单链VH-L1-Cκ-L2-CH1-抗体的制作方法
本发明涉及单链vh抗体(scvhab)构建体、包含这种构建体的纯重链抗体、以及在体外和体内产生该抗体的方法。
背景技术:
:抗体已成为重要的生物药物,因为它们(i)表现出可靶向多种分子形式的抗原的优异结合特性;(ii)是具有理想药代动力学的生理分子,从而使它们在受治疗的人和动物中具有良好的耐受性;和(iii)与可自然抵御致病因子的强大免疫学特性有关联。此外,存在用于从实验动物中快速分离抗体的成熟技术,该技术可以容易地针对不天然存在于体内的几乎任何异物发生特异性抗体反应。在其最基本的形式中,抗体是由两条相同的重(h)链组成,各条重链与相同的轻(l)链配对。h链和l链两者的n末端均由可变结构域(分别是vh和vl)组成,它们共同地提供具有独特抗原结合特异性的配对的h-l链。在种系dna中,不存在编码抗体vh和vl结构域的外显子。相反,各vh外显子通过h链基因座(igh)中存在的随机选择的v、d和j基因片段的重组而产生;同样地,单独的vl外显子是由在轻链基因座(igl)中随机选择的v和j基因片段的染色体重排所产生(参见图1中的小鼠igh基因座、igiκ基因座(igk或igκ)和igλ基因座(igl或igλ)(tonegawa,nature,302:575,1983;bassing,etal.,cell,109suppl:s45,2002)。小鼠基因组包含可以表达h链的两个等位基因(来自各亲本的一个等位基因)、可以表达κ(κ)l链的两个等位基因和可以表达λ(λ)l链的两个等位基因。在h链基因座处有多个v、d和j基因片段,并且在两个l链基因座处有多个v和j基因。各免疫球蛋白(ig)基因座的j基因的下游存在一个或多个编码抗体恒定区(c)的外显子。在重链基因座中,也存在用于表达不同抗体类型(同种型)的外显子。在小鼠中,被编码的同种型是igm、igd、igg1、igg2a/c、igg2b、igg3、ige和iga;在人类中,它们是igm、igd、igg1、igg2、igg3、igg4、ige、iga1和iga2。在b细胞发育期间,基因重排首先发生在含有h链v、d和j基因片段的两个同源染色体中的一个上。在前b细胞中,所得的vh外显子然后以rna水平被剪接到编码μh链的恒定区的外显子。大部分的由前b细胞合成的μh链保留在内质网(er)中,最终由于部分未折叠的ch1结构域与常居er分子伴侣bip之间的非共价相互作用而降解(haasandwabl,nature,306:387-9,1983;bole,etal.,jcellbiol.102:1558,1986)。然而,一小部分的μ链与取代轻链复合物有关联,该取代轻链复合物由不变λ5和vpreb蛋白组成,取代了bip并允许μh链/λ5/vpreb复合物与igα/β信号分子共同地作为前b细胞受体(prebcr)离开er,并经由分泌途径到达质膜(etal.,curr.top.microbiol.immunol393:3,2016)。随后,在一个时间在一个l链等位基因上发生vj重排,直到产生功能性l链,此后l链多肽可以完全置换bip并与μh链缔合,以形成用于抗原的全功能b细胞受体(bcr)。阻止细胞表面表达或不完全组装ig分子的分泌的er质量控制机制是非常严格的,因此诸如hl、hhl或hh的分子通常保留在er中,并且如果不通过组装成完整的h2l2结构而获救则被降解(该系统主要集中于保留的igh链上;因此,经常会分泌游离的l链)。然而,数十年来已知游离的单克隆h链可以在罕见的b细胞增殖性疾病(称为重链疾病(hcd))中被分泌(franklinetal.,am.j.med.,37:332,1964)。hcd中的h链被截短,随后的结构研究表明ch1结构域几乎始终被删除(corcosetal.,blood,117:6991,2011)。从机理上讲,ch1缺失使h链不发生其与bip的抑制性相互作用,因此允许其分泌,并且还阻止与l链的二硫键介导的共价缔合,因此hcd蛋白是hh二聚体。纯重链的abs(hcabs)也可以在非疾病环境中被发现。i)正常骆驼中大约75%的血清igg由hcab组成,其缺少ch1结构域并且还具有结构改变的vh结构域,由此防止与vl结构域的有效缔合(delosrios,etal.,cur.opin.struct.biol.,33:27,2015)。