。与α
‑
螺旋不同,β
‑
片层由蛋白质链之间的氢键形成,而不是在一条链内。
108.α
‑
螺旋是通过每螺旋弯转3.6个氨基酸残基的卷绕和每个残基沿着螺旋轴平移
1.5埃或每转平移5.4埃形成的结构(图2)。在这样的构型中,每个肽残基的羰基通过氢键与下一个第四残基的胺基基团结合。当考虑到围绕刚性肽键为中心的酰胺基团时,每个酰胺基团都通过氢键与第三个下一酰胺基团结合。因此,α
‑
螺旋可以表示为三个氢键链的嵌套(图3)。
109.在传统表示方法中,上部c
α
碳与从酰胺基团向右移动的下部n氮直接相关。因此,一方面,每个酰胺基团通过c
α
碳与两个最近的相邻酰胺基团连接,另一方面,每个酰胺基团通过氢键与第三个最近的相邻酰胺基团连接。因此,α
‑
螺旋的分子网络可以分解为相互连接的氢键的三个一维子网(图3)。
110.可以通过两种主要模式来实现α
‑
螺旋中振动能量的传输:本质上对应于c=o键的拉伸的酰胺
‑
i模式和与n
‑
h键的拉伸相关的酰胺
‑
a模式。这种传输主要为非线性现象,一方面与分子内非谐性部分有关,另一方面与螺旋骨架的低频模式强耦合有关。在这种情况下,控制螺旋结构的氢键起主导作用。事实上,由于氢键连接同一子网的酰胺基团,它促进了每个子网中非线性效应的出现。
111.因此,作为第一种方法,可以通过将α
‑
螺旋中的动力学建模为包含由一组氢键连接的n酰胺基团的一维网络来研究单个子网内的振动传输。
112.这种研究揭示了由现在所描述的三种物理现象之间的竞争所导致的某种复杂性。
113.第一种物理现象是酰胺基团与氢键之间的偶极耦合。这对应于振动激子在网络中作为一系列平面波的传播。这种激子被称为振动子,并且可以被解释为与分子内振动的集体动力学相关的准粒子。由于偶极耦合,这种振动子因此是离域的。
114.另一种物理现象是由于每种模式的分子内势的非谐性的束缚态。这些状态由距离间振动子的位置限定,使得振动子相互被陷俘(trapped)。在平行的情况下,通过平移的不变性导致类似平面波的振动子的质心离域,这些束缚态表现为类似非线性系统中的经典孤子。
115.所包括的第三种物理现象是高频内部模式与低频外部振动(声子)之间的相互作用。这种相互作用改变了振动子状态的性质。通过这种相互作用,振动子的存在引起网络的局部变形,该变形立即跟随振动子的传播。由变形修整的振动子形成“小极化子”。这种包层机制(即,传统自陷的量子等价物)扰乱了两个过程中的振动动力学。首先,它降低了跳位常数、并且降低了极化子的离域能力。其次,它是非线性的第二个来源,并且促进了负责新构成特定束缚态的极化子之间的非局部相互作用。
116.通过在对声子的热力学平衡系统的框架中模拟这三种现象之间的这种竞争,似乎可以考虑为该系统由广义的哈伯德的玻色子哈伯德量来描述。
117.这种形式可以示出α
‑
螺旋构型在每一组酰胺基团的水平上都发出振动子。振动子优先与与酰胺基团的集体外部运动相关的低频声子相互作用。
118.此外,借助如前所述的一维模型,根据第二种物理现象与第三种物理现象之间的竞争可以形成两类束缚态。低频束缚态形成低于自由态连续体的带,并且主要表征位于同一位置的两个极化子。相比之下,高频束缚态能够与连续体共振,并且对应于两个极化子被陷俘于两个最近的相邻位置的情况。
119.除平均场近似之外,对极化子声子耦合影响的分析示出了双极化子态的弛豫发生在皮秒级时间内。
120.此外,低频束缚态进行向高频束缚态的转变。相比之下,高频束缚态在低频束缚态和自由态连续体中都会衰减。最后,在声子的作用下,自由态在与其相关的连续体中发生转变。
121.时间分辨泵探测光谱(time
‑
resolved pump
‑
probe spectra)的计算示出了束缚态具有特定特征。事实上,自由态有助于负峰的形成,而束缚态有利于正峰的存在。由于这种峰的位置表示探测到的束缚态的能量,对分子内振动的集体非线性性质的考虑对于正确理解光谱至关重要。
