用于裂解玉米赤霉烯酮的手段和方法与流程
本发明涉及一种用于增加α/β-水解酶的稳定性的方法。另外,本发明涉及一种通过本发明的方法能够获得的α/β-水解酶。本发明还提供具有降低的总平均亲水性(gravy)值和/或包含特定突变的α/β-水解酶。此外,本发明涉及本发明的α/β-水解酶用于降解玉米赤霉烯酮(zen)的用途。
背景技术:
真菌毒素是丝状真菌产生的次生代谢物。真菌毒素的一个重要代表是由许多镰孢菌真菌产生并在世界各地均可发现的玉米赤霉烯酮(zen)(以前称为f-2毒素)。这些真菌尤其侵染栽培植物,例如各种各样的谷类,其中真菌侵染通常出现在收获之前,这时真菌生长和/或真菌毒素产生可以在贮藏之前或甚至收获之后发生,即贮藏之前或不适当的贮藏条件下发生。联合国粮农组织(fao)估计,在世界范围内25%的农产品被真菌毒素污染,这导致巨大的经济损失。在一项跨越8年的国际研究中,从2004年1月至2011年12月分析了总共19,757个样品,其中72%样品经测试呈至少一种真菌毒素阳性,发现39%样品被污染,和37%样品经测试呈zen阳性(schatzmayr和streit(2013)‘globaloccurrenceofmycotoxinsinthefoodandfeedchain:factsandfigures.’worldmycotoxinjournal6(3):213-222)。在世界的所有地区中,并且在经测试的所有类别的谷物和饲料作物,例如玉米、大豆粉、小麦、麦麸、ddgs(含有可溶物的干酒糟)中,以及在成品动物饲料混合物中,以高至100%的发生率发现了zen。
zen与雌激素受体结合并且可以引起激素紊乱,在经口摄取后被立即吸收,并且被哺乳动物转变为两个立体异构的代谢物,分别为α-玉米赤霉烯醇(α-zel)和/或β-玉米赤霉烯醇(β-zel)。例如,α-zel以及α-玉米赤霉醇(α-zal)和/或玉米赤霉酮(zan)具有比zen更强的雌激素效应。尽管缀合的zen衍生物具有比zen本身低的雌激素活性,然而在消化道中自这些缀合的zen衍生物可以再次释放zen,从而恢复其完全的雌激素活性。
zen具有高达20,000mg/kg体重的口服ld50,在长期暴露的动物或人中可出现亚急性和/或亚慢性的毒性作用,例如致畸、致癌、雌激素和免疫抑制效应。被zen污染的饲料导致哺乳动物中的发育障碍。其中猪,特别是仔猪对zen是极其敏感的。在饲料中超过0.5ppm的zen浓度导致发育障碍,超过1.5ppm的浓度可以导致猪中的超雌激素性。在牛中,12ppmzen浓度可引起自然流产。
由于通过粘膜,特别是通过胃粘膜以及口腔粘膜zen被迅速吸收,因此立即的且定量的灭活是必要的。在口服给药zen后30分钟就已经可以在血液中检测到。由于zen的有害效应,欧盟具有对食品中的zen的法定上限以及对在饲料中的zen上限的推荐(ecno.1881/2006)。
用于减少食品和饲料产品的zen污染的最初策略是限制真菌生长,例如通过遵循“良好的农业规范”。这尤其包括确保种子没有害虫和真菌侵染,或者确保及时从田间移除农业废弃物。此外,还可以通过使用杀真菌剂来减少田间的真菌生长。收获之后,收获的物料应当以低于15%的残留水分并在低温下贮藏,以防止真菌生长。同样地,在进一步加工之前应当去除被真菌侵染污染的物料。尽管有这一系列的预防性措施,即使在拥有最高农业标准的地区(比如北美和中欧),在2004年至2011年,仍然发现有高达37%的测试玉米样品被zen污染(schatzmayr和streit(2013))。
技术实现要素:
为了克服上述问题和缺陷,需要开发另一种α/β-水解酶,该种α/β-水解酶能够解除zen的毒性且适用于用作食品或饲料添加剂或食品或饲料产品。
在以下描述了、在实施例中说明了、在附图中示出了并在权利要求中反映了本发明的方案。
本发明涉及一种用于增加α/β-水解酶的稳定性的方法,所述α/β-水解酶包含对应于seqidno:1的位置145至218的序列,或包含与对应于seqidno:1的位置145至218的序列具有58%或更高的序列同一性的序列(cap-结构域;与seqidno:1的cap-结构域有58%的同一性),所述方法包括:
在以下位置处置换至少一个氨基酸:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,采用这样的氨基酸置换所述氨基酸:所采用的氨基酸具有比被置换的氨基酸更负的亲水指数,其中所述亲水指数由kyte和doolittle亲水指数确定,由此获得具有增加的稳定性的α/β-水解酶。
此外,本发明涉及一种通过本发明的方法能够获得的α/β-水解酶。
本发明还提供一种具有多肽序列的α/β-水解酶,所述多肽序列包含对应于seqidno:1的位置145至218的序列,或包含与对应于seqidno:1的位置145至218的序列具有大于58%的序列同一性的序列,
其中,所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,所述α/β-水解酶具有的总平均亲水性(gravy)值与具有seqidno:1的多肽序列的α/β-水解酶的gravy值相比负至少0.6%。
本发明还涉及一种具有多肽序列的α/β-水解酶,所述多肽序列包含对应于seqidno:1的位置145至218的序列,或包含与对应于seqidno:1的位置145至218的序列具有大于58%的序列同一性的序列,
其中,所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置185至191的位置,或
-对应于seqidno:2的位置184至190的位置,或
-对应于seqidno:3、4或5的位置185至191的位置,或
-对应于seqidno:6的位置201至208的位置,
其中,所述氨基酸置换选自v→a、g→r、g→s、a→p、a→r、a→d、a→h、a→n、a→g、s→d、p→h、m→d、g→e、i→a、i→v、h→n和q→k,和/或
其中采用选自p、r、d、h、g或n的氨基酸置换所述氨基酸,优选地,所采用的氨基酸选自r、d、h、g或n,更优选地,所采用的氨基酸选自r或n。
本发明还涉及一种具有多肽序列的α/β-水解酶,所述多肽序列包含对应于seqidno:6的位置161至235的序列,或包含与对应于seqidno:6的位置161至235的序列具有大于58%的序列同一性的序列,
其中,所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,所述α/β-水解酶具有的gravy值与具有seqidno:6的多肽序列的α/β-水解酶的gravy值相比负至少0.6%。
此外,本发明涉及一种本发明的α/β-水解酶用于降解zen的用途。
进一步地,本发明涉及一种包含本发明的α/β-水解酶的组合物,优选地,所述组合物为食品或饲料添加剂或食品或饲料产品。
另外,本发明涉及本发明的α/β-水解酶或组合物,其用于在治疗或预防疾病中使用。
进一步地,本发明涉及一种包含本发明的α/β-水解酶或组合物的试剂盒。
附图说明
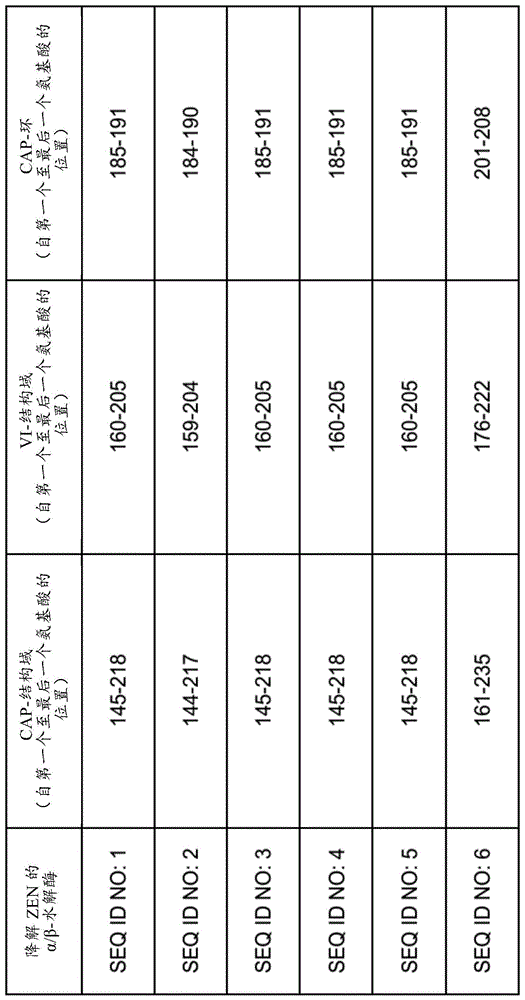
图1:cap-结构域、vi-结构域和cap-环的位置。seqidnos:1-6的cap-结构域、vi-结构域和cap-环的氨基酸位置。
图2a-2g:vi-结构域和/或cap-环中的不同突变及其对gravy值的影响。2a:在seqidno:1的vi-结构域中的修饰对seqidno:1变体的gravy值的影响。2b:在seqidno:1的vi-结构域中的修饰对seqidno:1变体的cap-结构域的gravy值的影响。2c:在seqidno:1的vi-结构域中的修饰对seqidno:1变体的vi-结构域的gravy值的影响。2d:在seqidno:1的cap-环中的修饰对seqidno:1变体的cap-环的gravy值的影响。2e:在seqidno:6的vi-结构域中的修饰对seqidno:6变体的gravy值的影响。2f:在seqidno:6的vi-结构域中的修饰对seqidno:6变体的vi-结构域的gravy值的影响。2g:在seqidno:6的vi-结构域中的修饰对seqidno:6变体的vi-结构域的gravy值的影响。
图3a-3b:相对于多肽seqidno:1或seqidno:6,降解zen的多肽的温度稳定性增加百分比。3a:相对于多肽seqidno:1,降解zen的多肽的温度稳定性增加百分比(t(50%))。3b:相对于多肽seqidno:6,降解zen的多肽的温度稳定性增加百分比(t(50%))。
图4:与在ph7.5(=ph稳定性)下温育后的活性相比,在ph4.0下温育后的降解zen的多肽变体的活性。在ph4.0下温育后的降解zen的多肽变体与在ph7.5下温育后的相同多肽变体相比的残余活性百分比。亲本多肽seqidno:1的残余活性(ph稳定性)为2.5%。
图5:6500qtrap上的选择的反应监测参数,用于分析来自猪饲喂试验的样品。在与6500qtrap质谱仪相连的agilent1290系列uhplc系统上进行来自猪饲喂试验的样品的分析。示出了选择的反应监测参数。产物离子以定量(quantifier)/定性(qualifier)给出。
图6:来自与seqidno:1相比的猪饲喂试验的尿液样品的分析结果。在与6500qtrap质谱仪相连的agilent1290系列uhplc系统上测定每组尿液样品中zen加α-zel的总量(每组的平均值;n=3)。对照组饲喂含zen的食物,但不含降解zen的多肽。seqidno:1组、变体a组和变体b组饲喂与对照组相同的食物,但另外含有2.5u/kg食物、5u/kg食物、10u/kg食物或20u/kg食物的指定的降解zen的多肽。以百分比的形式示出了与seqidno:1相比的尿液中zen加α-zel的量的变化。
图7:来自与seqidno:1相比的猪饲喂试验的粪便样品的分析结果。在与6500qtrap质谱仪相连的agilent1290系列uhplc系统上测定每克冻干的粪便中zen加α-zel的总量(每组的平均值;n=3)。对照组饲喂含zen的食物,但不含降解zen的多肽。seqidno:1组、变体a组和变体b组饲喂与对照组相同的食物,但另外含有2.5u/kg食物、5u/kg食物、10u/kg食物或20u/kg食物的指定的降解zen的多肽。以百分比的形式示出了与seqidno:1相比的粪便中zen加α-zel的量的变化。
图8:6500qtrap上的选择的反应监测参数,用于分析来自仔鸡饲喂试验的样品。在与6500qtrap质谱仪相连的agilent1290系列uhplc系统上进行来自仔鸡饲喂试验的样品的分析。示出了选择的反应监测参数。产物离子以定量/定性给出。
图9:来自与seqidno:1相比的仔鸡饲喂试验的嗉囊样品的分析结果。在与6500qtrap质谱仪相连的agilent1290系列uhplc系统上测定每千克冻干的嗉囊样品中zen的浓度(每组的平均值;n=8)。对照组饲喂含zen的食物,但不含降解zen的多肽。其它组饲喂与对照组相同的食物,但另外含有指定量的酶活性的降解zen的多肽变体b。以百分比的形式示出了与对照组相比的嗉囊中zen的浓度变化。
具体实施方式
惊讶地发现,在特定区域,即vi-结构域和cap-环中包含本文所述的突变的α/β-水解酶表现出更高的温度稳定性和/或ph稳定性。不受理论的束缚,认为vi-结构域和cap-环在例如用于底物进入酶的活性位点的酶活性中起重要作用。酶的这部分的高度柔性可对活性产生正面影响,然而,这种柔性同样会对稳定性产生负面影响。
本发明的发明人将seqidno:1的cap-结构域识别为位置145至218的氨基酸,将seqidno:2的cap-结构域识别为位置144-217,将seqidno:3、4和5的cap-结构域识别为位置145-218,和将seqidno:6的cap-结构域识别为位置161-235。此外,本申请的发明人将seqidno:1的vi-结构域识别为氨基酸位置160至205,将seqidno:2的vi-结构域识别为氨基酸位置159-204,将seqidno:3、4和5的vi-结构域识别为氨基酸位置160-205,和将seqidno:6的vi-结构域识别为氨基酸位置176-222。特别地,通过phenix全精(ensemblerefinement)(https://www.phenix-online.org/;burnley和gros(2012)‘phenix.ensemble_refinement:ateststudyofapoandholobace1‘computationalcrystallographynewsletter,volume4,pp.51–58)将例如seqidno:1或6的变体的动力学模拟与x射线衍射数据的组合通过生成65个结构反映了柔性环。由氨基酸位置185至191定义的seqidno:1的区域(在本文中定义为cap-环)是该柔性环的一部分(并且由如本文所述的seqidno:2-6中的等效位置定义的区域也是)。
引入cap-结构域,特别是引入本文定义的vi-结构域,或更具体地引入本文定义的cap-环中的如本文所述的突变提供了足够的温度稳定性而不丧失活性性质和/或ph稳定性,使得可以将这种酶用在升高温度的工艺过程中。
这一点特别重要,因为在食品和饲料工业中,常将诸如制粒的热处理用于生产具有降低的微生物载量的卫生产品中。
饲料的制粒是增强流动性、减少粉尘形成和降低微生物(特别是沙门氏菌)载量的特别普遍的标准化方法。在制粒过程中,产品通常通过热蒸汽润湿、加热并随后在压力下通过矩阵(matrix)压制。酶或多肽的这种热处理经常导致其酶活性降低和/或其不可逆变性。
此外,当将酶用于食品或饲料中时,其通常经受被动物胃肠道内的条件灭活。特别是低ph环境可导致酶或多肽的酶活性暂时或永久地降低或甚至消除。
然而,降解zen的酶通常具有低的温度稳定性和/或ph稳定性,从而不能被混合到饲料或食品中。因此,将多肽或酶用作制粒食品或饲料的添加剂成为了相当大的技术挑战。
本文所述的α/β-水解酶具有增加的稳定性,尤其是在温度和/或ph稳定性方面,因此非常适合用于食品和饲料的生产过程。
因此,本发明涉及一种用于增加α/β-水解酶的稳定性的方法,所述α/β-水解酶包含对应于seqidno:1的位置145至218的序列,或包含与对应于seqidno:1的位置145至218的序列具有58%或更高的序列同一性的序列(cap-结构域;与seqidno:1的cap-结构域有58%的同一性),所述方法包括:
在以下位置处置换至少一个氨基酸:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,采用这样的氨基酸置换所述氨基酸:所采用的氨基酸具有比被置换的氨基酸更负的亲水指数,
其中所述亲水指数由kyte和doolittle亲水指数确定,
由此获得具有增加的稳定性的α/β-水解酶,优选地,与置换所述氨基酸之前的α/β-水解酶相比,所述α/β-水解酶具有增加的稳定性,和/或与不含所述氨基酸置换的α/β-水解酶相比,所述α/β-水解酶具有增加的稳定性。
本发明还涉及一种用于增加α/β-水解酶的稳定性的方法,所述α/β-水解酶包含对应于seqidno:6的位置161至235的序列,或包含与对应于seqidno:6的位置161至235的序列具有58%或更高的序列同一性的序列(cap-结构域;与seqidno:6的cap-结构域有58%的同一性),所述方法包括:
在以下位置处置换至少一个氨基酸:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,采用这样的氨基酸置换所述氨基酸:所采用的氨基酸具有比被置换的氨基酸更负的亲水指数,
其中所述亲水指数由kyte和doolittle亲水指数确定,
由此获得具有增加的稳定性的α/β-水解酶,优选地,与置换所述氨基酸之前的α/β-水解酶相比,所述α/β-水解酶具有增加的稳定性,和/或与不含所述氨基酸置换的α/β-水解酶相比,所述α/β-水解酶具有增加的稳定性。
如本文所用的增加的稳定性可以意指,与包含对应于seqidno:1(以及seqidno:3、4、5)的位置145至218的序列的α/β-水解酶相比,本发明的α/β-水解酶具有更高的稳定性。可选地或另外地,如本文所用的增加的稳定性意指,与包含对应于seqidno:2的位置144至217的序列的α/β-水解酶相比,本发明的α/β-水解酶具有更高的稳定性。可选地或另外地,α/β-水解酶包含对应于seqidno:6的位置161至235的序列。