ii)其中κ链和l链基因位点均被灭活的小鼠仍然产生血清igg,但该抗体的产生需要类别转换重组(csr)中的错误,这导致b细胞dna中编码ch1结构域的外显子的缺失(zou,etal.,j.exp.med.,204:3271,2007)。hcab作为治疗剂是具有吸引力的,因为它们是高度稳定的,并且比常规的免疫球蛋白小。不受vl抗原结合部分阻碍的分子的vh抗原结合部分可以识别在蛋白质结构的口袋内的表位,其包括否则不可接近常规abs的病毒和g偶联蛋白受体上的酶活性位点和表位。来源于例如其中内源性vh基因已被骆驼vh基因取代且编码ch1的外显子已被删除的小鼠的基于骆驼的hcab是这种抗体的潜在来源。然而,它们的缺点是骆驼vh结构域在它们可能被用作治疗剂的人类和其他动物中具有免疫原性。其中内源性vh基因已被它们的人类对应物取代并结合κ和λl链基因座的失活的小鼠可能是hcab的来源。然而,这种抗体的产生依赖于免疫应答期间csr中相对较少的错误,因此不是高效的。wo2014/141192a2公开了纯重链抗体的产生和产生该抗体的转基因非人类动物。这种抗体缺少ch1结构域。us8754287b2公开了产生缺少ch1结构域的重链抗体的小鼠、以及包含种系修饰以删除编码ch1结构域的核酸的转基因小鼠。schusser等人(eur.j.immunol.,46:2137,2016)描述了在vjcl基因敲除鸡中没有相关轻链的纯重链抗体的表达。klein等人(biochemistry,18:1473,1979)描述了轻链的分离的可变结构域和恒定结构域与免疫球蛋白g的fd’片段的相互作用。需要生产用于诊断和治疗用途的hcab抗体的高效且有成本效益的方法。更具体地,需要能够产生抗原特异性hcab的小型、快速繁殖的动物。这种动物可用于产生能够大规模生产纯h链单克隆抗体的杂交瘤。根据前述的目的,提供了能够产生hcab的转基因非人类动物。技术实现要素:所提供的本
发明内容以简化的形式介绍了概念的选择,在下面的详细说明中将对这些概念做进一步描述。本
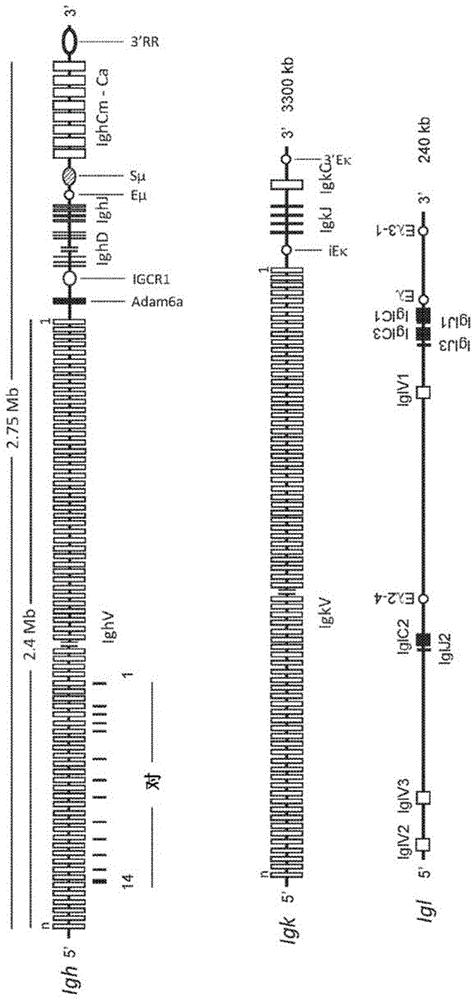
发明内容并非旨在标示要求保护的主题的关键或必要特征,也并非旨在限制要求保护的主题的范围。基于下面的书面详细说明,包括在附图中示出且在所附权利要求中定义的各方面,要求保护的主题的其他特征、细节、应用和优点将变得显而易见。本发明的目的是提供在转基因动物或体外细胞培养物中可以容易地产生的抗体构建体。该目的是由本发明权利要求的主题所解决,并且如本文的进一步描述。根据本发明,提供一种单链vh抗体(scvhab),其按照从n末端到c末端:vh-l1-cl-l2-ch1的顺序包含由vh结构域及免疫球蛋白恒定结构域cl和ch1所组成的抗原结合部分,其中l1和l2各自独立地是肽接头;并且其中cl通过β片层接触与ch1配对,从而获得cl/ch1二聚体。具体地,cl是cκ或者cλ,优选地其中cλ选自由cλ1、cλ2和cλ3组成的组,尤其包含人序列,例如图16中所示。根据进一步的具体实施方案,cλ选自由cλ6和cλ7组成的组,尤其包含人序列,例如图16中所示。当在小鼠中产生抗体时,通常h和l链的恒定区具有小鼠起源(例如包含ch1,以及cλ1、cλ2或cλ3序列中的任一序列)以便更好地与小鼠免疫系统相互作用。在分离各自的小鼠抗体后,可以容易地将c区人源化例如,包含人ch1和/或人cλ1、cλ2、cλ3、cλ6或cλ7序列中的任一序列)。具体地,cl/ch1二聚体通过抗体结构域的缔合而形成,例如形成一对结构域。具体地,cl结构域与ch1结构域的缔合通过共价键,例如通过利用接头l2,经由c-n键将cl结构域的c端连接到ch1结构域的n端。具体地,另外通过氨基酸侧链的相互作用(例如通过β折叠的β链之间的连接界面)使cl/ch1二聚体的构象稳定化,并且任选地通过一个或多个二硫键而彼此连接。具体地,cl/ch1二聚体包含至少一个域间二硫键。具体地,通过还原每个cl和ch1结构域的最c末端半胱氨酸而形成至少一个域间二硫键,从而能够形成二硫键。具体地,vh结构域的c末端通过c-n键共价连接到cl结构域的n末端,任选地利用接头l1为该区域提供增加的灵活性。