122.通过使用三维模型,而不是一维模型,还显示出两个动态状态之间存在转换。
123.在低温下,互联网络调位常数起着至关重要的作用,并且极化和双极化状态具有重要的三维特性。
124.在生物温度下,修整机制促进网络内跳位,使得极化子只能在单个子网络中传播。量子态失去了其三维特性,因此可以用一维模型很好地描述。
125.这些特性使α
‑
螺旋构型中的多肽成为运输振动子的优良候选物质。
126.第一个理论被提出来支持这一论断。这种量子理论解释了能量是如何沿着蛋白质的α
‑
螺旋二级结构传输。该理论认为,蛋白质是仅由20个单体形成的生物聚合物。这些单体是与肽键结合以提供长多肽链的α
‑
氨基酸。这些螺旋缠绕的链具有准周期性结构,沿着该准周期性结构分布有通过氢键彼此连接的肽单元h
‑
n
‑
c=o。由于蛋白质的几何形状,这些肽单元形成三个平行于螺旋轴的分子链。因此,特定的链代表由一系列包含c=o基团的肽单元形成的一维分子网络。从振动的角度来看,c=o键的拉伸表征了被称为酰胺
‑
i模式的正常高频模式。
127.这一理论赋予c=o模式特殊的角色。事实上,当atp(adenosine triphosphate,三磷酸腺苷)分子与蛋白质的特定位点结合时,它会与介质中的水分子相互作用并且释放能量,通过共振耦合将能量转化为特定肽单元的c=o键的内部振动激发。相邻c=o基团之间的偶极的相互作用导致内部振动离域,并且在α
‑
螺旋能量传输的原点处形成振动激子。
128.第一个理论得到了改善,并且现在对应于孤子旁氏模型(soliton pang'smodel)。值得注意的是,已经示出了孤子的产生、传播和稳定性取决于激子
‑
声子哈密顿相互作用的对称性和激子能量应用的初始位置。
129.这些理论示出了,在α
‑
螺旋构型中,与c=o振动沿着多肽的传播相关的酰胺
‑
i振动子在运输由atp水解释放的能量和将化学能量转化为机械功中起着关键作用。
130.换而言之,这意味着可以在分子振动上对量子位进行编码。由振动量子传递信息。因此,振动激子(振动子)似乎是沿着分子网络(尤其是多肽)传输量子信息的优良候选物质。
131.这种振动传递的机制被考虑为是依赖于网络上的随机游走。
132.在详述这种理论的含义之前,先来概述网络上的随机游走以及此类概念在量子力学框架中的实现。
133.随机游走是一个理论概念,只需要两个元素:游走者和网络。游走者可以被解释为能够根据概率定律移动的移动物体。由一组连接的节点组成的网络起到游走媒介的作用。当网络的两个节点被连接时,两个节点之间就会建立联系,使得游走者可以从一个节点跳到另一个节点。
134.网络中随机游走这种概念能够描述一些已知现象,例如描述分子或原子在表面上运动的布朗运动或扩散现象。
135.这种概念也在算法中使用,特别是在蒙特卡罗算法中使用。
136.网络中随机游走概念在量子物理学中的类似模型被称为量子游走。在这种模型中,游走者不再被考虑为经典的移动体,而是被看作动力学受薛定谔方程(equation)控制的量子物体。
137.无论它们本质上是电子的,例如弗伦克尔激子(frenkel excitons),还是振动的,例如振动子,激子都遵循薛定谔方程作为这些元素作用于分子网络。激子的行为就像真正的能够传递能量甚至信息的量子游走者。
138.我们已经研究了在复杂分子网络上的耗散的激子量子游走,并且还分析了通过在局部声子存在的复杂分子网络上激子演化来进行量子信息传递的情况下的涌现性质。
139.为此,通过聚焦声子的存在提出了复杂分子网络的模型,更具体地说是聚焦在叠加的激子态传输的情况下激子
‑
声子缠结可能导致的量子退相干而提出了复杂分子网络的模型。
140.可以通过使用准退化微扰算子方法直接在系统的一般哈密顿量上运行来遵循这种方法。该方法通过表达编码两类修整过程的有效哈密顿量来允许对网络上的激子
‑
声子问题完全重新表述。
141.第一过程描述了通过虚拟声子云对激子的包裹。这种现象反映在激子本征态的重新定义中。