如本文所用的增加的稳定性还可以意指,与seqidno:1的α/β-水解酶相比,本发明的α/β-水解酶具有更高的稳定性。可选地或另外地,如本文所用的增加的稳定性还可以意指,与seqidno:2的α/β-水解酶相比,本发明的α/β-水解酶具有更高的稳定性。可选地或另外地,如本文所用的增加的稳定性还可以意指,与seqidno:6的α/β-水解酶相比,本发明的α/β-水解酶具有更高的稳定性。
这包括,与不含本文所述的置换的α/β-水解酶相比,所述例如通过本发明的方法获得的具有增加的稳定性的α/β-水解酶或本发明的α/β-水解酶具有增加的稳定性。类似的,与置换本文公开的氨基酸之前的α/β-水解酶相比,所述例如通过本发明的方法获得的具有增加的稳定性的α/β-水解酶或本发明的α/β-水解酶具有增加的稳定性。
本领域技术人员已知各种α/β-水解酶,其特别描述于lenfant等,(2013)‘esther,thedatabaseoftheα/β-hydrolasefoldsuperfamilyofproteins:toolstoexplorediversityoffunctions’nucleicacidsresearch,volume41,issued1,d423–d429和mindrebo等,(2016)‘unveilingthefunctionaldiversityofthealpha-betahydrolasefoldinplants’curropinstructbiol.233–246中。简言之,所有的α/β-水解酶都具有特定的折叠特征,称为α/β-折叠(alpha/beta-折叠)。该种α/β-水解酶折叠是许多系统起源和催化功能差别很大的水解酶所共有的。每种酶的核心是α/β-结构(而非桶),其含有由α-螺旋(aa-af)连接的8条β-链(b1-b8)(ollis等,(1992)‘thealpha/betahydrolasefold‘proteineng.5(3):197-211)。因此,本文所述的α/β-水解酶可包含α/β-折叠。优选地,本文所述的α/β-水解酶包含由排列成中央β-折叠的8条β-链(b1-b8)组成的α/β-水解酶核心结构域,并另外包含6个交叉α-螺旋(aa-af)。
在多数家族成员中,β-链为平行取向,但是一些成员具有第一链倒位,从而导致了反式平行取向。处于折叠的酶的原型具有催化三联体,该三联体由位于第五β-链和之后的α-螺旋(亲核体肘)之间的γ-转角顶部的亲核残基、一个酸性氨基酸残基(谷氨酸或天冬氨酸)和一个组氨酸残基组成。一些其它成员缺少一个或所有催化残基。因此,一些成员是无活性的;一些成员参与表面识别。优选地,本文所述的α/β-水解酶包含所述催化三联体。
α/β-水解酶不同类别的成员及它们的结构特征特别描述于kourist等,(2010)‘thealpha/beta-hydrolasefold3dmdatabase(abhdb)asatoolforproteinengineering.’chembiochem.11(12):1635-43)中。
作为酶,通常将α/β-水解酶描述为负责酯键和肽键的水解。然而,α/β-水解酶还参与碳-碳键的断裂、脱羧反应和杂芳环的无辅助因子(cofactor-independent)的双氧化。因此,α/β-水解酶可包括该超家族中的催化成员(酶)。非限制性实例是水解酶(乙酰胆碱酯酶、羧酸酯酶、双烯内酯水解酶、脂肪酶、角质酶、硫酯酶、丝氨酸羧基肽酶、脯氨酸亚氨基肽酶,脯氨酸寡肽酶、环氧化物水解酶)以及需要hcn、h2o2或o2而非h2o的活化用于反应机制的酶(卤代烷脱卤素酶、卤代过氧化物酶、羟基腈裂解酶)。非催化成员可包括神经连接蛋白、谷蛋白(glutactin)、神经趋化因子、甲状腺球蛋白的c-末端结构域、卵黄蛋白、ccg1-相互作用因子-b和二肽氨基肽酶vi。
esther数据库收集并注释与该超家族的基因和蛋白质序列相关的公开信息。因此,本领域技术人员还可以从esther(http://bioweb.supagro.inra.fr/esther/general?what=index)(α/β-水解酶-折叠蛋白超家族的数据库)中获得α/β-水解酶。
本领域技术人员还可以确定α/β-水解酶是否包含cap-结构域。实现此目的的一种方法在实施例中描述或如下所述:
1、在在线酶数据库中检索α/β-水解酶或通过将给定序列与seqidno:1-6比较。
2、优选地通过采用实施例2中所述的程序确定α/β-水解酶是否包含cap-结构域、vi-结构域或cap-环。
3、优选地通过采用实施例4中所述的程序确定包含cap-结构域、vi-结构域或cap-环的α/β-水解酶能否水解zen。
如本文所述的α/β-水解酶包含对应于seqidno:1的位置145至218的序列,或包含与对应于seqidno:1的位置145至218的序列具有58%或更高的序列同一性的序列(cap-结构域)。因此,包含该序列的任何α/β-水解酶均涵盖在术语α/β-水解酶内。这一序列对应于seqidno:1的α/β-水解酶的cap-结构域。
另外地或可选地,如本文所述的α/β-水解酶还可包含对应于seqidno:2的位置144至217的序列,或包含与对应于seqidno:2的位置144至217的序列具有58%或更高的序列同一性的序列(cap-结构域)。因此,包含该序列的任何α/β-水解酶均涵盖在术语α/β-水解酶内。这一序列对应于seqidno:2的α/β-水解酶的cap-结构域。
另外地或可选地,如本文所述的α/β-水解酶还可包含对应于seqidno:3、4或5的位置145至218的序列,或包含与对应于3、4或5的位置145至218的序列具有58%或更高的序列同一性的序列(cap-结构域)。因此,包含该序列的任何α/β-水解酶均涵盖在术语α/β-水解酶内。这一序列对应于3、4或5的α/β-水解酶的cap-结构域。
另外地或可选地,如本文所述的α/β-水解酶还可包含对应于seqidno:6的位置161至235的序列,或包含与对应于seqidno:6的位置161至235的序列具有58%或更高的序列同一性的序列(cap-结构域)。因此,包含该序列的任何α/β-水解酶均涵盖在术语α/β-水解酶内。这一序列对应于seqidno:6的α/β-水解酶的cap-结构域。
例如,所述α/β-水解酶可包含与seqidno:1的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。另外地或可选地,所述α/β-水解酶可包含与seqidno:2的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。另外地或可选地,所述α/β-水解酶可包含与seqidno:3、4和/或5的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。另外地或可选地,所述α/β-水解酶可包含与seqidno:6的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。
当在本文使用时,术语“多肽”意指肽、蛋白质或多肽,它们可以互换使用并涵盖给定长度的氨基酸链,其中氨基酸残基通过共价肽键连接。本发明还涵盖除本领域技术人员已知的标准遗传密码的20种蛋白原性氨基酸之外的氨基酸,比如硒代半胱氨酸。此类多肽包括seqidno:1-6中的任何一种。
术语多肽还指,且不排除多肽的修饰。修饰包括糖基化、乙酰化、酰基化、磷酸化、adp-核糖基化、酰胺化、黄素的共价连接,血红素部分的共价连接、核苷酸或核苷酸衍生物的共价连接、脂质或脂质衍生物的共价连接、磷脂酰肌醇的共价连接、交联、环化、二硫键形成、去甲基化、共价交联的形成、半胱氨酸的形成、焦谷氨酸盐的形成、制剂化(formulation)、γ-羧基化、糖基化、gpi锚形成、羟基化、碘化、甲基化、豆蔻酰化、氧化、聚乙二醇化、蛋白酶解加工、磷酸化、异戊烯化、外消旋化、硒酰化(selenoylation)、硫酸盐化、转移-rna介导的向蛋白质添加氨基酸比如精氨酸化和泛素化;参见,例如proteins-structureandmolecularproperties,2nded.,t.e.creighton,w.h.freemanandcompany,newyork(1993);post-translationalcovalentmodificationofproteins,b.c.johnson,ed.,academicpress,newyork(1983),pgs.1-12;seifter,meth.enzymol.182(1990);626-646,rattan,ann.nyacad.sci.663(1992);48-62。
根据本发明,在两个或更多个多肽序列(比如seqidno:1-6)的情况下术语“同一的”或“同一性百分比”指当在比较窗口上或在测量的指定区域上采用本领域已知的序列比较算法或通过手动比对和目视检查进行最大对应性的比较和比对时,两个或更多个序列或子序列相同或具有相同的特定百分比的核苷酸(例如至少85%、90%、95%、96%、97%、98%或99%的同一性)。将具有例如80%至95%或更高的序列同一性的序列认为基本上是同一的。这一定义也可应用于测试序列的互补序列。本领域技术人员将知道如何使用例如基于如本领域已知的clustalw计算机程序(thompsonnucl.acidsres.2(1994),4673-4680)或fastdb(brutlagcomp.app.biosci.6(1990),237-245)的那些来测定序列间/中的同一性百分比。
本领域技术人员还可用的是blast和blast2.6算法(altschulnucl.acidsres.25(1977),3389-3402)。用于氨基酸序列的blastp程序使用默认的字长(w)6,期望阈值10和两条链都进行比较。此外,还可使用blosum62评分矩阵(henikoffproc.natl.acad.sci.,usa,89,(1989),10915;henikoff和henikoff(1992)‘aminoacidsubstitutionmatricesfromproteinblocks.‘procnatlacadsciusa.1992nov15;89(22):10915-9)。
例如,可以将代表基本局部比对搜索工具的blast2.6(altschul,nucl.acidsres.25(1997),3389-3402;altschul,j.mol.evol.36(1993),290-300;altschul,j.mol.biol.215(1990),403-410)用于搜索局部序列比对。
如本文所使用的“cap-结构域”涉及α/β-水解酶的cap-结构域,其例如描述于kourist等,(2010)‘thealpha/beta-hydrolasefold3dmdatabase(abhdb)asatoolforproteinengineering.’chembiochem.11(12):1635-43的图1中或描述于carr和ollis(2009)‘α/βhydrolasefold:anupdate.’protein&peptideletters,2009,16(10):1137-1148中。还设想cap-结构域可位于β-折叠和α-螺旋之间例如α/β-水解酶的b6与ad之间的偏移(excursion)内,例如ollis等,(1992)‘thealpha/betahydrolasefold‘proteineng.5(3):197-211中所述。例如,cap-结构域可以起始于α/β-水解酶核心结构域的b6β-链的c末端后不远处,并且可以持续至α/β-水解酶核心结构域的adα-螺旋的n末端起始处。然而,该cap-结构域还可包含β-折叠或其他蛋白质结构。
本发明的方法需要在α/β-水解酶的以下位置处置换至少一个氨基酸:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置。这些位置均位于vi-结构域中。
如本文所用的‘vi-结构域’是cap-结构域的一部分。因此,cap-结构域包含vi-结构域。该vi-结构域可以起始于存在于cap-结构域中的qxagp基序(seqidno:7)之后的第一个氨基酸并且可以持续至eydpe基序(seqidno:8)之前的最后一个氨基酸,其中eydpe基序不是vi-结构域的一部分。这些基序在本文表2中描述的序列中加下划线。
例如,所述vi-结构域可包含对应于seqidno:1的位置160至205的序列,或包含与对应于seqidno:1的位置160至205的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。另外地或可选地,所述vi-结构域可包含对应于seqidno:2的位置159至204的序列,或包含与对应于seqidno:2的位置159至204的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。另外地或可选地,所述vi-结构域可包含对应于seqidno:3、4和/或5的位置160至205的序列,或包含与对应于seqidno:3、4和/或5的位置160至205的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。另外地或可选地,所述vi-结构域可包含对应于seqidno:6的位置176至222的序列,或包含与对应于seqidno:6的位置176至222的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。
当根据本发明使用时,术语“位置”意指氨基酸在本文描述的氨基酸序列中的位置。如本文所用的术语“对应”还包括位置不仅由在前的氨基酸的数量决定。根据本发明可被置换的给定氨基酸的位置可以由于缺失或添加的氨基酸而改变,或可被置换的给定氨基酸的位置可以由于(突变体或野生型)α/β-水解酶中的其它地方的氨基酸缺失或添加而改变。
因此,就根据本发明的“对应位置”,优选地理解为氨基酸在指定编号上可以不同,但是仍可具有相似的相邻氨基酸。术语“对应位置”还包括可被交换、缺失或添加的所述氨基酸。具体而言,当将例如seqidno:1-6中的任一个、优选seqidno:1的参考序列(目标序列(subjectsequence))与目的氨基酸序列(查询序列(querysequence))比对时,本领域技术人员可以例如,在查找本文所述的氨基酸位置(即对应于seqidno:1中示出的氨基酸序列的位置185和/或188的位置)时检查目的序列来寻找seqidno:1的序列(或编码该蛋白质的相应氨基酸序列)。
在本发明的方法中,采用这样的氨基酸置换所述氨基酸:所采用的氨基酸具有比被置换的氨基酸更负的亲水指数,其中所述亲水指数由kyte和doolittle亲水指数确定。
如本文所述,“氨基酸置换”意指相对于所识别的seqidno的相应位置(例如seqidno:1-6的本文指定的位置中的任意一个)的氨基酸的替换。例如,在一个实施方案中,所述替换是相对于对应于seqidno:1的位置160至205的位置的氨基酸的氨基酸置换。
“亲水指数”在本文中也称为“亲水值”,是代表氨基酸侧链的疏水性或亲水性的数字。特别地,对于亲水指数,已经对每个氨基酸赋予反映其相对亲水性的值。因此,氨基酸的亲水性可以由亲水指数来确定。jackkyte和russellf.doolittle(kyte和doolittle(1983)"asimplemethodfordisplayingthehydropathiccharacterofaprotein".j.mol.biol.157(1):105–32)提出了氨基酸的该亲水指数。具有最不负的亲水指数的氨基酸为异亮氨酸(4.5)和缬氨酸(4.2)。根据kyte和doolittle,具有最负的亲水指数的氨基酸为精氨酸(-4.5)和赖氨酸(-3.9)。认为亲水指数在蛋白质结构中是重要的。具有较不负的亲水指数的氨基酸倾向于内部(就蛋白质的三维形状而言)而具有更负的亲水指数的氨基酸更常见于蛋白质表面。已将kyte和doolittle的亲水指数总结于本文的表1:
表1:kyte和doolittle的亲水指数
进一步设想至少一个、两个、三个、四个、五个、六个、七个、八个、九个、十个、十一个、十二个或更多个氨基酸被置换。
所述方法还设想在以下位置处置换至少一个氨基酸:
-对应于seqidno:1的位置185至191的位置,和/或