具体地,利用至少一个域间二硫键,优选地将cκ(uniprotkb-p01837)中的cys107、或cλ1(uniprotkb-a0a0g2je99)中的cys105、或cλ2(uniprotkb-p01844)中的cys103、或cλ3(uniprotkb-p01845)中的cys103连接到相关ch1(uniprotkb-p01868)中的cys102的二硫桥,使cl/ch1结构域对进一步稳定化。使scvhab中的cl/ch1对稳定化的任何的这种域间二硫桥在本文中被理解为域间、链内二硫桥。通过将新的cys残基或任何其他硫醇形成氨基酸或氨基酸类似物导入位置从而在氧化反应时形成另外的s-s桥,可以对另外的二硫桥进行工程化。scvhab具体地以按类似于fab片段的方式进行布置的vh、cl和ch1抗体结构域为特征,不同之处在于将vh连接到cl结构域而不是ch1结构域,并且ch1和cl结构域另外地通过单链接头连接。因此,cl结构域是抗体重链的一部分。scvhab具体地以缺少任何轻链,尤其是没有vl结构域为特征。将示例性scvhab的结构示于图2a中。接头可以包含:酸性接头、碱性接头和结构基序、或者它们的组合。根据一具体方面,l1和l2中的任一个或两者均是人工肽,优选地是富含甘氨酸和/或丝氨酸的接头。根据另一具体方面,l1和l2的中任一个或两者均包含天然抗体序列(尤其是抗体结构域或铰链区的氨基或羧基末端序列)的一部分或由其组成。具体地,l1是具有长度为3-40个氨基酸的氨基酸序列的肽接头,优选地由下列所组成:a)任意组合的甘氨酸和/或丝氨酸的序列;或者b)vh框架序列。根据具体实施方案,scvhab不包含接头l1。这种scvhab包含直接键合或共价连接到cl结构域的vh结构域(即,vh-cl-l2-ch1)。具体地,l1包含多个甘氨酸和丝氨酸残基,或者至少由3、4、5、6、7、8、9、10、多达40个连续氨基酸中的任一个、或包含代替氨基酸的相同长度的肽组成。具体地,l2是用作连接cl/ch1结构域对的系链的肽接头。示例性的l2接头由长度为20至250、优选地40至225、或60至225个氨基酸的氨基酸序列组成,优选地由任意组合的甘氨酸和/或丝氨酸和/或精氨酸的序列组成。示例性的l2接头以序列的重复为特征,例如(ggaggaggggggtcc[seqidno:11])n,其中n=4-16。在某些情况下,l2是长度为15-90、具体地20-80、更具体地25-50或25-40个氨基酸的氨基酸序列,优选地由任意组合的甘氨酸和/或丝氨酸的序列组成。具体地,l2包含多个甘氨酸和丝氨酸残基,或者由至少20、25、26、27、28、29、30、31、32、33、34、35、40、多达80个连续氨基酸中的任一个、或者包含代替氨基酸的相同长度的肽组成。具体地,本文中所描述的肽接头包含富含丝氨酸和甘氨酸的氨基酸序列或者由它们组成,优选地其中每个氨基酸是丝氨酸或甘氨酸,更优选地其中肽接头由丝氨酸和甘氨酸残基的重复序列组成,例如(glyglyglyglyser[seqidno:35])n,其中n=2-16。通常,使用6-10个重复序列使以支持结构域的β折叠区域中的接触表面的方式而配对的cl/ch1结构域的结构稳定化。例如,任何的以下序列均适合用作肽接头l1:(glyglyglyglyser[seqidno:35])n,其中n=1-8。例如,任何的以下序列均适合用作肽接头l2:(glyglyglyglyser[seqidno:35])n,其中n=4-16。具体地,vh包含亲和力成熟的抗原结合位点或vh-cdr序列的天然构象。抗原结合部分具体地包含vh结构域的三个cdr环,即vh-cdr1、vh-cdr2和vh-cdr3或者由它们组成。抗原结合部分可通过改变一个或多个cdr环而亲和力成熟,从而优化或提高与靶抗原结合的亲和力。可以通过在任何或每个cdr序列中的一个或多个点突变(例如1、2、3或更多的点突变)而获得这种改变,以获得亲和力成熟的抗原结合位点,例如通过体内过程或者通过体外诱变技术。由转基因非人类动物所产生的抗体通常被理解为自然或天然抗体。这种自然抗体可以来源于天然抗体库或者在体内经历亲和力成熟从而获得高亲和力抗体,这些高亲和力抗体与特异性靶抗原结合,例如kd小于10-7m,例如在10-7与10-10m之间。通过体外诱变方法(例如,采用随机诱变和/或文库技术)所产生的亲和力成熟抗体可以导致甚至更高的亲和力,例如kd小于10-8m,例如小于10-11m。天然抗体有利地以vh-cdr序列的天然构象为特征。这种天然构象以抗原结合位点的天然存在的一级结构、和/或全长vh结构域的天然存在的一级结构为特征。