142.第二过程通过激子虚拟跃迁来代表声子的产生。在这个框架中,可以观察到对应于代表声子传输过程的新算子的声子的第二修整描述。换而言之,与描述局部振动的裸声子不同,修整过的声子能够重新定位、导致扩展模式的出现。
143.基于这种形式,可以推导出激子在这种系统中的行为。
144.特别是,对激子的修整本质上导致了对能量中未扰动激子特征值的重新定义。这种性质为激子表现得像极化子的这种事实的标志。换而言之,由于激子与围绕激子的声子云相互作用,激子不能被考虑为自由粒子。
145.新扩展声子模式的特性源自于网络的对称性。
146.这些特性使得解释激子动力学中观察到的所有奇异特征成为可能。在短时间内,极性行为解释了动力学急剧减速的原因。在长时间内,非马尔可夫动力学(non
‑
markovian dynamic)的出现与每个扩展声子模式的时间演化有关。特别地,这示出了激子特征值的知识是解释退相干演化的关键数据,也是信息量子超级重现的产生。这些结果表明,复杂网络的对称性可以在信息的生存中发挥重要作用。
147.这种方法的局限性在于当只要激子在分子网络上演化时,激子将自然倾向于受到其环境的影响。在分子网络的背景下,声子、由网络振动量化产生的量子粒子代表了激子与之相互作用最多的第一环境元素。因此,为了真实地描述激子量子游走,更准确的方法是将激子考虑为与环境相互作用的开放系统。
148.在这种情况下,我们已经进行了研究以量化这种环境在多大程度上会影响激子量子游走的顺利进行。这种分析示出了纯相移环境的存在可以显著改善激子网络传输的过程。值得注意的是,在激子量子游走的背景下,相干传播机制(自由演化)与非相干跳位过程
(与声子的能量交换)之间的微妙混合可以使超高效的混合量子传输的产生成为可能。因此,与人们可以想象的相反,现实激子传输框架中存在的量子退相干过程具有意想不到的有益效果。
149.在中短时间内,激子
‑
声子相互作用的影响基本上会导致量子退相干过程的出现。在这些条件下,激子系统会逐渐失去其发展叠加量子态的能力,并且最初编码的量子信息会被破坏。
150.然而,这种量子退相干过程已经示出了只是表面现象。事实上,激子
‑
声子系统永远不会完全丧失对其初始状态的记忆,使得信息可以在一段时间后重新出现。
151.已经在低温方案下观察到了这些强烈非马尔可夫行为的早期迹象。然后,令人惊讶的是,对这些行为的研究使观察到在低温和高温下出现的长期量子超级重现成为可能。
152.此外,量子重现周期似乎与温度无关,但依赖于耦合参数:温度不影响这些峰的出现周期,但主要是在短期内放大了重现的幅度。
153.超级重现的存在清楚地证明了非马尔可夫动力学可以在很长一段时间内找到最初编码的信息。换而言之,与人们可能想象的相反,退相干过程并非不可逆转。
154.换而言之,这意味着一致性的这种崩溃显然不是不可逆转的。事实上,只要等待很长一段时间,就会出现部分甚至全部的量子重现。
155.实际上,这种现象的物理起源与两个不同的因素有关:所研究网络有多小,以及声子脉动的退化。
156.当考虑到有限大小的网络时,声子环境的自由度的数量变得有限,使得在退相干因子的计算中起作用的各种相位项非常少。
157.此外,网络的对称性产生了布洛赫声子脉动(bloch phonon pulsations)的n≠1顺序的简并性:在随后的退相干因子的计算中只有两类相位项在起作用。
158.在短时间内,这些项将允许相移并且导致退相干因子下降。然而,在长时间内,减少的模式数量有利于时间窗口的出现,在此期间平均项可以再次在相中演变。
159.然后,这些重新定相的时刻产生了量子重现,即激子
‑
声子系统返回到接近其初始状态的状态。系统不会丧失对其过去状态的记忆:这是非马尔可夫行为的特征。
160.因此,在包括α
‑
螺旋的多肽中,尽管存在声子,但量子重现的发生使得信息的有效传输成为可能。
161.此外,我们观察到阿尔法螺旋中对应于n
‑
h键拉伸的振动状态胺
‑