-对应于seqidno:2的位置184至190的位置,和/或
-对应于seqidno:3、4或5的位置185至191的位置,和/或
-对应于seqidno:6的位置201至208的位置。
所有这些位置均位于cap-环内。
在上下文中,应注意cap-结构域和vi-结构域可进一步包含环(序列/结构域)。该‘环’在本文中也称为‘cap-环’,其可以起始于存在于vi-结构域中的g(f/y)xxaa(seqidno:9)基序之后的第一个氨基酸之后并且可以持续至arxf基序(seqidno:10)(或对于seqidno:6而言的qlfp基序(seqidno:11))之前的最后一个氨基酸,其中arxf基序(seqidno:10)(或对于seqidno:6而言的qlfp基序(seqidno:11))不是cap-环的一部分。这些基序在下文表2中描述的序列中加下划线。
例如,所述cap-环可包含对应于seqidno:1的位置185至191的序列,或包含与对应于seqidno:1的位置185至191的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。另外地或可选地,所述cap-环可包含对应于seqidno:2的位置184至190的序列,或包含与对应于seqidno:2的位置184至190的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。另外地或可选地,所述cap-环可包含对应于seqidno:3、4和/或5的位置185至191的序列,或包含与对应于seqidno:3、4和/或5的位置185至191的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。另外地或可选地,所述cap-环可包含对应于seqidno:6的位置201至208的序列,或包含与对应于seqidno:6的位置201至208的序列具有至少58%、59%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%的同一性的序列。
因此,如本文所述的α/β-水解酶可包含如本文所述的vi-结构域和/或cap-环。
还预期采用选自r、k、n、q、d、e、h、p、y、w、s、t、g、a、m、c、f、l或v的氨基酸置换所述氨基酸。
进一步设想采用选自r、k、n、q、d、e、h、p、y、w、s、t或g的氨基酸置换所述氨基酸。
还预期采用选自r、k、n、q、d、e、h或p的氨基酸置换所述氨基酸。
进一步设想采用选自r、k、d、q、d、n、e、p、g、t、s或h的氨基酸置换所述氨基酸。
还预期采用选自s、p、r、d、h、g或n的氨基酸置换所述氨基酸。还可采用选自r、d、h、g或n的氨基酸置换所述氨基酸。
还预期采用选自p、s、r或h的氨基酸置换所述氨基酸。还可采用选自r或n的氨基酸置换所述氨基酸。
进一步设想,所述氨基酸置换选自以下的一个或多个:v→a、g→r、g→s、a→p、a→r、a→d、a→h、a→n、a→g、s→d、p→h、m→d、g→e、i→a、i→v、h→n、q→k、f→y和/或v→c。
所述氨基酸置换还可选自以下的一个或多个:v160a、g185r、g185s、a186p、a186r、a188d、a188h、a188n、a188g、a188r、s189d、p190h、m191d、g199e、i200a、i200v、h203n、q205k、f183y和/或v197c。
还预期所述氨基酸置换选自以下的一个或多个:v→a、g→r、g→s、a→p、a→r、a→d、a→h、a→n、a→g、s→d、p→h、m→d、g→e、i→a、i→v、h→n和/或q→k。所述氨基酸置换还可选自以下的一个或多个:v160a、g185r、g185s、a186p、a186r、a188d、a188h、a188n、a188g、a188r、s189d、p190h、m191d、g199e、i200a、i200v、h203n和/或q205k。
所述氨基酸置换还可选自g185r、a186r、a188r、a188d、a188h、a188n和/或m191d。
进一步设想,采用选自r、d、h、g、n或p的氨基酸置换所述氨基酸。
还预期,本发明的方法包括在以下位置处置换至少一个氨基酸:
-对应于seqidno:1的位置185至191的位置,和/或
-对应于seqidno:2的位置184至190的位置,和/或
-对应于seqidno:3、4或5的位置185至191的位置,和/或
-对应于seqidno:6的位置201至208的位置。
并且其中采用选自r、d、h、g、n或p的氨基酸置换所述氨基酸。
本发明涉及一种用于增加α/β-水解酶的稳定性的方法。所述增加的稳定性可以为gravy值的降低,ph稳定性的增加和/或温度稳定性的增加。
如本文所用,蛋白质的“gravy值”是其相对疏水性或亲水性的量度。将两个量度组合于亲水度标度或亲水指数中。根据kyte和doolittle(kytej,doolittlerf(may1983)."asimplemethodfordisplayingthehydropathiccharacterofaprotein".j.mol.biol.157(1):105–32),通过每个残基的亲水值(亲水指数,参见上文表1)相加并除以序列中残基的数量,计算所述gravy值。因此,gravy值可以通过全部氨基酸的亲水值(指数)之和除以序列中氨基酸残基的数量(根据kyte和doolittle的计算)来计算。如本文所用,术语“温度稳定性”是指在临时暴露于升高的温度后,酶维持其催化活性的性质。通过测量并比较酶或多肽溶液在10分钟热处理之前与之后或在相同的限定条件下未热处理的酶活性来确定温度稳定性。
特别地,可以按照以下方式来测量温度稳定性。采用样品缓冲液(ph7.5的teorellstenhagen缓冲液(stenhagen&teorell.(1938)nature141,415),含有0.1mg/ml牛血清白蛋白)将多肽稀释至0.001526923u/ml的浓度并放置于冰上待用。将40份50μl稀释的多肽溶液的等分试样转移至4组12-连管(例如购自starlab)的管中,其中每组连管的第一个和最后一个管空置。所述连管采用12-连帽(例如购自starlab)密封。作为阳性对照,将4份50μl稀释酶溶液的等分试样转移至4个pcr管中。将全部pcr管和连管均在冰上放置直至开始温度温育步骤。作为阴性对照,将4份50μl样品缓冲液的等分试样转移至4个pcr管中。这些管在25℃下储存。
在选定温度+/-10℃下,将4组12-连管在具有梯度功能(例如eppendorfmastercycler梯度)的预热pcr循环仪中温育。pcr循环仪自动计算沿pcr循环仪加热块的温度梯度(选定温度的+/-10℃)。将含有阳性对照的pcr管在冰上温育,将含有阴性对照的pcr管在25℃下温育。在0、5、10和20分钟后,将1组pcr连管和1个阴性对照管转移以放置在冰上直至温育结束,所述温育结束即温育开始后20分钟。在全部温育步骤完成并将所有连管和管均放置在冰上之后,开始zen降解分析。
制备zen降解分析缓冲液(teorellstenhagen缓冲液,ph7.5,含有0.1mg/ml牛血清白蛋白和5.3ppmzen),并将660μl分析缓冲液的等分试样转移至48个反应管中。将所述管密封并保存在25℃下直至开始zen降解分析。对于降解分析,将来自pcr连管的40个温度处理样品的每一个的40μl、4个阴性对照的每一个的40μl和4个阳性对照的每一个的40μl加入含有660μl分析缓冲液的管中,由此实现分析反应中最终zen浓度为5ppm。此外,由此还实现了多肽的终浓度以在三个小时内有效降解zen(即90%-100%的zen降解)。
通过将温度处理样品、阳性或阴性对照加入分析缓冲液中,开始降解分析。将zen降解反应在37℃的预热水浴中温育。在降解反应开始后立即通过涡旋混合约2秒,并将100μl的0h样品转移到新的反应管中。在0.5、1.0、2.0和3.0小时后,从zen降解分析反应中抽取其它的样品。一旦将样品自降解反应中抽取出,就通过在99℃下温育10分钟使该样品中的酶热灭活。随后,将管离心(2分钟,25℃,14674xg)并将90μl上清液转移至具有插入件的hplc小瓶中。将这些hplc小瓶储存在4℃下直至按照实施例4所述进行hplc-dad测量。
采用通过zen降解样品的hplc-dad分析确定的zen浓度的线性降低,以单位/升(u/l)计算酶活性。将一个单位定义为在所述条件下在1分钟内降解1μmolzen的酶活性的量。在不同温度下温育0、5、10和20分钟后的残余活性计算如下:温度处理样品中的酶活性除以阳性对照的酶活性的平均值,再乘以100。
将温度稳定性(t(50%))定义为这样的温度:在该温度下温育10分钟后的多肽具有相比于阳性对照的50%残余活性。以下实例用于说明:亲本酶在冰上温育10分钟后具有50u/ml的酶活性,在59.3℃下温育10分钟后具有25u/ml的活性,则t(50%)值为59.3℃。如果酶变体具有61.0℃的t(50%)值,则温度稳定性(t(50%))与亲本酶相比的相对增加为2.9%。这是将两个t(50%)值之间的差值1.7℃除以亲本酶的t(50%)值59.3℃再乘以100得到的。
因此,如本文所用的温度稳定性是一种当临时暴露于选自20℃至85℃的温度时酶活性对失活的抗性的量度。可以将温育10分钟后的热处理的酶的残余活性为50%时的温度与阳性对照进行比较。在本文中将多肽变体相对于其亲本多肽的t(50%)的增加定义为增加的温度稳定性,并且这种增加可以相对地表示为百分比值或绝对地以摄氏度表示。
如本文所用的术语“ph稳定性”指在特定ph下临时温育后,多肽维持其催化活性的性质,因此“ph稳定性”通过在特定ph下临时温育后的多肽的残余活性来反映。通过将在不同ph缓冲液中温育60分钟后的多肽溶液降解zen的酶活性与在ph7.5的缓冲液中温育60分钟后的相同多肽溶液的酶活性进行比较来确定在特定ph下温育后的残余活性。所述ph稳定性是酶对临时暴露于特定ph环境的抗性的量度。将ph稳定性的增加定义为与在ph4.0下温育后的亲本酶变体的残余活性相比,在ph4.0(=ph处理)下温育后的多肽变体的残余活性的增加。
所述ph稳定性可以按照如下测量。将降解zen的多肽在不同ph值的缓冲溶液中温育1小时。将含有多肽变体的等分试样转移至8个含有8种不同ph值的温育缓冲液的样品管中。所述温育缓冲液为不含奶且半浓缩的fedstatesimulatedgastricfluid中间缓冲液(jantratid等,(2008)’dissolutionmediasimulatingconditionsintheproximalhumangastrointestinaltract:anupdate.’pharmres.2008jul;25(7):1663-76)。将8个样品管中的温育缓冲液的ph值分别设定为3.5、4.0、4.2、4.4、4.6、4.8、5.0和6.0。还将多肽变体的1份等分试样转移至一个含有样品缓冲液(teorellstenhagen缓冲液,ph7.5,含有0.1mg/ml牛血清白蛋白)的管中作为阳性对照。作为阴性对照,将100μl样品缓冲液在预热的水浴中于37℃下温育1小时。温育后,在类似于本文其它地方所述的或实施例(例如实施例4)中所述的分析缓冲溶液中测定样品降解zen的能力。zen降解分析缓冲液的加入确保所有样品中ph值恒定为ph7.5。在整个zen降解分析反应中取样并按照例如实施例4中所述采用hplc-dad测量来分析zen、水解的玉米赤霉烯酮(hzen)和脱羧水解的玉米赤霉烯酮(dhzen)的浓度。例如按照实施例4所述计算活性。
将ph稳定性的增加定义为与未突变的亲本酶溶液在相同处理后的残余活性相比,在ph4.0下温育后的多肽溶液的残余活性的增加。通过ph-处理的多肽溶液的活性与相同多肽变体溶液在ph7.5下温育后的活性的比较来定义所述残余活性。按照如下计算所述残余活性:ph-处理的样品的酶活性除以在ph7.5下温育的对照的酶活性,再乘以100。以下实例用于说明:如果多肽样品在ph4.0下温育后的酶活性是0.5u/l且该多肽在ph7.5下温育后的酶活性是2.7u/l,则残余活性是18.5%。如果将亲本多肽seqidno:1在ph4.0下温育后的残余活性测量为2.5%,则与亲本多肽相比,该多肽变体的ph稳定性增加为7.4倍。
本发明还涉及一种通过本文所述的方法可获得的α/β-水解酶。
本发明还涉及一种具有多肽序列的α/β-水解酶,所述多肽序列包含对应于seqidno:1的位置145至218的序列,或包含与对应于seqidno:1的位置145至218的序列具有大于58%的序列同一性的序列,
其中,所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,所述α/β-水解酶具有的gravy值与具有seqidno:1的多肽序列的α/β-水解酶的gravy值相比负至少0.6%。
本发明还涉及一种具有多肽序列的α/β-水解酶,所述多肽序列包含对应于seqidno:6的位置161至235的序列,或包含与对应于seqidno:6的位置161至235的序列具有大于58%的序列同一性的序列,
其中,所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,所述α/β-水解酶具有的gravy值与具有seqidno:6的多肽序列的α/β-水解酶的gravy值相比负至少0.6%。
还预期,所述α/β-水解酶具有的gravy值与具有seqidno:1的多肽序列的α/β-水解酶的gravy值相比低至少3.0%、4.2%、4.8%、6.0%、6.6%、7.8%、10.2%、12.0%或更高。
还预期,所述α/β-水解酶具有的gravy值与具有seqidno:6的多肽序列的α/β-水解酶的gravy值相比低至少1.0%、2.0%、2.5%、2.6%、3.0%、4.0%、5.0%、6.0%、6.8%或更高。
本发明还涉及一种具有多肽序列的α/β-水解酶,所述多肽序列包含对应于seqidno:1的位置145至218的序列,或包含与对应于seqidno:1的位置145至218的序列具有大于58%的序列同一性的序列,
其中,所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置185至191的位置,或
-对应于seqidno:2的位置184至190的位置,或
-对应于seqidno:3、4或5的位置185至191的位置,或
-对应于seqidno:6的位置201至208的位置,
其中,所述氨基酸置换选自v→a、g→r、g→s、a→p、a→r、a→d、a→h、a→n、a→g、s→d、p→h、m→d、g→e、i→a、i→v、h→n和q→k,和/或
其中,采用选自p、r、d、h、g或n的氨基酸置换所述氨基酸,优选地,所采用的氨基酸选自r、d、h、g或n,更优选地,所采用的氨基酸选自r或n。此类α/β-水解酶可具有比不含此置换的或在引入这些置换之前的相同α/β-水解酶更高的稳定性。例如,此类α/β-水解酶可具有与seqidno:1的α/β-水解酶相比更高的稳定性。
本发明还涉及一种具有多肽序列的α/β-水解酶,所述多肽序列包含对应于seqidno:6的位置161至235的序列,或包含与对应于seqidno:6的位置161至235的序列具有大于58%的序列同一性的序列,
其中,所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置185至191的位置,或
-对应于seqidno:2的位置184至190的位置,或
-对应于seqidno:3、4或5的位置185至191的位置,或
-对应于seqidno:6的位置201至208的位置,
其中,所述氨基酸置换选自f→y或v→c。此类α/β-水解酶可具有比不含此置换的或在引入这些置换之前的相同α/β-水解酶更高的稳定性。例如,此类α/β-水解酶可具有与seqidno:6的α/β-水解酶相比更高的稳定性。
设想如本文所述的α/β-水解酶包含以下氨基酸置换:
-g→r和a→n,优选地g185r和a188n;
-g→s和a→r,优选地g185s和a188r;
-g→r、a→r、a→h、s→d、p→h和m→d,优选地g185r、a186r、a188h、s189d、p190h和m191d;
-v→a、g→r、a→n、g→e、i→a、h→n和q→k,优选地v160a、g185r、a188n、g199e、i200a、h203n和q205k;