vh结构域的天然构象可以在体内产生,例如在使亲本vh结构域的cdr序列发生突变时产生,或者通过使用人工抗体展示系统及含有人工抗体序列的各个文库产生亲本vh结构域的变体而产生,可以对这些文库进行选择以产生合适的抗体。根据一具体方面,scvhab的抗原结合部分的c末端融合到具有或不具有铰链区的其他免疫球蛋白恒定结构域。具体地,其他免疫球蛋白结构域按照从n末端到c末端的顺序至少包含ch2-ch3。具体地,单链构建体按照从n末端到c末端的顺序包含由vh-l1-cl-l2-ch1-铰链-ch2-ch3组成的延伸scvhab。本发明进一步提供包含两个延伸scvhab的重链抗体(hcab),其中第一延伸scvhab的ch2-ch3结构域与第二延伸scvhab的ch2-ch3结构域配对,从而获得fc区。本文中所描述的fc区具体地包含除第一恒定区免疫球蛋白结构域以外的抗体恒定区。因此,“fc区”是指iga、igd和igg的最后两个恒定区免疫球蛋白结构域,和这些结构域的柔性铰链n末端,以及ige和igm的最后三个恒定区免疫球蛋白结构域。就igg而言,fc包含免疫球蛋白结构域ch2和ch3(cγ2和cγ3)以及在ch1(cγ1)与ch2(cγ2)之间的铰链。fc区也可包含采用各个天然存在抗体结构域的人工变体形式的ch2或ch3结构域,例如,与所述天然存在抗体结构域具有至少90%的序列一致性。尤其是,本文中所描述的fc区包含ch2和ch3结构域的二聚体或者由其组成,这些结构域是抗体重链(hc)的一部分,其中第一hc的ch2结构域与第二hc的ch2配对,并且第一hc的ch3结构域与第二hc的ch3配对。这种二聚体可以是同源二聚体(即,由相同氨基酸序列的两个ch2-ch3结构域链组成)、或者异源二聚体(即,由两个ch2-ch3结构域链组成,其中各结构域链具有不同的氨基酸序列,例如具有不同的ch3氨基酸序列,以使fc稳定化)。具体地,铰链区来源于将ch1结构域的c末端连接到ch2结构域的n末端的抗体重链铰链区。可替代地,可以使用大约相同长度的任何其他天然或人工接头。合适的铰链区是自然igg或iga重链铰链区(seqidno:22-33)、或者具有相同长度+/-1或2个氨基酸的其功能变体,这些区域任选含有一个或多个、多达5个或以下的点突变。例如,可使用来源于小鼠抗体的铰链区,例如igg1(seqidno:28)、igg2a(seqidno:29)、igg2b(seqidno:30)、igg2c(seqidno:31)、igg3(seqidno:32)或iga(seqidno:33)。可取代地,适当地使用人铰链区,例如igg1(seqidno:22)、igg2(seqidno:23)、igg3(seqidno:24)、igg4(seqidno:25)、iga1(seqidno:26)、或iga2(seqidno:27)。铰链区通常包含一个或多个半胱氨酸残基,用以在hcab中形成二硫桥。具体地,如果使用小鼠igg1重链铰链区,hcab包含在cys104、cys107和cys109(uniprotkb-p01868)处的两个铰链区之间的链间二硫桥。具体地,第一和第二延伸scvhab是由以下抗体结构域和连接序列组成的单链构建体:按照从n末端到c末端的顺序vh-l1-cl-l2-ch1-铰链-ch2-ch3。将示例性hcab的结构示于图2b中。hcab中的第一和第二延伸scvhab可以具有相同或不同的氨基酸序列。例如,第一延伸scvhab包含第一vh,第二延伸scvhab包含第二vh。第一和第二vh可包含相同或不同的抗原结合位点,从而例如特异性地识别两种不同的靶抗原。因此,hcab可以是单特异性和二价的,或者双特异性和单价的。当产生scvhab或hcab时,所选择的结构域和/或铰链区具有人或非人动物来源。例如,在转基因小鼠中,优选地使用下列中的一个或多个生产scvhab或hcab:scvhab的vh-cl-l2-ch1或者hcab的vh-cl-l2-ch1-铰链-ch2-ch3。在这些情况下,vh外显子是在前b细胞中在重链基因座处的vdj重排期间而形成,并且在单独的b细胞中将有所不同。例如,其他元件的核苷酸序列如下:cl[cκ(seqidno:10)、cλ1(seqidno:36)、cλ2(seqidno:37)或cλ3(seqidno:38)]、l2(被鉴定为seqidno:11的序列的4-16个序列的重复)、ch1(seqidno:12)、铰链(seqidno:14)、ch2(seqidno:15)和ch3(seqidno:16)。