a模式。edler等人(2004,physical review letters,vol 93,10,124
‑
127)示出了“稳定的阿尔法螺旋的n
‑
h模式的飞秒红外泵探测光谱发现了两个激发态吸收带,当螺旋展开时,这两个吸收带消失。与极化子理论的定量比较示出了这两个带反映了两类双振动束缚态,这两类双振动束缚态分别与同一位点处和最近相邻位点处的两个振动子的陷俘有关。后一种状态源自于螺旋中的声学声子,该声学声子与相邻位点相关联”。实验结果明确示出了这两个正带是螺旋构象的标志,在该螺旋构象中单个n
‑
h振动与声学声子相关。声子能够形成可以通过其非谐性识别的自陷态。当螺旋结构被破坏时,相关性消失,并且实验地观察到的泵
‑
探针响应是孤立振动子的响应。该理论将两个正峰归因于两种分别对应于在同一位点处和最近邻位点处两个振子的陷俘的双振子束缚态的存在。后一种状态源自于声学声子虚拟云与每个振动子之间的重叠。这是对阿尔法螺旋中振动自陷状态的直接观察。
162.由于多肽的阿尔法螺旋,认知过程、学习和记忆成为可能
163.神经科学领域的不同研究均与更好地理解短期和/或长期的认知机制、学习和记忆有关。
164.我们已经对文献进行了深入的研究,以识别关联多肽和认知机制的文件。
165.表1识别了现有技术中描述的多肽对这些过程的维持、恢复或改进具有积极影响。要么将这些多肽注射到大脑中,要么诱导或提升这些多肽的表达。通过对这些多肽的二级结构进行建模,我们已经建立每个多肽都包括阿尔法螺旋。
166.相反地,报道了对认知机制没有影响的其他多肽,或者更糟糕的是,通过对其二级结构的建模示出了对认知机制有害的其他多肽缺少阿尔法螺旋。
167.例如,虽然barajas
‑
azpeleta等人发现双翅菌肽b和另一种免疫肽(革兰氏阴性细菌结合蛋白样3;gnbp样3)可以调节长期记忆(2018,plos genet 14(10):e1007440),作者指出,attacinb的情况并非如此“令人惊讶的是,尽管attacinb的上调与长期记忆密切相关,attacinb rnai的表达无论在是身体脂肪还是在神经元中都不会干扰记忆。”168.此外,kohler等人(kohler lb,christensen c,rossetti c,fantin m,sandi c,bock e,berezin v(2010)eur j cell biol 89:817
‑
827)示出了dennexina反常的记忆保持。
169.最后,已知β
‑
淀粉样蛋白42(aβ42)在阿尔茨海默病的发病机制中起关键作用,因为aβ42的聚集能力和神经毒性大于aβ40(a.morimoto等,j.biol chem.279,第50期,12月10日发行,第52781
‑
52788页,2004年)。他们也与路易体痴呆有关。id limon等(neuroscience research 63(2009)129
‑
137)证实注射到大鼠海马体中的aβ(25
‑
35)会损害空间记忆。
170.从上文可以看出,每当肽被描述为具有与记忆相关的作用时,它都包含α
‑
螺旋,而缺乏α
‑
螺旋的多肽则被报道为没有作用或起到有害作用。
171.172.173.[0174][0175]
这些发现使得由beltramo等人提出的用于视觉处理模型定义的新方案成为可能(science 363,64
‑
69;2019)。
[0176]
根据视觉处理的标准模型,来自视网膜的所有视觉信息必须首先通过大脑后部的初级视觉皮层(v1)(图4),该初级视觉皮层提取如线条和边缘等简单特征,然后被分发到多个“高阶”视觉区域,这些“高阶”视觉区域提取越来越复杂的特征,如形状、阴影、运动等。
[0177]
由beltramo等发表的研究(science 363,64
‑
69;2019)示出了这些被认为是高阶视觉区域的一个(涉及移动物体的感知)根本不依赖来自v1的信息。相反,这个被称为后鼻腔皮层(por)的区域似乎直接从大脑底部处被称为上丘的进化上古老的感觉处理中心获得视觉数据。