-v→a、g→s、a→r、g→e、i→a、h→n和q→k,优选地v160a、g185s、a188r、g199e、i200a、h203n和q205k;
-v→a、g→e、i→a、h→n和q→k,优选地v160a、g199e、i200a、h203n和q205k;
-v→a、g→r、a→r、a→h、g→e、i→v、h→n和q→k,优选地v160a、g185r、a186r、a188h、g199e、i200v、h203n和q205k;和/或
f→y和v→c,优选地f183y和v197c。
本发明还涉及编码本文所述的α/β-水解酶的核酸分子。可将所述核酸分子引入或插入表达载体。术语“表达载体”是指能够在体内或体外表达基因的核酸分子构建体。特别地,其涵盖dna构建体,此类dna构建体适用于将编码多肽的核苷酸序列转移至宿主细胞,从而将该核苷酸序列整合到基因组中或者游离地存在于染色体外的空间中,并且在该细胞内表达编码所述多肽的核苷酸序列和任选地自该细胞中提取出所述多肽。
本文所述的表达载体可在宿主细胞中表达。术语“宿主细胞”是指所有这样的细胞:其包含待表达的核苷酸序列,或包含表达载体,并且能够产生根据本发明的酶或多肽。特别地,该术语是指原核细胞和/或真核细胞,优选地为巴斯德毕赤酵母(pichiapastoris)、大肠杆菌(escherichiacoli)、枯草芽孢杆菌(bacillussubtilis)、链霉菌属(streptomyces)、汉逊酵母属(hansenula)、木霉属(trichoderma)、乳杆菌属(lactobacillus)、曲霉属(aspergillus)、植物细胞和/或芽孢杆菌属(bacillus)、木霉属(trichoderma)或曲霉属(aspergillus)的孢子。本文使用的名称巴斯德毕赤酵母(p.pastoris)与名称komagataellapastoris具有相同的含义,其中巴斯德毕赤酵母是较老的名称而k.pastoris是系统更新的名称(yamada等,(1995)‘thephylogeneticrelationshipsofmethanol-assimilatingyeastsbasedonthepartialsequencesof18sand26sribosomalrnas:theproposalofkomagataellagen.nov.(saccharomycetaceae)’bioscience,biotechnologyandbiochemistry,vol.59,issue3,pp.439-444)。值得注意的是,最近已将komagataellapastoris的种重新分配为komagataellaphaffii(kurtzman(2009)“biotechnologicalstrainsofkomagataella(pichia)pastorisarekomagataellaphaffiiasdeterminedfrommultigenesequenceanalysis.”jindmicrobiolbiotechnol.36(11):1435-8)。本文所用的komagataellaphaffii可以例如涉及菌株komagataellaphaffiicbs7435、komagataellaphaffiigs115或komagataellaphaffiijc308。
本发明还涉及本文所述的α/β-水解酶用于降解玉米赤霉烯酮(zen)的用途。
zen是一种具有以下结构式的非甾类雌激素大环内酯,其通过聚酮化合物代谢途径合成:
并且,根据iupac命名法,其名称为(2e,11s)-15,17-二羟基-11-甲基-12-氧杂双环[12.4.0]十八碳-1(18),2,14,16-四烯-7,13-二酮。
然而,在自然界中还存在许多zen衍生物,其可通过zen的酶促或化学修饰而形成。例子包括由真菌、植物或哺乳动物代谢形成的糖苷zen缀合物或含硫酸酯的那些,以及在人或动物机体中形成的zen代谢物,等等。在下文中,将zen衍生物理解为天然存在的的或者通过化学或生物化学合成而合成的zen缀合物或zen代谢物,但特别地是指α-玉米赤霉烯醇(α-zel;(2e,7r,11s)-7,15,17-三羟基-11-甲基-12-氧杂双环[12.4.0]十八碳-1(18),2,14,16-四烯-13-酮)、β-玉米赤霉烯醇(β-zel;(2e,7s,11s)-7,15,17-三羟基-11-甲基-12-氧杂双环[12.4.0]十八碳-1(18),2,14,16-四烯-13-酮)、α-玉米赤霉醇(α-zal;(7r,11s)-7,15,17-三羟基-11-甲基-12-氧杂双环[12.4.0]十八碳-1(18),14,16-三烯-13-酮)、β-玉米赤霉醇(β-zal;(7s,11s)-7,15,17-三羟基-11-甲基-12-氧杂双环[12.4.0]十八碳-1(14),15,17-三烯-13-酮)、玉米赤霉烯酮-14-硫酸酯(z14s;[(2e,11s)-15-羟基-11-甲基-7,13-二氧代-12-氧杂双环[12.4.0]十八碳-1(18),2,14,16-四烯-17-基]硫酸氢酯)、玉米赤霉烯酮-14-糖苷(z14g;(2e,11s)-15-羟基-11-甲基-17-[(3r,4s,5s,6r)-3,4,5-三羟基-6-(羟甲基)-四氢吡喃-2-基]氧基-12-氧杂双环[12.4.0]十八碳1(18),2,14,16-四烯-7,13-二酮)以及玉米赤霉酮(zan;(11s)-15,17-二羟基-11-甲基-12-氧杂双环[12.4.0]十八碳-1(18),14,16-三烯-7,13-二酮)。
由于其高的化学和物理稳定性,还可以在经加工的食品和动物饲料产品比如面包或啤酒中检测到zen以及zen衍生物,尤其是α-zel、β-zel、z14s、α-zal、β-zal、z14g和zan。
采用序列id号为seqidno:1至6的多肽中的任一个成功水解了zen和zen衍生物。按照下述反应机制,认为zen或其衍生物的水解发生在酯基团处:
通过本发明的α/β-水解酶来进行将zen水解形成无毒的水解的玉米赤霉烯酮(hzen)和/或水解的zen衍生物。认为hzen至脱羧的水解的zen(dhzen)和/或脱羧的水解的zen衍生物的进一步的脱羧是自发发生的。
本文所述的α/β-水解酶能够且适用于降解zen。例如,所述α/β-水解酶可适用于不依赖于氧地且无辅助因子地水解裂解zen和/或其衍生物的酯基。
可以通过在37℃的温度下将本发明的α/β-水解酶添加至具有0.1mg/ml牛血清白蛋白的teorellstenhagen缓冲液(ph7.5)中来测量zen降解。反应中的初始底物浓度为15.71μmzen。可以使用hplc或本领域技术人员熟知的其他方法从自反应中提取的样品中检测zen、hzen、dhzen和/或其它衍生物。
特别地,可按照如下在ph7.5的含有0.1mg/ml的牛血清白蛋白的样品缓冲液(teorellstenhagen缓冲液(stenhagen&teorell.(1938)nature141,415)中于37℃的温度下达3小时来测量zen降解。将多肽/酶用样品缓冲液稀释并放置于冰上待用。作为阴性对照,温育含有5μg/mlzen的样品缓冲液。对于降解组(approach),将含有5μg/mlzen的样品缓冲液与多肽/酶溶液混合以达到最终酶浓度,该最终酶浓度在3小时内将可用的zen降解至90%至100%的程度。通过向降解组中加入多肽或酶,开始反应。未向阴性对照中添加酶。在各自的反应开始后立即涡旋约2秒,并将0h的样品(100μl)转移到新的反应管中。将反应温育在37℃下的预热的水浴中,将样品通过99℃下温育10分钟来热灭活,离心(2分钟,25℃,14674xg)并将90μl上清液转移至具有插入件的hplc小瓶中。将样品储存在4℃下直至hplc-dad测量。在0.5、1.0、2.0和3.0小时后重复取样。
可以通过如vekiru等(vekiru等,(2016)‘isolationandcharacterisationofenzymaticzearalenonehydrolysisreactionproducts’worldmycotoxinjournal9:353-363)中所述的hplc-dad分析zen、hzen和dhzen的浓度。在配有于274nm下操作的二极管阵列检测器(dad)的agilent1100系列hplc上进行分析。当通过采用溶剂a:20%甲醇水溶液+5mm乙酸铵和溶剂b:90%甲醇水溶液+5mm乙酸铵,和采用以下梯度:0-0.1min0%b相、0.1-3min线性增加至90%b相,3-5min线性增加至100%b相并持续1.9min,然后在0.1min内降至0%b相,在zorbaxsb-aq、4.6×150mm、5μm柱(agilenttechnologies)上于35℃下进行了分离时,分析物的保留时间对zen而言为7.03min,对hzen而言为5.17min和对dhzen而言为5.95min。在开始下一次运行之前,将柱修复2.0min。将流速设定为0.8ml/min并将注入体积设定为15μl。
定量基于采用zen、hzen和dhzen外标的校准。单位每升(u/l)的酶活性由zen降解的线性范围的斜率计算,所述zen降解的线性范围的斜率由样品中zen浓度对采样时间点作的图确定。为了以u/l确定样品中酶活性的量,在如上所述的图中线性范围的斜率可以以μmzen/小时计算并除以60以确定μm/min。通过考虑可能的稀释度并通过将这些适当的稀释因子囊括在计算中,可以以u/l确定样品中的酶活性。以下实例用于说明:如果线性范围的斜率为10μm/h,则未稀释样品的酶活性为0.17u/l;通过10/60=0.17计算得到。
在本文的上下文中,应注意术语“单位”或“u”是指酶催化活性的量度,并将其定义为在限定条件下每分钟反应的或裂解的底物(在此为玉米赤霉烯酮)微摩尔(μmol)量。通过酶或多肽溶液的“活性”定义了酶或多肽溶液的酶浓度,以单位/毫升(u/ml)或单位/升(u/l)溶液表示。
本发明还涉及一种包含本文所述的α/β-水解酶的组合物。所述组合物可以为食品或饲料添加剂或食品或饲料产品。
制备此类食品和/或饲料组合物的方法是本领域技术人员已知的,并特别描述于wo99/35240中。
本发明还涉及一种本文所述的α/β-水解酶或组合物,其用于在治疗或预防疾病中使用。此类疾病可以是影响激素平衡的疾病,比如雌激素平衡,特别是zen导致的霉菌毒素症。
本发明还涉及一种包含本文所述的α/β-水解酶或组合物的试剂盒。
本发明的特征还在于以下项目:
1、一种用于增加α/β-水解酶的稳定性的方法,所述α/β-水解酶包含对应于seqidno:1、3、4、5的位置145至218的序列,和/或包含对应于seqidno:2的位置144至217的序列,和/或包含对应于seqidno:6的位置161至235的序列,和/或包含与对应于seqidno:1、3、4、5的位置145至218的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:2的位置144至217的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:6的位置161至235的序列具有58%或更高的序列同一性的序列(cap-结构域;与seqidno:1、2、3、4、5、6的cap-结构域有58%的同一性),所述方法包括:
在以下位置处置换至少一个氨基酸:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,采用这样的氨基酸置换所述氨基酸:所采用的氨基酸具有比被置换的氨基酸更负的亲水指数,
其中所述亲水指数由kyte和doolittle亲水指数确定,
由此获得具有增加的稳定性的α/β-水解酶。
2、根据项目1所述的方法,其中某种氨基酸的亲水指数为:
3、根据项目1或2所述的方法,其中所述方法包括在以下位置处置换至少一个氨基酸:
-对应于seqidno:1的位置185至191的位置,和/或
-对应于seqidno:2的位置184至190的位置,和/或
-对应于seqidno:3、4或5的位置185至191的位置,和/或
-对应于seqidno:6的位置201至208的位置。
4、根据前述项目中任一项所述的方法,其中采用选自r、k、d、q、d、n、e、p、g、t、s或h的氨基酸置换所述氨基酸。
5、根据前述项目中任一项所述的方法,其中采用选自r、k、n、q、d、e、h、p、y、w、s、t、g、a、m、c、f、l或v的氨基酸置换所述氨基酸。
6、根据前述项目中任一项所述的方法,其中采用选自r、k、n、q、d、e、h、p、y、w、s、t或g的氨基酸置换所述氨基酸。
7、根据前述项目中任一项所述的方法,其中采用选自r、k、n、q、d、e、h或p的氨基酸置换所述氨基酸。
8、根据前述项目中任一项所述的方法,其中采用选自s、p、r、d、h、g或n的氨基酸置换所述氨基酸。
9、根据前述项目中任一项所述的方法,其中采用选自r、d、h、g、n或p的氨基酸置换所述氨基酸。
10、根据前述项目中任一项所述的方法,其中采用选自r、d、h、g或n的氨基酸置换所述氨基酸。
11、根据前述项目中任一项所述的方法,其中采用选自p、s、r或h的氨基酸置换所述氨基酸。
12、根据前述项目中任一项所述的方法,其中采用选自r或n的氨基酸置换所述氨基酸。
13、根据前述项目中任一项所述的方法,其中所述氨基酸置换选自以下的一个或多个:v→a、g→r、g→s、a→p、a→r、a→d、a→h、a→n、a→g、s→d、p→h、m→d、g→e、i→a、i→v、h→n和/或q→k。
14、根据前述项目中任一项所述的方法,其中所述氨基酸置换选自以下的一个或多个:v160a、g185r、g185s、a186p、a186r、a188d、a188h、a188n、a188g、a188r、s189d、p190h、m191d、g199e、i200a、i200v、h203n和/或q205k。
15、根据前述项目中任一项所述的方法,其中所述氨基酸置换选自以下的一个或多个:v→a、g→r、g→s、a→p、a→r、a→d、a→h、a→n、a→g、s→d、p→h、m→d、g→e、i→a、i→v、h→n和/或q→k。
16、根据前述项目中任一项所述的方法,其中所述氨基酸置换选自g185r、a186r、a188r、a188d、a188h、a188n和/或m191d。
17、根据前述项目中任一项所述的方法,其中所述增加的稳定性为gravy值的降低,ph稳定性的增加和/或温度稳定性的增加。
18、根据前述项目中任一项所述的方法,其中所述gravy值通过将全部氨基酸的亲水值(指数)之和除以序列中氨基酸残基的数量来计算(根据kyte和doolittle的计算)。
19、根据前述项目中任一项所述的方法,其中所述gravy值通过将序列中全部氨基酸的亲水值(指数)之和除以序列中氨基酸的总数来计算。
20、根据前述项目中任一项所述的方法,其中采用选自r、d、h、g、n或p的氨基酸置换所述氨基酸。
21、根据前述项目中任一项所述的方法,其中所述方法包括在以下位置处置换至少一个氨基酸:
-对应于seqidno:1的位置185至191的位置,或
-对应于seqidno:2的位置184至190的位置,或
-对应于seqidno:3、4或5的位置185至191的位置,或
-对应于seqidno:6的位置201至208的位置,
和其中采用选自r、d、h、g、n或p的氨基酸置换所述氨基酸。
22、一种通过前述项目中任一项所述的方法能够获得的α/β-水解酶。
23、一种α/β-水解酶,所述α/β-水解酶包含对应于seqidno:1、3、4、5的位置145至218的序列,和/或包含对应于seqidno:2的位置144至217的序列,和/或包含对应于seqidno:6的位置161至235的序列,和/或包含与对应于seqidno:1、3、4、5的位置145至218的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:2的位置144至217的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:6的位置161至235的序列具有58%或更高的序列同一性的序列(cap-结构域;与seqidno:1、2、3、4、5、6的cap-结构域有58%的同一性),
其中所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,所述α/β-水解酶具有的gravy值比具有seqidno:1的多肽序列的α/β-水解酶的gravy值负至少0.6%。