编码cl、ch1、ch2或ch3抗体结构域的核酸序列各自具有小鼠来源。众所周知,本文中所描述的抗体可以采用其他物种的一个或多个各自序列而制备,包括例如非小鼠动物或人类来源、或者它们的任意组合,例如编码各自人抗体结构域的人类核酸序列,例如包含如下氨基酸序列的抗体结构域:cl[例如包含seqidno:39的cκ、包含seqidno:40的cλ1、包含seqidno:41的cλ2、包含seqidno:42的cλ3、包含seqidno:43的cλ6或包含seqidno:44的cλ7]、包含seqidno:45的ighg1ch1、包含seqidno:46的ighg1ch2和包含seqidno:47的ighg1ch3。众所周知,人抗体结构域的任何序列只是示例性的。可取代地,可以使用各个不同等位基因的人抗体结构域的序列。作为编码抗体结构域的动物或人类来源的核苷酸序列、或者动物或人类氨基酸序列的取代方案,可使用经修饰的(人工)核苷酸序列和各自的氨基酸序列,例如包含至少80%或至少90%的序列一致性的各个序列,假如各个抗体结构域具有在本文中所描述的各个抗体结构内配对和连接的功能。根据一具体方面,以可溶形式(例如水溶性形式)以适合使用于药物制剂的浓度,提供本文中所描述的scvhab或本文中所描述的hcab。本文中具体地提供包含分离形式(例如是从血清或产生这些抗体的动物的血液部分中分离、或者从细胞培养部分中分离)的本文中所描述的scvhab或本文中所描述hcab的可溶性制剂。根据一具体方面,本发明提供用于医学用途的本文中所描述的scvhab或本文中所描述的hcab。医学用途包括人类的治疗或兽医用途。因此,本发明提供用于疾病预防或治疗的对受试者(例如人或非人哺乳动物)进行治疗的方法,该方法包含向所述受试者施用有效量的所述scvhab或hcab。根据一具体方面,本发明提供编码本文中所描述的scvhab的核酸分子。根据另一具体方面,本发明提供编码本文中所描述的hcab的核酸分子。根据一具体方面,本发明提供包含本文中所描述的scvhab或本文中所描述的hcab的抗体库,该抗体库包含多种抗体,各抗体特异性地识别相同的靶抗原。这种抗体库被理解为具有相同抗体类型或结构的抗体文库,其中各抗体的抗原结合位点不同,以便例如产生识别相同表位的亲本抗体的抗体变体,例如亲和力成熟或者经优化的抗体变体;或者特异性地识别靶抗原但识别这种靶抗原的不同表位的抗体。可以适当地对这种抗体库进行筛选,并且可以根据期望的结构或功能特性来选择单独的文库成员,以产生抗体产物。根据一具体方面,本发明提供包含本文所描述的scvhab或本文中所描述的hcab的抗体库,该抗体库包含识别不同靶抗原的多种抗体。这种抗体库通过用具有许多不同靶抗原的复杂的多组分抗原(如病毒或细菌)进行免疫而获得,每个靶抗原具有多个表位。根据具体实施方案,抗体库被理解为天然的抗体文库,也称为免疫前抗体库,其是由最近从骨髓(它们的产生部位)中刺激产生的成熟的但未接触抗原的b细胞所表达。本文中所描述的抗体库具体地以多样性为特征,包括至少102种抗体,优选至少105、106或107中的任一种,各自以不同的抗原结合位点为特征。根据一具体方面,提供本文中所描述的抗体库,其中a)编码所述抗体的基因来源于非免疫或免疫小鼠的b细胞,或者b)抗体由哺乳动物的浆细胞分泌,优选是啮齿动物来源,尤其是小鼠来源。具体地,抗体库可通过从b细胞中克隆编码抗体库的基因或者通过由多种哺乳动物浆细胞分泌抗体而获得。具体地,由哺乳动物浆细胞分泌的抗体以糖基化模式为特征,该模式是具有哺乳动物浆细胞起源的物种的特征。大部分生理抗体同种型是以h2l2的二聚体的形式分泌,但iga可以以更高阶的二聚体或三聚体(h2l2)2和(h2l2)3的形式分泌,igm可以以五聚体(h2l2)5或六聚体(h2l2)6的形式分泌。然而,值得注意的是本文中所描述的hcab不含有l链。因此,本发明进一步提供通过对表达并分泌这种抗体的哺乳动物浆细胞进行工程化而产生本文中所描述的抗体的方法、尤其是本文中所描述的抗体库。具体地,哺乳动物浆细胞具有非人类动物来源,例如哺乳动物来源、脊椎动物来源,尤其是啮齿动物(例如小鼠或大鼠);或兔;或禽类来源(例如鸡)。