[0178]
在beltramo等的研究(science 363,64
‑
69;2019)中,v1中的活动被光暂时沉默,并且在研究中发现,即使没有来自v1的输入,por神经元也继续对移动刺激做出反应。因此,当皮层中的主要视觉区域沉默时,por中的视觉反应不受影响。如果por对移动物体的反应不是来自v1,那么一定有另一条途径将por与从视网膜进入的视觉信息连接起来。por神经元进一步示出了获得两个解剖学输入的来源—一个来自v1,并且另一个来自上丘,每个来源都通过丘脑(大脑的中央中继站)的不同区域。
[0179]
基于先前的研究,上丘—por系统还可以与恐惧反应、空间注意力和导航,甚至面部识别相关联—所有这些都是por所在的颞叶皮层区域的特性。
[0180]
beltramo等的发现(science 363,64
‑
69;2019)对被称为“盲视”的现象也有影响,在这一现象中,即使不能有意识地感知,因为v1受损而失明的人仍然能够识别物体的位置和跨越障碍(j.l.barbur,l.weiscrantz,ja harlow,pnas,96(1999),第11637
‑
1164页;l.weiskrantz,j.l.barbur,a.sahraie,pnas,92(1995),第6122
‑
6126页)。基于对灵长类动物的研究,盲视被认为是取决于上丘,但beltramo等的结果表明盲视也可能涉及皮层的por样区域。
[0181]
鉴于我们的发现和上述有关视觉处理所涉及的途径的知识,我们得出结论是由beltramo等(science 363,64
‑
69;2019)提出的视觉处理模型实际上涉及如下途径(见图4):
[0182]
‑
途径1:来自视网膜的视觉信息通过初级视觉皮层(v1),该皮层提取线条和边缘等简单特征,然后被分发到多个“高阶”视觉区域,提取越来越复杂的特征,如形状、阴影、运动等;
[0183]
‑
途径2:来自视网膜的视觉信息通过皮层的其他部分,例如后鼻腔皮层(por)。编码的信息穿过神经元到达具有二级α
‑
螺旋结构的多肽。
[0184]
这些信息在阿尔法螺旋级别以量子位编码。信息的传输可以通过量子游走来完成,并且在阿尔法螺旋的构成网络中以100%的效率进行。
[0185]
长时间的量子重现现象允许信息的保存和存储。
[0186]
因此,信息被存档、存储在多肽内。
[0187]
在这些多肽的输出处,这些信息通常被解码为待传送至皮层的调频(脉冲、方波...),以生成第二图像。
[0188]
根据这种视觉处理模型,初级视觉皮层(途径1)提供实时获得的图像(图像1),而途径2请求同一对象的图像2(如果图像2在之前已存储)。通过路径1获得的图像(图像1)与存储在路径2中的图像(图像2)将在轨迹的末端、在额叶皮层的水平进行比较。图像1(实时)与图像2(记忆)的一致性(视觉感知的整合和合成)产生对图像、物体、面部等的识别。在先
前的学习过程中存储的图像2缺失,将导致识别过程的失败。运动识别是可以实现的,因为时间t0通道2已经记住了图像。因此,可以将在t+1实时感知的、来自通道1的图像与通道2存储在t0的图像进行比较。如果通道1被停用,图像2仍然可以在t+1与在t0存档的图像2进行比较,以感知运动。
[0189]
上述结论为预防和治疗与视觉过程相关的病理领域及广义的认知,特别是记忆和学习的认知开辟了实际前景。此外,阿尔法螺旋多肽可以构成在中型生物和半电子系统(例如:生物计算机)中编码,传输和存储量子信息(量子存储器)的新手段。这使得阿尔法螺旋多肽适用于任何有利于使用量子存储器的设备。例如,该设备是生物计算机
……
纳米机器或纳米机器人。该设备部分或全部由阿尔法螺旋多肽制成,因此可用于传感、驱动或计算。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1