24、一种具有多肽序列的α/β-水解酶,所述多肽序列包含对应于seqidno:6的位置161至235的序列,或包含与对应于seqidno:6的位置161至235的序列具有大于58%的序列同一性的序列,
其中,所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,所述α/β-水解酶具有的gravy值比具有seqidno:6的多肽序列的α/β-水解酶的gravy值负至少0.6%。
25、一种具有多肽序列的α/β-水解酶,所述多肽序列包含对应于seqidno:1、3、4、5的位置145至218的序列,和/或包含对应于seqidno:2的位置144至217的序列,和/或包含对应于seqidno:6的位置161至235的序列,和/或包含与对应于seqidno:1、3、4、5的位置145至218的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:2的位置144至217的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:6的位置161至235的序列具有58%或更高的序列同一性的序列(cap-结构域;与seqidno:1、2、3、4、5、6的cap-结构域有58%的同一性),包含
其中所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置185至191的位置,或
-对应于seqidno:2的位置184至190的位置,或
-对应于seqidno:3、4或5的位置185至191的位置,或
-对应于seqidno:6的位置201至208的位置,
其中,所述氨基酸置换选自v→a、g→r、g→s、a→p、a→r、a→d、a→h、a→n、a→g、s→d、p→h、m→d、g→e、i→a、i→v、h→n、q→k、f→y和/或v→c,和/或
其中采用选自p、r、d、h、g或n的氨基酸置换所述氨基酸,优选地,所采用的氨基酸选自r、d、h、g或n,更优选地,所采用的氨基酸选自r或n。
26、根据前述项目中任一项所述的α/β-水解酶,其中所述α/β-水解酶包含以下氨基酸置换:
-g→r和a→n,优选g185r和a188n;
-g→s和a→r,优选g185s和a188r;
-g→r、a→r、a→h、s→d、p→h和m→d,优选g185r、a186r、a188h、s189d、p190h和m191d;
-v→a、g→r、a→n、g→e、i→a、h→n和q→k,优选v160a、g185r、a188n、g199e、i200a、h203n和q205k;
-v→a、g→s、a→r、g→e、i→a、h→n和q→k,优选v160a、g185s、a188r、g199e、i200a、h203n和q205k;
-v→a、g→e、i→a、h→n和q→k,优选v160a、g199e、i200a、h203n和q205k;
-v→a、g→r、a→r、a→h、g→e、i→a、h→n和q→k,优选v160a、g185r、a186r、a188h、g199e、i200a、h203n和q205k,和/或
-v→a、g→r、a→r、a→h、g→e、i→v、h→n和q→k,优选v160a、g185r、a186r、a188h、g199e、i200v、h203n和q205k;
-v→a、g→r、a→r、a→h、s→d、p→h、m→d、g→e、i→v、h→n和q→k,优选v160a、g185r、a186r、a188h、s189d、p190h、m191d、g199e、i200v、h203n和q205k;和/或
-f→y和v→c,优选f183y和v197c。
27、一种前述项目中任一项所述的α/β-水解酶用于降解玉米赤霉烯酮(zen)的用途。
28、一种包含前述项目中任一项所述的α/β-水解酶的组合物,优选地,所述组合物为食品或饲料添加剂或食品或饲料产品。
29、根据前述项目中任一项所述的α/β-水解酶或组合物,其用于在治疗或预防疾病中使用。
30、包含前述项目中任一项所述的α/β-水解酶或组合物的试剂盒。
31、一种用于增加α/β-水解酶的稳定性的方法,所述α/β-水解酶包含对应于seqidno:1、3、4、5的位置160至205的序列,和/或包含对应于seqidno:2的位置159至204的序列,和/或包含对应于seqidno:6的位置176至222的序列,和/或包含与对应于seqidno:1、3、4、5的位置160至205的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:2的位置159至204的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:6的位置176至222的序列具有58%或更高的序列同一性的序列(vi-结构域;与seqidno:1、2、3、4、5、6的vi-结构域有58%的同一性),所述方法包括:
在以下位置处置换至少一个氨基酸:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,采用这样的氨基酸置换所述氨基酸:所采用的氨基酸具有比被置换的氨基酸更负的亲水指数,
其中所述亲水指数由kyte和doolittle亲水指数确定,
由此获得具有增加的稳定性的α/β-水解酶。
32、一种用于增加α/β-水解酶的稳定性的方法,所述α/β-水解酶包含seqidno:1、2、3、4、5或6的序列,所述方法包括
在以下位置处置换至少一个氨基酸:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,采用这样的氨基酸置换所述氨基酸:所采用的氨基酸具有比被置换的氨基酸更负的亲水指数,
其中所述亲水指数由kyte和doolittle亲水指数确定,
由此获得具有增加的稳定性的α/β-水解酶。
33、一种α/β-水解酶,所述α/β-水解酶包含对应于seqidno:1、3、4、5的位置160至205的序列,和/或包含对应于seqidno:2的位置159至204的序列,和/或包含对应于seqidno:6的位置176至222的序列,和/或包含与对应于seqidno:1、3、4、5的位置160至205的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:2的位置159至204的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:6的位置176至222的序列具有58%或更高的序列同一性的序列(vi-结构域;与seqidno:1、2、3、4、5、6的vi-结构域有58%的同一性),其中所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置160至205的位置,或
-对应于seqidno:2的位置159至204的位置,或
-对应于seqidno:3、4或5的位置160至205的位置,或
-对应于seqidno:6的位置176至222的位置,
其中,所述α/β-水解酶具有的gravy值比具有seqidno:1的多肽序列的α/β-水解酶的gravy值负至少0.6%。
34、一种具有多肽序列的α/β-水解酶,所述多肽序列包含对应于seqidno:1、3、4、5的位置145至218的序列,和/或包含对应于seqidno:2的位置144至217的序列,和/或包含对应于seqidno:6的位置161至235的序列,和/或包含与对应于seqidno:1、3、4、5的位置145至218的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:2的位置144至217的序列具有58%或更高的序列同一性的序列,和/或包含与对应于seqidno:6的位置161至235的序列具有58%或更高的序列同一性的序列(cap-结构域;与seqidno:1、2、3、4、5、6的cap-结构域有58%的同一性),包含
其中所述多肽序列包含在以下位置处的至少一个氨基酸置换:
-对应于seqidno:1的位置185至191的位置,或
-对应于seqidno:2的位置184至190的位置,或
-对应于seqidno:3、4或5的位置185至191的位置,或
-对应于seqidno:6的位置201至208的位置,
其中所述氨基酸置换选自v→a、g→r、g→s、a→p、a→r、a→d、a→h、a→n、a→g、s→d、p→h、m→d、g→e、i→a、i→v、h→n和q→k,和/或
其中采用选自p、r、d、h、g或n的氨基酸置换所述氨基酸,优选地,所采用的氨基酸选自r、d、h、g或n,更优选地,所采用的氨基酸选自r或n。
****
需要指出的是,除非上下文另有明确指示,本文所用单数形式“一个(a)”、“一种(an)”和“所述/该(the)”包括所提及的复数。因此,例如,提及的“一种试剂”包括一种或多种这样不同的试剂,提及“所述方法”包括提及本领域普通技术人员已知的可修饰或置换本文所述的方法的等效步骤和方法。
除非另有说明,一系列元素之前的术语“至少”将被理解为是指该系列中的每一个元素。本领域技术人员将认识到或采用常规实验能够确定本文所述的本发明的具体实施方案的许多等价方案。本发明旨在涵盖这种等价方案。
本文中使用的术语“和/或”均包括“和”、“或”以及“由所述术语连接的元素的所有或任何其它组合”的含义。
术语“小于”或反义词“大于”不包括具体数字。
例如,小于20意指小于所示出的数字。类似地,大于或高于意指大于或高于示出的数字,例如大于80%意指大于或高于示出的数字80%。
本说明书及随后的权利要求中,除非上下文有其它要求,词语“包括/包含(comprise)”以及变形词语诸如“包括/包含(comprises)”和“包括/包含(comprising)”将被理解为意指包括陈述的整数或步骤或者整数或步骤的集合,但并不排除任何其它的整数或步骤或者整数或步骤的集合。当本文使用时,术语“包括/包含(comprising)”能够用术语“含有(containing)”替代或有时在本文中使用时用术语“具有(having)”替代。当在本文中使用时,“由...组成”排除未说明的任何元素、步骤或成分。
术语“包括(including)”意指“包括但不限于”。“包括(including)”与“包括但不限于”可互换使用。
应当理解,本发明不限于本文所描述的特定方法、操作程序、材料、试剂和物质等并由此可以改变。本文所用的术语仅用于描述具体的实施方案的目的,并不意在限制本发明的范围,本发明的范围仅由权利要求限定。
贯穿本说明书的文本所引用的所有出版物(包括所有的专利、专利申请、科学出版物、说明书等),无论在前还是在后,均以其全文并入本文。本文无任何内容将被解释为承认由于在先发明,本发明无权早于这样的公开内容。就通过引入并入的材料与本说明书矛盾或不一致来说,本说明书将优先于任何这样的材料。
本文所引用的所有文件及专利文件的内容均以其全文并入本文。
本申请中使用了以下序列。
表2:本申请中使用的序列。可以位于本文所述的vi-结构域侧区的基序示于seqidno:7和8。可以位于本文所述的cap-环侧区的基序示于seqidno:9、10、11。基序中的“x”可以是任何氨基酸。seqidno:9的基序中的第二个氨基酸可以是f或y,表示为“(f/y)”。vi-结构域和cap-环均包含在cap-结构域中,而例如ollis等,(1992)‘thealpha/betahydrolasefold‘proteineng.5(3):197-211所述,cap-结构域可位于β-折叠和α-螺旋之间例如α/β-水解酶的b6和ad之间的偏移(excursion)内。vi-结构域和cap-环以seqidno:1-6中的下划线示出。
通过以下仅出于说明性目的的实施例可以更好地理解本发明及其优势。这些实施例并非意在以任何方式限制本发明的范围。
本发明的实施例
实施例1:编码裂解玉米赤霉烯酮的多肽的多核苷酸的修饰、克隆和表达
使用quikchangesite-directedmutagenesiskit(stratagene),按照制造商的说明书,借助于pcr手段,通过核苷酸序列的突变来实施氨基酸置换、插入或缺失。可选地,还合成了完整的核苷酸序列(例如通过thermofisherscientific的geneartgenesynthesis)。通过标准方法将通过pcr诱变产生的核苷酸序列和从geneart获得的那些整合到用于在大肠杆菌(e.coli.)中表达的表达载体中。用所述表达载体转化大肠杆菌bl21(de3)并使所述核苷酸序列在该菌株中表达(j.m.cregg,pichiaprotocols,secondedition,isbn-10:1588294293,2007;j.sambrook等,2012,
实施例2:降解zen的α/β-水解酶、其cap-结构域、其vi-结构域和cap-环的测定
为了测定氨基酸序列是否是降解zen的α/β-水解酶,将目标序列与seqidno:1的序列比对以确定序列同一性。此外,针对目标序列进行拓扑预测和同源性建模。使用clcsequenceviewer7.8.1,采用以下参数实施序列比对:空位开放罚分:10.0,空位延伸罚分:1.0,终止空位罚分:任意其它,比对:非常精确。
通过采用以下参数的同源性建模,用yasarastructure16.7.22(1993-2016byelmarkrieger,bioinformatics30,2981-2982)实施拓扑预测和同源性建模:psi-blast迭代:3,psi-blaste-值:0.5,低聚状态:4,模板:5,具有相同的序列:1,每个模板对齐:5,建模速度:快,末端延伸:10,环样品:50,采用pssp:是。在yasara同源性建模报告中记录了通过psi-pred二级结构预测算法(jones(1999)‘proteinsecondarystructurepredictionbasedonposition-specificscoringmatrices’j.mol.biol.292:195-202)的拓扑预测。记录全部得到的同源性模型的z-得分。将所得到的具有最佳z-得分的模型用于结构分析以及α/β-水解酶的结构特征和例如由ollis等,(1992)‘thealpha/betahydrolasefold‘proteineng.5(3):197-211所述的偏移的确定。
为了识别新的目标序列中的cap-结构域、vi-结构域和cap-环,根据ollis等,(1992)‘thealpha/betahydrolasefold‘proteineng.5(3):197-211中的图2a来标记α/β-水解酶核心结构域的二级结构。cap-结构域、vi-结构域以及cap-环位于降解zen的α/β-水解酶的b6与ad之间的偏移内。cap-结构域可以起始于α/β-水解酶核心结构域的b6β-链的c末端后不远处,并且可以持续至α/β-水解酶核心结构域的adα-螺旋的n末端起始处。vi-结构域是cap-结构域的一部分并起始于存在于cap-结构域中的qxagp基序之后的第一个氨基酸并且持续至eydpe基序之前的最后一个氨基酸,其中eydpe基序不是vi-结构域的一部分。cap-环是vi-结构域的一部分并起始于存在于vi-结构域中的g(f/y)xxaa基序之后的第一个氨基酸并且持续至arxf基序(或对于seqidno:6而言的qlfp基序)之前的最后一个氨基酸,其中arxf基序(或对于seqidno:6而言的qlfp基序)不是cap-环的一部分。例如,cap-结构域、vi-结构域和cap-环的位置由如本文所述的针对seqidno:1-6的多肽的位置确定并示于图1。
实施例3:总平均亲水性(gravy)值的确定