具体地,哺乳动物浆细胞来源于啮齿动物,优选小鼠。根据一具体方面,本发明提供免疫球蛋白重链基因座,包含a)可变重链区,其包含每个vh、dh和jh基因片段中一个或多个,b)恒定重链区,其包含编码cl和ch1结构域的恒定外显子,和c)连接区,对这些区域进行工程化,以表达本文中所描述的scvhab或本文中所描述的hcab。具体地,将这些区域定位在所述基因座内,以便将编码l1-cl部分和l2的外显子插入编码ch1结构域的外显子的5'。具体地,恒定重链区进一步包含编码ch2和ch3结构域的外显子。具体地,基因座是重组基因座,其来源于动物,但包含至少一个外源元件,例如与该基因座的调节元件不天然地相关的一个或多个外源重链区。具体地,使用表达载体,该表达载体在宿主细胞的转染时与宿主细胞基因组发生重组,并且在生产性vdj重排后,表达编码的抗体并将其插入质膜中并且/或者由宿主细胞分泌。具体地,载体包含一个或多个外源或异源调控元件,例如可操作地连接到抗体编码序列的启动子,这些调控元件与所述抗体编码序列不天然地相关。根据一具体方面,本发明提供包含本文中所描述的基因座的重组宿主细胞。根据一具体的方面,本发明提供用本文中所描述的基因座或本文中所描述的载体进行转染的宿主细胞。具体地,宿主细胞包含非功能性内源κ轻链基因座、和非功能性内源性λ轻链基因座。该轻链基因座是非功能性基因座,例如经过修饰而失去功能或完全删除。根据一具体方面,本发明提供包含本文中所描述的基因座的转基因非人动物。具体地,该转基因非人类动物是哺乳动物,例如脊椎动物,尤其是啮齿动物(例如小鼠或大鼠);或兔;或鸟类(例如鸡)。优选地,该转基因非人动物是啮齿动物,优选小鼠。具体地,该转基因非人类动物是禽类,并且该动物使用原始生殖细胞繁殖。因此,本文中所描述的方法可以进一步包含:分离包含导入的抗体编码区的原始生殖细胞,并使用所述生殖细胞繁殖含有经取代免疫球蛋白基因座的转基因非人动物。具体地,本文中所描述的转基因非人动物包含在任何内源性轻链基因座κ或λ或二者内部的功能失出突变(包括例如沉默突变、或者使某些基因座或基因失活的突变)或缺失。具体地,转基因非人类动物在它们的基因组中携带经修饰的免疫球蛋白等位基因或其他转基因。在具体实施方案中,本发明的转基因动物进一步包含人免疫球蛋白区。例如,已开发出许多方法用人免疫球蛋白序列取代内源性小鼠免疫球蛋白区,以产生用于药物发现目的的部分或完全人抗体。这种小鼠的例子包括在例如美国专利号7,145,056;7,064,244;7,041,871;6,673,986;6,596,541;6,570,061;6,162,963;6,130,364;6,091,001;6,023,010;5,593,598;5,877,397;5,874,299;5,814,318;5,789,650;5,661,016;5,612,205;和5,591,669中所描述的小鼠。在特别有利的方面,本发明的转基因动物包含嵌合的免疫球蛋白片段,如在wabl和killeen的美国公布号2013/0219535中所描述。这种转基因动物具有包含被导入的部分人免疫球蛋白区的基因组,其中被导入的区域包含基于非人脊椎动物的内源基因组的人可变区编码序列和非编码调控序列。优选地,本发明的转基因细胞和动物具有其中部分或全部的内源性免疫球蛋白区被去除的基因组。在另一有利方面,对动物的基因组含量进行修改,使得它们的b细胞能够在每个细胞中表达多于一个的功能性vh结构域,即,这些细胞产生双特异性抗体,如在wo2017035252a1中所描述。根据一具体方面,本发明提供形成转基因非人动物的方法,包含:a)提供非人动物细胞;b)提供一种或多种包含编码如本文中所描述的scvhab或hcab的外显子的载体;c)将所述一种或多种载体导入所述非人动物细胞中;d)将所述外显子整合到所述非人动物细胞的基因组中,并选择转基因细胞,其中所述外显子已在靶位点处整合到所述非人动物细胞的细胞基因组中,该靶位点是在内源性免疫球蛋白重链基因座中,即在所述内源性免疫球蛋白重链基因座中的第一ch外显子的5’;和e)利用所述转基因细胞形成包含所述转基因细胞的转基因非人动物。具体地,转基因非人动物表达如本文中所描述的scvhab或hcab。具体地,转基因非人动物仅表达重链抗体,并且/或者不表达包括vl结构域的任何抗体构建体。具体地,使用标志来指示所述外显子成功地整合到细胞基因组中。具体地,该标志是可选择的标志,其能够在宿主中表达,从而允许容易地选择含有被导入核酸或载体的宿主。可选择标志的实例包括,例如赋予对抗菌剂(例如,嘌呤霉素、潮霉素、博来霉素或氯霉素)的耐药性的蛋白质、赋予宿主细胞代谢优势(例如营养优势)的蛋白质、以及赋予细胞功能或表型优势(例如,细胞分裂)的蛋白质。具体地,以能够将核酸序列加入到真核细胞中的方式导入载体,从而将核酸编码序列插入细胞中,其中核酸序列可瞬时地存在于细胞中或者可加入到细胞中或稳定地整合到细胞的基因组(尤其是染色体)中。具体地,通过任何靶向重组的方法,例如通过同源重组或通过位点特异性重组技术,将所述外显子在靶位点处整合到所述非人动物细胞的细胞基因组中。具体地,crispr/cas9基因组编辑系统可用于靶向重组(he,etal.,nuc.acidsres.,44:e85,2016)。具体地,所述非人动物细胞是所述非人动物的胚胎干(es)细胞。在一方面,用于取代内源性免疫球蛋白基因的宿主细胞是es细胞,然后将其用于形成转基因哺乳动物。因此,本文中所描述的具体方法可进一步包含:分离包含导入的抗体编码区的es细胞,并使用所述es细胞形成含有经工程化或取代的免疫球蛋白基因座的转基因动物。根据具体实施方案,提供用于形成转基因非人动物的方法,包含:a)提供非人动物细胞并且整合重组酶介导的盒式交换(rmce)靶位点,该靶位点的侧面与位于内源性免疫球蛋白重链基因位点的第一ch外显子的5'位置的位点特异性重组酶的识别序列相接;b)提供一种或多种载体,其包含编码本文中所描述的scvhab或hcab的外显子(这些外显子的侧面与用于位点特异性重组酶的其他识别位点相接),以及选择用于将载体靶向整合到细胞基因组中的一种或多种标志,其中其他识别位点能够与所述rmce靶位点重组;c)将识别所述rcme靶位点和其他识别位点的所述一种或多种载体和位点特异性重组酶导入所述非人动物细胞中;d)将所述外显子整合到所述非人动物细胞的基因组中,并选择转基因细胞,其中所述外显子已在所述rmce靶位点被整合到所述非人动物细胞的细胞基因组中;和e)利用所述转基因细胞形成包含所述转基因细胞的转基因非人动物。具体地,用于位点特异性重组酶的任何的所述识别位点是重组酶识别位点(例如,cre/lox、flp-frt等),其中该重组酶能够在其两个识别位点之间切除dna序列。根据另一具体实施方案,提供形成转基因非人动物的方法,包含a)提供非人动物细胞,其包含内源性免疫球蛋白重链基因位点的第一ch外显子的靶位点5';b)提供一种或多种载体,其包含编码本文中所描述的scvhab或hcab的外显子(这些外显子的侧面和与所述靶位点为同源的dna序列相接),以及选择用于将载体靶向同源重组到细胞基因组中的的一种或多种标志;c)将所述一种或多种载体导入所述非人动物细胞中;d)将所述外显子整合到所述非人动物细胞的基因组中,并选择转基因细胞,其中所述外显子已在所述靶位点被整合到所述非人动物细胞的细胞基因组中;和e)利用所述转基因细胞形成包含所述转基因细胞的转基因非人动物。具体地,可使用同源靶向载体或“靶向载体”,其是包含编码靶向序列、位点特异性重组位点、和任选地可选择标志基因的核酸的载体,其用于使用宿主细胞中的同源介导重组来修饰内源性免疫球蛋白区。例如,在本发明中可以利用同源靶向载体将位点特异性重组位点导入宿主细胞基因组的特定区域中。根据一具体方面,本发明提供用于产生抗体的方法,包含:a)在非人动物中表达异源免疫球蛋白重链基因座,该基因座包含i)可变重链区,其包含每个vh、dh和jh基因片段中的一个或多个,ii)恒定重链区,其包含编码cl和ch1结构域的恒定外显子,和iii)连接区,将这些区域工程化并定位成表达本文中所描述的scvhab或hcab,其中非人动物不表达内源性κ和/或λ基因座;和b)产生抗体,其分别是所述scvhab和所述hcab或分别至少包含所述scvhab或所述hcab的vh结构域。具体地,非人动物包含本发明的基因座,并且将在本文中进一步描述。具体地,利用合适的基因靶向技术(例如采用位点特异性重组酶技术、或crispr/cas9技术的靶向同源重组)处理非人动物以加入基因座。具体地,非人动物是本发明的转基因非人动物,并且在本文中进一步描述。特别地,非人动物不表达内源性κ和/或λ基因座,因为所述内源性κ和/或λ基因座被删除或沉默,或者为了功能失去而发生突变。