多肽氨基酸序列的总平均亲水性(gravy)值定义为全部氨基酸的亲水值(kyte和doolittle,1982,本文中引用)之和除以多肽长度,所述多肽长度对应于多肽的氨基酸总数。采用https://web.expasy.org/protparam上的protparam程序(gasteigere.等;proteinidentificationandanalysistoolsontheexpasyserver,injohnm.walker(ed):theproteomicsprotocolshandbook,humanapress,2005,pp.571-607)计算多肽和所定义的多肽的部分的gravy值。将seqidno:1的cap-结构域定义为自氨基酸位置145至218(包括这两个位置)的部分,将seqidno:1的vi-结构域定义为自氨基酸位置160至205(包括这两个位置)的氨基酸序列,和将seqidno:1的cap-环定义为自氨基酸位置185至191(包括这两个位置)的氨基酸序列的部分。对于整个seqidno:1的多肽,计算的gravy值是-0.167;对于seqidno:1的cap-结构域,gravy值是-0.284;对于seqidno:1的vi-结构域,gravy值是-0.043;和对于seqidno:1的cap-结构域中的cap-环,gravy值是+0.271。将seqidno:6的cap-结构域、vi结构域和cap-环分别定义为自氨基酸位置161-235、176-222、201-208(包括所述范围的这两个位置)的部分。对于整个seqidno:6的多肽,计算的gravy值是-0.192;对于seqidno:6的cap-结构域,gravy值是-0.388;对于seqidno:6的vi-结构域,gravy值是-0.468;和对于seqidno:6的cap-环,gravy值是-0.362。
由至少一个突变导致的多肽的整个氨基酸序列相对于seqidno:1或6的未突变/未置换的多肽的整个氨基酸序列的gravy值降低百分比通过如下计算:将两个gravy值之差除以未突变的多肽的gravy值,再乘以100。为了说明,将针对seqidno:1变体v160a的计算示于此处:((-0.174)-(-0.167))/(-0.167)x100=4.2%。其它实例的结果列于图2a和2e中。
由cap-结构域中的突变导致的cap-结构域相对于seqidno:1或6的未突变的cap-结构域的序列的gravy值降低百分比通过如下计算:将两个gravy值之差除以未突变的cap-结构域的gravy值,再乘以100。为了说明,将针对seqidno:1变体v160a的计算示于此处:((-0.316)-(-0.284))/(-0.284)x100=11.3%。其它实例的结果列于图2b和2f中。
由vi-结构域中的突变导致的vi-结构域相对于seqidno:1或6的未突变的vi-结构域的序列的gravy值降低百分比通过如下计算:将两个gravy值之差除以未突变的vi-结构域的gravy值,再乘以100。为了说明,将针对seqidno:1变体g185r的计算示于此处:((-0.133)-(0.043))/(-0.043)x100=209.3%。其它实例的结果列于图2c和2g中。
由cap-环中的突变导致的cap-环相对于seqidno:1或6的未突变的cap-环的序列的gravy值降低百分比通过如下计算:将两个gravy值之差除以未突变的cap-环的gravy值,再乘以100。为了说明,将针对seqidno:1变体g185r的计算示于此处:((-0.314)-(+0.271))/(+0.271)x100=-215.9%。由于亲本cap-环的gravy值为正,因此该值为负。为了简化突变的其它实例的数据表示,将百分比值在图2d中以正值示出。
实施例4:降解zen的多肽的活性的确定
采用标准方法克隆编码降解zen的多肽的相应基因,在大肠杆菌中细胞内地表达,并通过本领域技术人员已知的方法,例如通过采用frenchpresscell的裂解自大肠杆菌中分离产生的多肽。通过标准方法,例如bca方法(piercebcaproteinassaykitprod#23225)实施蛋白浓度的确定。
在ph7.5的含有0.1mg/ml的牛血清白蛋白的样品缓冲液(teorellstenhagen缓冲液(stenhagen&teorell.(1938)nature141,415)中于37℃的温度下达3小时来实施酶活性的确定。将多肽用样品缓冲液稀释并放置于冰上待用。将1500ppm(w/v)的乙腈中zen储备溶液用样品缓冲液按1:10稀释,并储存于25℃下直至进一步稀释以用于降解反应。在反应管中制备了降解组和一个阴性对照。作为阴性对照,温育含有5μg/mlzen的样品缓冲液。对于降解反应,将样品缓冲液与多肽溶液混合以达到5μg/ml最终zen浓度和最终酶浓度,该最终酶浓度在3小时内实现90%至100%的zen降解。通过向含有zen的样品缓冲液中加入多肽,开始反应。未向阴性对照中添加酶。在各自的反应开始后立即涡旋约2秒,并将0h的样品(100μl)取出并转移到新的反应管中。将反应温育在37℃下的预热的水浴中,将样品通过在99℃下温育10分钟来热灭活,离心(2分钟,25℃,14674xg)并将90μl上清液转移至具有插入件的hplc小瓶中。将样品储存在4℃下直至hplc-dad测量。在0.5、1.0、2.0和3.0小时后重复取样。
可以通过如vekiru等(vekiru等,(2016)‘isolationandcharacterisationofenzymaticzearalenonehydrolysisreactionproducts’worldmycotoxinjournal9:353-363)中所述的hplc-dad分析zen、hzen和dhzen的浓度。在配有于274nm下操作的dad检测器的agilent1100系列hplc上进行分析。当通过采用溶剂a:20%甲醇水溶液+5mm乙酸铵和溶剂b:90%甲醇水溶液+5mm乙酸铵,和采用以下梯度:0-0.1min0%b相、0.1-3min线性增加至90%b相,3-5min线性增加至100%b相并持续1.9min,然后在0.1min内降至0%b相,在zorbaxsb-aq、4.6×150mm、5μm柱(agilenttechnologies)上于35℃下进行了分离时,分析物的保留时间对zen而言为7.03min,对hzen而言为5.17min和对dhzen而言为5.95min。在开始下一次运行之前,将柱修复2.0min。将流速设定为0.8ml/min并将注入体积设定为15μl。定量基于采用zen、hzen和dhzen的外标进行的校准。单位每升(u/l)的酶活性由zen降解的线性范围的斜率计算,所述zen降解的线性范围的斜率由样品中zen浓度对采样时间点作的图确定。为了以u/l确定样品中酶活性的量,在如上所述的图中线性范围的斜率可以以μmzen/小时计算并除以60以确定μm/min。通过考虑可能的稀释度并通过将这些适当的稀释因子囊括在计算中,可以以u/l确定样品中的酶活性。以下实例用于说明:如果线性范围的斜率为10μm/h,则未稀释样品的酶活性为0.17u/l;通过10/60=0.17计算得到。
实施例5:降解zen的多肽的温度稳定性
降解zen的多肽的产生和量化按照如上实施例中所述实施。为了评估温度稳定性,将降解zen的酶于不同温度下在缓冲溶液中温育,然后测试这些酶在最佳条件下降解zen的能力。在整个热温育期间取样,并相对于未经热处理的对照计算残余活性。
对于温度稳定性测试,采用样品缓冲液(ph7.5的含有0.1mg/ml牛血清白蛋白的teorellstenhagen缓冲液)将多肽稀释至0.001526923u/ml的浓度并放置于冰上待用。将40份50μl稀释的多肽溶液的等分试样转移至4组12-连管(例如购自starlab)的管中,其中每组连管的第一个管和每组连管的最后一个管未使用而是空置。所述连管采用12-连帽(例如购自starlab)密封。作为阳性对照,将4份50μl稀释酶溶液的等分试样转移至4个pcr管中。将全部pcr管和连管均在冰上放置直至开始温度温育步骤。作为阴性对照,将4份50μl样品缓冲液的等分试样转移至4个pcr管中。这些管储存在25℃下。
在选定温度+/-10℃下,将4组12-连管在具有梯度功能(例如eppendorfmastercycler梯度)的预热pcr循环仪中温育。pcr循环仪自动计算沿pcr循环仪加热块的温度梯度(选定温度的+/-10℃)。将含有阳性对照的pcr管在冰上温育,将含有阴性对照的pcr管在25℃下温育。在0、5、10和20分钟后,将1组pcr连管和1个阴性对照管转移以放置在冰上直至温育结束,所述温育结束即温育开始后20分钟。
在全部温育步骤完成并将所有连管和管均放置在冰上之后,开始zen降解分析。
制备zen降解分析缓冲液(teorellstenhagen缓冲液,ph7.5,含有0.1mg/ml牛血清白蛋白和5.3ppmzen),并将660μl分析缓冲液的等分试样转移至48个反应管中。将所述管密封并保存在25℃下直至开始zen降解分析。对于降解分析,将来自pcr连管的40个温度处理样品的每一个的40μl、4个阴性对照的每一个的40μl和4个阳性对照的每一个的40μl加入含有660μl分析缓冲液的管中,由此实现分析反应中最终zen浓度为5ppm。此外,由此还实现了多肽的终浓度以在三个小时内有效降解zen(即90%-100%的zen降解)。
通过将温度处理样品、阳性或阴性对照加入分析缓冲液中,开始降解分析。将zen降解反应在37℃下的预热的水浴中温育。在降解反应开始后立即通过涡旋混合约2秒,并将100μl的0h样品转移到新的反应管中。在0.5、1.0、2.0和3.0小时后,从zen降解分析反应中抽取其它的样品。一旦将样品自降解反应中取出,就通过在99℃下温育10分钟使该样品中的酶热灭活。随后,将管离心(2分钟,25℃,14674xg)并将90μl上清液转移至具有插入件的hplc小瓶中。将这些hplc小瓶储存在4℃下直至按照实施例4所述进行hplc-dad测量。
采用通过zen降解样品的hplc-dad分析确定的zen浓度的线性降低,以例如单位/升(u/l)或单位/毫升(u/ml)来计算酶活性。将一个单位定义为在所述条件下在1分钟内降解1μmolzen的酶活性的量。在不同温度下温育0、5、10和20分钟后的残余活性计算如下:温度处理样品中的酶活性除以0-分钟样品的酶活性的平均值,再乘以100。
将温度稳定性(t(50%))定义为这样的温度:在该温度下温育10分钟后的多肽具有相比于阳性对照的50%残余活性。以下实例用于说明:亲本酶在冰上温育10分钟后具有50u/ml的酶活性,在59.3℃下温育10分钟后具有25u/ml的活性,则t(50%)值为59.3℃。如果酶变体具有61.0℃的t(50%)值,则温度稳定性(t(50%))相比于亲本酶的相对增加为2.9%。这是将两个t(50%)值之间的差值1.7℃除以亲本酶的t(50%)值59.3℃再乘以100得到的。
如图3a和3b所示,单个突变以及突变的组合示出了温度稳定性的增加。
实施例6:降解zen毒素的多肽的ph稳定性
将降解zen的多肽在不同ph值的缓冲溶液中温育1小时,然后测试这些多肽在最佳条件下降解zen的能力。定期取样并采用hplc-dad测量分析zen、hzen和dhzen的浓度。
用于实验的ph值为ph3.5、4.0、4.2、4.4、4.6、4.8、5.0和6.0。将测试的多肽转移至8个含有8种不同ph的温育缓冲液的样品管中。所述温育缓冲液为不含奶且半浓缩的fedstatesimulatedgastricfluid中间缓冲液(jantratid等,(2008)’dissolutionmediasimulatingconditionsintheproximalhumangastrointestinaltract:anupdate.’pharmres.2008jul;25(7):1663-76),设定为ph3.5、4.0、4.2、4.4、4.6、4.8、5.0和6.0。还将多肽变体的1份等分试样转移至一个含有样品缓冲液(teorellstenhagen缓冲液,ph7.5,含有0.1mg/ml牛血清白蛋白)的管中作为阳性对照。温育溶液中的测试多肽的浓度为0.001526923u/ml,体积为100μl。将管涡旋约2秒并在预热的水浴中于37℃下温育1小时。作为阴性对照,将100μl样品缓冲液在预热的水浴中于37℃下温育1小时。温育1小时后,采用最终浓度为8.72527e-05u/ml的测试多肽进行zen降解分析。为了开始zen降解反应,将40μl温育的样品转移至660μl分析缓冲液(teorellstenhagen缓冲液,ph7.5,含有0.1mg/ml牛血清白蛋白和5.3ppmzen)中。分析缓冲液的加入确保所有样品中ph值恒定为ph7.5。在每个反应开始后立即涡旋约2秒并将0h样品(100μl)取出并转移至新的反应管中。将反应在37℃下的预热的水浴中温育,将自反应中取出的样品通过在99℃下温育10分钟来热灭活,离心(2分钟,25℃,14674xg)并将90μl上清液转移至具有插入件的hplc小瓶中。将样品储存在4℃下直至hplc-dad测量。在0.5、1.0、2.0和3.0小时后自每个降解分析反应中取样。按照实施例4所述通过hplc-dad分析zen、hzen和dhzen并按照实施例4所述计算活性。
将ph稳定性的增加定义为与未突变的亲本酶溶液在相同处理后的残余活性相比,在ph4.0下温育后的多肽溶液的残余活性的增加。通过ph-处理的多肽溶液的活性与相同多肽变体溶液在ph7.5下温育后的活性的比较来定义所述残余活性。按照如下计算所述残余活性:ph-处理的样品的酶活性除以在ph7.5下温育的对照的酶活性,再乘以100。以下实例用于说明:如果多肽样品在ph4.0下温育后的酶活性是0.5u/l且该多肽样品在ph7.5下温育后的酶活性是2.7u/l,则该多肽样品的残余活性将是18.5%。此外,如果多肽变体在ph4.0下温育后的残余活性测量是18.5%,且具有seqidno:1的亲本多肽在ph4.0下温育后的残余活性测量是2.5%,则与亲本多肽相比,该多肽变体的ph稳定性增加为7.4倍。在引入如本文所述的突变后增加的ph稳定性的数据示于图4。
实施例7:多肽变体用于解除猪中zen毒性的测试
选择总共12只断奶仔猪(雌性;年龄38天-40天),并根据采用12个单独的笼子、每笼1只仔猪的实验设置进行随机分配(4组,每组3个笼子/重复)。三个测试组接受降解zen的酶,一个对照组未接受任何降解zen的酶。所有仔猪均为奥地利基因型
进入笼舍后,所有仔猪均饲喂包含以下以百分比(w/w)计的成分的饮食:29.70%大麦、10.00%小麦、9.98%玉米、0.27%菜籽油、15.30%全脂大豆、10.94%压熟玉米(maizepressurecooked)、5.00%马铃薯蛋白、5.13%葡萄糖、3.75%棕榈仁、椰子油脂、3.75%乳糖、1.35%木质纤维素、1.23%磷酸一钙、0.93%碳酸钙、0.48%氯化钠、0.25%磷酸镁、0.42%维生素/微量元素预混物、0.70%l-赖氨酸、0.30%l-苏氨酸、0.27%dl-甲硫氨酸、0.15%l-缬氨酸、0.07%l-色氨酸和0.02%甜味剂。
在实验期间,将所有组的饮食均补充zen至终浓度为500μgzen/kg食物。对于测试组,采用seqidno:1的亲本多肽及其两种多肽变体。以下列浓度测试了多肽:2.5u/kg食物、5u/kg食物、10u/kg食物和20u/kg食物。在3天的适应阶段后,以2.5u/kg食物的浓度开始应用多肽一天,然后是饮食中没有多肽且没有zen的一天清除日(awash-outday)。清除日后,非对照仔猪接受含有zen的饮食,该饮食中具有浓度为5u/kg食物的相同多肽,然后是清除日,依此类推。在试验期间,收集12小时的尿液,并且每天采集一次粪便样品。将样品储存在-20℃下直至lc-ms/ms测量。
为了使排泄的尿液体积标准化,测量了尿液样品中肌酸酐的浓度。为了确定肌酸酐含量,将尿液样品用水1:5000稀释。将尿液样品用水稀释至2.5mm肌酸酐的终浓度。将100μl稀释的尿液样品与20μl含有528u的β-葡萄糖醛酸糖苷酶的100mmpbs缓冲液混合,并在37℃下温育过夜。过夜温育后,加入380μl冷甲醇,以14674×g离心,将上清液转移到hplc小瓶中并在-20℃下储存直至分析。对于粪便样品的分析,将500mg冷冻干燥的粪便每次用5ml乙腈/水(50/50,v/v)提取三次(90、30和30分钟)。在每次提取步骤后,通过离心(10分钟,14674×g)使样品澄清。将合并的上清液的等分试样离心并通过hplc-ms/ms测量。在与6500qtrap质谱仪相连的agilent1290系列uhplc系统上进行分析。柱温设定为30℃,流速为0.25ml/min。流动相a和b分别由水/乙酸和乙腈/乙酸(均为99.9/0.1,v/v)组成。梯度以5%b0.5min开始,然后以线性增加至36%b继续直至17.0min,并在17.0至22.0min之间线性增加至100%b,然后是100%b至24.0min,之后在24.0至24.1min之间急速下降至5%b。最后,将柱在5%b下重新平衡至27.0min。尿液样品的注入体积为2μl,粪便样品的注入体积为3μl。在phenomenexkinetexc18柱(150x2.1mm,2.6μm)上进行分离。定量基于采用zen、α-zel、hzen和dhzen的外标进行的校准。α-zel是一种具有较高的雌激素活性的zen的代谢物,通过肝生物转化在猪体内产生(malekinejad等,(2006)‘hydroxysteroiddehydrogenasesinbovineandporcinegranulosacellsconvertzearalenoneintoitshydroxylatedmetabolitesalpha-zearalenolandbeta-zearalenol.vetrescommun:445-53)。所选择的反应监测(srm)参数示于图5。
除了具有seqidno:1的多肽之外,已经测试了两种具有seqidno:1的多肽的多肽变体:变体a和变体b。
与seqidno:1相比,变体a包含以下突变:v160a/g185r/a186r/a188h/g199e/i200v/h203n/q205k。
与seqidno:1相比,变体b包含以下突变:v160a/g185r/a186r/a188h/s189d/p190h/m191d/g199e/i200v/h203n/q205k。
自尿液和粪便样品的分析中得到的结果示于图6和7。与seqidno:1相比,zen加α-zel的组合浓度的变化百分比结果通过以下得出:将两组的浓度之间的差值除以具有seqidno:1的组的浓度,再乘以100。在饮食中未接受降解zen的多肽的对照组的饲喂试验过程中,zen加α-zel的浓度的增加可能是由猪的zen及zen衍生物的肝肠循环引起的并由此积累(biehl等,(1993)‘biliaryexcretionandenterohepaticcyclingofzearalenoneinimmaturepigs.’toxicolapplpharmacol.1993jul;121(1):152-9)。
实施例8:不同浓度的多肽变体用于解除仔鸡中zen毒性的测试
对于饲喂试验,采用90日龄、混合性别的仔鸡(ross308)。在两个阶段这些仔鸡饲喂两种不同的饮食。在适应期,仔鸡接受阶段1饮食(第1-14天期间),在实验期,仔鸡接受阶段2饮食(第15-28天期间)。以百分比(w/w)计的阶段1饮食的组成:55.00%玉米、29.00%大豆hp、1.00%葵花籽油、6.92%全脂大豆、0.72%大豆蛋白浓缩物、1.88%棕榈仁、椰子油脂、1.96%碳酸钙、1.89%磷酸一钙、0.35%碳酸氢钠、0.23%氯化钠、0.13%磷酸镁、0.24%维生素/微量元素预混物、0.34%l-赖氨酸、0.12%l-苏氨酸和0.24%dl-甲硫氨酸。
以百分比(w/w)计的阶段2饮食的组成:62.00%玉米、23.80%大豆hp、2.00%葵花籽油、5.53%全脂大豆、0.58%大豆蛋白浓缩物、1.50%棕榈仁、椰子油脂、1.56%碳酸钙、1.71%磷酸一钙、0.28%碳酸氢钠、0.19%氯化钠、0.10%磷酸镁、0.19%维生素/微量元素预混物、0.27%l-赖氨酸、0.10%l-苏氨酸和0.20%dl-甲硫氨酸。
对于适应期(14天),将仔鸡随机分配至3个笼子。适应期后,将仔鸡按照平均重量均匀分配至7个笼子(组)中,每个笼子8只。试验持续时间为14天。根据饲养公司的标准建议调整气候条件。每天手动喂食一次,饲料和水随时供应。饮食中zen的浓度为400μgzen/kg食物。对照组饲喂包含400μgzen/kg食物、不添加任何降解zen的酶的饮食。以下列浓度测试了多肽变体a(包含以下突变的seqidno:1:v160a/g185r/a186r/a188h/g199e/i200v/h203n/q205k):5u/kg食物、10u/kg食物、20u/kg食物、40u/kg食物、80u/kg食物和160u/kg食物。安乐死后,自试验开始时和试验结束时的仔鸡中取出嗉囊样品。将样品冷冻且冻干。为了分析zen、hzen和dhzen,称取1g每种样品至50ml管中并于25℃下在旋转震荡器上用15ml80%乙腈萃取两次,每次30min。然后将管以2300xg在25℃下离心10min,并将上清液合并至一个新的50ml管中。取1ml以2300xg在25℃下再次离心5min,并将上清液转移至一个用于lc-ms/ms测量的小瓶中。将样品在-20℃下储存直至测量并通过lc-ms/ms分析。在与6500+qtrap质谱仪相连的agilent1290系列uhplc系统上进行分析。柱温设定为30℃,流速为0.25ml/min。流动相a和b分别由水/乙酸和乙腈/乙酸(均为99.9/0.1,v/v)组成。梯度以15%b0.5min开始,然后以线性增加至60%b继续直至13.5min,并在13.5至14.0min之间急速增加至100%b,然后是100%b至16.9min,之后在16.9至17.0min之间急速下降至15%b。最后,将柱在15%b下重新平衡至20.0min。注入体积为2μl。在phenomenexkinetexc18柱(150x2.1mm,2.6μm)上进行分离。定量基于采用zen、hzen和dhzen的外标进行的校准。所选择的反应监测(srm)参数示于图8。
从来自试验结束的嗉囊样品的分析中得到的结果示于图9。zen浓度的降低百分比按照如下计算:(对照组的zen浓度减去样品组的zen浓度)除以对照组的zen浓度,再乘以100。
参考文献
altschulnucl.acidsres.25(1977),3389-3402
altschul,j.mol.evol.36(1993),290-300
altschul,j.mol.biol.215(1990),403-410
biehl等,(1993)‘biliaryexcretionandenterohepaticcyclingofzearalenoneinimmaturepigs.’toxicolapplpharmacol.1993jul;121(1):152-9
brutlagcomp.app.biosci.6(1990),237-245
burnley和gros(2012)‘phenix.ensemble_refinement:ateststudyofapoandholobace1‘computationalcrystallographynewsletter,volume4,pp.51–58
cregg,pichiaprotocols,secondedition,isbn-10:1588294293,2007
proteins-structureandmolecularproperties,2nded.,t.e.creighton,w.h.freemanandcompany,newyork(1993)
elmarkrieger,bioinformatics30,2981-2982
gasteigere.等,proteinidentificationandanalysistoolsontheexpasyserver,injohnm.walker(ed):theproteomicsprotocolshandbook,humanapress,2005,pp.571-607
henikoffproc.natl.acad.sci.,usa,89,(1989),10915
henikoff和henikoff(1992)‘aminoacidsubstitutionmatricesfromproteinblocks.‘procnatlacadsciusa.1992nov15;89(22):10915-9
jantratid等,(2008)’dissolutionmediasimulatingconditionsintheproximalhumangastrointestinaltract:anupdate.’pharmres.2008jul;25(7):1663-76
jones(1999)‘proteinsecondarystructurepredictionbasedonposition-specificscoringmatrices’j.mol.biol.292:195-202
post-translationalcovalentmodificationofproteins,b.c.johnson,ed.,academicpress,newyork(1983),pgs.1-12;seifter,meth.enzymol.182(1990),626-646
kourist等,(2010)‘thealpha/beta-hydrolasefold3dmdatabase(abhdb)asatoolforproteinengineering.’chembiochem.11(12):1635-43
kurtzman(2009)“biotechnologicalstrainsofkomagataella(pichia)pastorisarekomagataellaphaffiiasdeterminedfrommultigenesequenceanalysis.”jindmicrobiolbiotechnol.36(11):1435-8
kyte和doolittle(1983)"asimplemethodfordisplayingthehydropathiccharacterofaprotein".j.mol.biol.157(1):105–32
lenfant等,(2013)‘esther,thedatabaseoftheα/β-hydrolasefoldsuperfamilyofproteins:toolstoexplorediversityoffunctions’nucleicacidsresearch,volume41,issued1,d423–d429
malekinejad等,(2006)‘hydroxysteroiddehydrogenasesinbovineandporcinegranulosacellsconvertzearalenoneintoitshydroxylatedmetabolitesalpha-zearalenolandbeta-zearalenol.vetrescommun:445-53
mindrebo等,(2016)‘unveilingthefunctionaldiversityofthealpha-betahydrolasefoldinplants’curropinstructbiol.233–246
ollis等,(1992)‘thealpha/betahydrolasefold‘proteineng.5(3):197-211
rattan,ann.nyacad.sci.663(1992);48-62
schatzmayr和streit(2013)‘globaloccurrenceofmycotoxinsinthefoodandfeedchain:factsandfigures.’worldmycotoxinjournal6(3):213-222
sambrook等,2012,molecularcloning,alaboratorymanual4thedition,coldspringharbor
stenhagen&teorell.(1938)nature141,415
thompsonnucl.acidsres.2(1994),4673-4680
vekiru等,(2016)‘isolationandcharacterisationofenzymaticzearalenonehydrolysisreactionproducts’worldmycotoxinjournal9:353-363)
yamada等,(1995)‘thephylogeneticrelationshipsofmethanol-assimilatingyeastsbasedonthepartialsequencesof18sand26sribosomalrnas:theproposalofkomagataellagen.nov.(saccharomycetaceae)’bioscience,biotechnologyandbiochemistry,vol.59,issue3,pp.439-444)。
序列表
<110>奥地利商艾尔柏有限公司
<120>用于裂解玉米赤霉烯酮的手段和方法
<130>lc19310008
<160>11
<170>siposequencelisting1.0
<210>1
<211>309
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>peptide
<222>()..()
<223>α/β水解酶结构域
<400>1
metvalthrserproalaleuargaspvalhisvalprohisalatyr
151015
progluglnglnvalaspleuglygluilethrmetasntyralaglu
202530
alaglyaspproaspargproalavalleuleuileprogluglnthr
354045
glysertrptrpsertyrgluglualametglyleuleusergluhis
505560
phehisvaltyralavalaspleuargglyglnglyargsersertrp
65707580
thrprolysargtyrserleuaspasnpheglyasnaspleuvalarg
859095
pheilealaleuvalvallysargprovalvalvalalaglyasnser
100105110
serglyglyvalleualaalatrpleuseralatyrsermetprogly
115120125
glnleuargglyvalleucysgluaspproprophephealaserglu
130135140
leuvalproalahisglyhisservalargglnglyalaglyproval
145150155160
phegluleupheargthrtyrleuglyaspglntrpservalglyasp
165170175
trpgluglyphecysargalaalaglyalaseralaserprometala
180185190
argserphevalalaaspglyileproglnhisleuglnglutyrasp
195200205
proglutrpalaargvalphetyrgluglythrvalglyleusercys
210215220
prohisgluargmetleuglyglnvallysthrprovalleuleuthr
225230235240
hishismetargglyileaspprogluthrglyasnleuleuglyala
245250255
leuseraspgluglnalaleuargalaargargleumetaspserala
260265270
glyvalthrvalasptyrgluservalproaspalaserhismetmet
275280285
hisglnseralaproalaargtyrvalgluilephethrargtrpala
290295300
alaalaleualapro
305
<210>2
<211>308
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>peptide
<222>()..()
<223>α/β水解酶结构域
<400>2
metalaaspproalaglnargaspvaltyrvalprohisalatyrpro
151015
glulysglnalaaspleuglygluilethrmetasntyralagluala
202530
glygluproaspmetproalavalleuleuileprogluglnthrgly
354045
sertrptrpglytyrgluglualametglyleuleualagluasnphe
505560
hisvaltyralavalaspleuargglyglnglyargsersertrpala
65707580
prolysargtyrserleuaspasnpheglyasnaspleuvalargphe
859095
ilealaleuvalvallysargprovalilevalalaglyasnserser
100105110
glyglyvalleualaalatrpleuseralatyrsermetproglygln
115120125
valargglyalaleucysgluaspalaprophephealasergluleu
130135140
valthrthrcysglyhisserileargglnalaalaglyprometphe
145150155160
gluleupheargthrtyrleuglyaspglntrpservalglyasptrp
165170175
thrglytyrcysargalaalaaspalaserserserprometalaarg
180185190
tyrphevalalaaspgluileproglnhismetargglutyrasppro
195200205
glutrpalaargalaphetrpgluglythrvalalaleuhiscyspro
210215220
hisgluglnleuleuthrglnvallysthrprovalleuleuthrhis
225230235240
hismetargaspileaspproaspthrglyhisleuvalglyalaleu
245250255
seraspgluglnalaalaargalaargleuleumetgluseralagly
260265270
vallysvalasptyralaservalproaspalaleuhismetmethis
275280285
glnpheaspproproargtyrvalgluilephethrglntrpalaala
290295300
thrleualaala
305
<210>3
<211>309
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>peptide
<222>()..()
<223>α/β水解酶结构域
<400>3
metvalthrserproalaleuargaspvalhisvalprohisalatyr
151015
progluglnglnvalaspleuglygluilethrmetasntyralaglu
202530
alaglyaspproglyargproalavalleuleuileprogluglnthr
354045
glysertrptrpsertyrgluglualametglyleuleualagluhis
505560
phehisvaltyralavalaspleuargglyglnglyargsersertrp
65707580
thrprolysargtyrserleuaspasnpheglyasnaspleuvalarg
859095
pheilealaleuvalvalargargprovalvalvalalaglyasnser
100105110
serglyglyvalleualaalatrpleuseralatyrsermetprogly
115120125
glnileargglyvalleucysgluaspproprophephealaserglu
130135140
leuvalproalahisglyhisservalargglnglyalaglyproval
145150155160
phegluleupheargthrtyrleuglyaspglntrpservalglyasp
165170175
trpgluglypheargseralaalaaspalaseralaserprometala
180185190
argserphevalalaaspthrileproglnhisleulysglutyrasp
195200205
proglutrpalaargalaphetyrgluglythrvalglyleuasncys
210215220
prohisgluargmetleuasnargvalasnthrprovalleuleuthr
225230235240
hishismetargglythraspprogluthrglyasnleuleuglyala
245250255
leuseraspgluglnalaalaglnvalargargleumetgluserala
260265270
glyvallysvalasptyrgluservalproaspalaserhismetmet
275280285
hisglnseraspproalaargtyralagluileleuthrprotrpthr
290295300
alaalaleualapro
305
<210>4
<211>309
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>peptide
<222>()..()
<223>α/β水解酶结构域
<400>4
metvalthrserproalaleuargaspvalhisvalprohisalatyr
151015
progluglnglnvalaspleuglygluilethrmetasntyralaglu
202530
alaglyaspproaspargproalavalleuleuileprogluglnthr
354045
glysertrptrpsertyrgluglualametglyleuleualagluhis
505560
phehisvaltyralavalaspleuargglyglnglyargsersertrp
65707580
thrprolysargtyrserleuaspasnpheglyasnaspleuvalarg
859095
pheilealaleuvalvallysargprovalvalvalalaglyasnser
100105110
serglyglyvalleualaalatrpleuseralatyrsermetprogly
115120125
glnleuargglyvalleucysgluaspproprophephealaserglu
130135140
leuvalproalahisglyhisservalargglnglyalaglyproval
145150155160
phegluleupheargthrtyrleuglyaspglntrpservalserasp
165170175
trpgluglyphecysargalaalaglyalaseralaserprometala
180185190
argserphevalalaaspglyileproglnhisleulysglutyrasp
195200205
proglutrpalaargalaphehisgluglythrvalglyleuasncys
210215220
prohisgluargmetleuglyargvalasnthrprovalleuleuthr
225230235240
hishismetargglythraspprogluthrglyasnleuleuglyala
245250255
leuseraspgluglnalaalaglnalaargleuleumetgluserala
260265270
glyvalargvalasptyrgluservalproaspalaserhismetmet
275280285
hisglnseraspproalaargtyralagluilephethrargtrpala
290295300
alaalaleualapro
305
<210>5
<211>309
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>peptide
<222>()..()
<223>α/β水解酶结构域
<400>5
metvalthrserproalaleuargaspvalhisvalprohisalatyr
151015
progluglnglnvalaspleuglygluilethrmetasntyralaglu
202530
alaglyaspproglyargproalavalleuleuileprogluglnthr
354045
glysertrptrpsertyrgluglualametglyleuleualagluhis
505560
phehisvaltyralavalaspleuargglyglnglyargsersertrp
65707580
thrprolysargtyrserleuaspasnpheglyasnaspleuvalarg
859095
phemetalaleuvalvalargargprovalvalvalalaglyasnser
100105110
serglyglyvalleualaalatrpleuseralatyrsermetprogly
115120125
glnileargglyvalleucysgluaspproprophephealaserglu
130135140
leuvalproalahisglyhisservalargglnglyalaglyproval
145150155160
phegluleupheargthrtyrleuglyaspglntrpservalglyasp
165170175
trpgluglypheargseralaalaglyalaseralaserprometala
180185190
argserphevalalaaspthrileproglnhisleulysglutyrasp
195200205
proglutrpalaargalaphetyrgluglythrvalglyleuasncys
210215220
prohisgluargmetleuasnargvalasnthrprovalleuleuthr
225230235240
hishismetargglythraspprogluthrglyasnleuleuglyala
245250255
leuseraspgluglnalaalaglnalaargargleumetgluserala
260265270
glyvallysvalasptyrgluservalproaspalaserhismetmet
275280285
hisglnseraspproalaargtyralagluileleuthrprotrpala
290295300
alaalaleualapro
305
<210>6
<211>328
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>peptide
<222>()..()
<223>α/β水解酶结构域
<400>6
metalaglugluglythrargserglualaalaaspalaalathrgln
151015
alaargglnleuproaspserargasnilephevalserhisargphe
202530
progluargglnvalaspleuglygluvalvalmetasnphealaglu
354045
alaglyserproaspasnproalaleuleuleuleuprogluglnthr
505560
glysertrptrpsertyrgluprovalmetglyleuleualagluasn
65707580
phehisvalphealavalaspileargglyglnglyargserthrtrp
859095
thrproargargtyrserleuaspasnpheglyasnaspleuvalarg
100105110
pheilealaleuvalilelysargprovalvalvalalaglyasnser
115120125
serglyglyleuleualaalatrpleuseralatyralametprogly
130135140
glnileargalaalaleucysgluaspalaprophephealaserglu
145150155160
leuvalproalatyrglyhisservalleuglnalaalaglyproala
165170175
phegluleutyrargasppheleuglyaspglntrpserileglyasp
180185190
trplysglyphevalglualaalalysalaserproalalysalamet
195200205
glnleupheprothrproaspglualaproglnasnleulysglutyr
210215220
aspproglutrpglyargalaphephegluglythrvalalaleuhis
225230235240
cysprohisaspargmetleuserglnvallysthrproileleuile
245250255
thrhishisalaargthrileaspprogluthrglygluleuleugly
260265270
alaleuseraspleuglnalagluhisalaglnaspileileargser
275280285
alaglyvalargvalasptyrglnserhisproaspalaleuhismet
290295300
methisleupheaspproalaargtyralagluileleuthrsertrp
305310315320
seralathrleuproalaasnasp
325
<210>7
<211>5
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>peptide
<222>()..()
<223>基序
<220>
<221>misc_feature
<222>(2)..(2)
<223>x为任意氨基酸
<220>
<221>unsure
<222>(2)..(2)
<223>the'xaa'atlocation2standsforgln,arg,pro,orleu.
<400>7
glnxaaalaglypro
15
<210>8
<211>5
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>peptide
<222>()..()
<223>基序
<400>8
glutyraspproglu
15
<210>9
<211>6
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>peptide
<222>()..()
<223>基序
<220>
<221>misc_feature
<222>(2)..(2)
<223>x为f或y
<220>
<221>misc_feature
<222>(3)..(4)
<223>x为任意氨基酸
<220>
<221>unsure
<222>(2)..(2)
<223>the'xaa'atlocation2standsforgln,arg,pro,orleu.
<220>
<221>unsure
<222>(3)..(3)
<223>the'xaa'atlocation3standsforgln,arg,pro,orleu.
<220>
<221>unsure
<222>(4)..(4)
<223>the'xaa'atlocation4standsforgln,arg,pro,orleu.
<400>9
glyxaaxaaxaaalaala
15
<210>10
<211>4
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>peptide
<222>()..()
<223>基序
<220>
<221>misc_feature
<222>(3)..(3)
<223>x为任意氨基酸
<220>
<221>unsure
<222>(3)..(3)
<223>the'xaa'atlocation3standsforgln,arg,pro,orleu.
<400>10
alaargxaaphe
1
<210>11
<211>4
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>peptide
<222>()..()
<223>基序
<400>11
glnleuphepro
1
- 还没有人留言评论。精彩留言会获得点赞!