根据具体实施方案,该方法进一步包含用抗原免疫非人动物因而引起针对该抗原的免疫反应从而导致亲和力成熟特异性单克隆或多克隆抗体的产生的步骤。可以以任何方便的方式,在有或没有佐剂的情况下,向非人动物施用抗原,并且可以按照预定计划进行施用。在免疫后,可以对来自经免疫动物的血清或奶进行分级,以便纯化对抗原具有特异性的药用级多克隆抗体。在转基因鸟类的情况下,也可以通过将蛋黄进行分级而制作抗体。浓缩的纯化的免疫球蛋白级分可利用色谱法(亲和力、离子交换、凝胶过滤等),使用盐(如硫酸铵)、有机溶剂(如乙醇)、或聚合物(如聚乙二醇)的选择性沉淀而获得。为了制作单克隆抗体,可从经免疫转基因动物中分离出抗体产生细胞,例如脾和/或淋巴结细胞,并且用于与经转化细胞系的细胞融合以产生杂交瘤,或者通过标准分子生物学技术对编码抗体的cdna进行克隆并且在转染细胞中表达。制作单克隆抗体的方法在本领域中是完善的。具体地,该方法进一步包含制备杂交瘤以及产生并筛选抗体产生细胞(尤其是特异性识别靶抗原的细胞)的步骤。具体地,该方法进一步包含从免疫的非人动物中分离核酸序列以在细胞培养物中产生特异性抗体或其片段,特别是抗原结合片段的步骤。这种抗体或其抗原结合片段在本文中被理解为超免疫抗体。根据具体实施方案,本文中所描述的抗体是在使用合适的生产宿主细胞系的细胞培养物中生产。具体地,该生产采用细菌、酵母、植物、昆虫或哺乳动物细胞培养。具体地,这些宿主细胞是在与编码本文中所描述的抗体的各个核酸分子的重组后使用。具体地,有利的是使用任何哺乳动物宿主细胞:bhk、cho、hela、hek293、mdck、nih3t3、ns0、per.c6、sp2/0或vero细胞。根据具体方面,本发明提供本文中所描述的转基因非人动物在产生scvhab或hcab抗体或者包含vh结构域的其片段,以及任选地进一步产生包含所述vh结构域的抗体中的用途。根据具体方面,本发明提供本文中所描述的转基因非人动物在产生文库,尤其是scvhab或hcab抗体的天然文库、或者包含vh结构域的其片段、或者编码或表达所述天然文库的核酸序列的文库的用途。本文中所描述的转基因细胞可用于产生表达文库以鉴定目标抗体,例如通过从b细胞克隆编码抗体的基因,或者通过选择在浆细胞膜上表达抗体的工程化小鼠中具有明确特异性的浆细胞,例如在us20170226162a1中所描述。因此,本发明还包括利用细胞技术所产生的抗体文库,用于鉴定由浆细胞所表达的抗原特异性抗体。当产生本文中所描述的scvhab或hcab时,可以通过合适的技术来表征vh结构域或其抗原结合位点,以将任何类型的抗体工程化,例如全长抗体或其抗原结合片段,或者甚至单vh结构域抗体及包含这种单vh结构域抗体的抗体构建体。例如,可以确定vh结构域或其抗原结合位点的氨基酸序列或编码核苷酸序列,并且与抗体构建体的其他序列或并入该vh结构域或其抗原结合位点的其他结合分子发生重组。一些示例性实施方案提供本发明的转基因动物,这些转基因动物进一步包含人免疫球蛋白区。例如,已开发出许多方法,采用人免疫球蛋白序列取代内源性小鼠免疫球蛋白区,以产生用于药物发现目的的部分或完全人抗体。这种小鼠的实例包括在例如美国专利号7,145,056;7,064,244;7,041,871;6,673,986;6,596,541;6,570,061;6,162,963;6,130,364;6,091,001;6,023,010;5,593,598;5,877,397;5,874,299;5,814,318;5,789,650;5,661,016;5,612,205;和5,591,669中所描述的小鼠。一些其他示例性实施方案提供本发明的转基因动物,这些转基因动物进一步包含如wabl和killeen的美国公布号2013/0219535中所描述的嵌合免疫球蛋白片段。这种转基因动物具有包含被导入的部分人免疫球蛋白区的基因组,其中被导入的区域包含基于非人脊椎动物的内源基因组的人可变区编码序列和非编码可变序列。优选地,本发明的转基因细胞和动物具有其中部分或全部的内源性免疫球蛋白区被去除的基因组。一些其他示例性实施方案提供本发明的转基因动物,这些转基因动物进一步包含对免疫球蛋白重链基因的改变,以允许产生双特异性抗体,例如在wo2017035252a1、us20170058052a1中所描述。其他实施方案提供来源于基因修饰动物的初级b细胞、永生化b细胞、或杂交瘤。其他实施方案包括由来自基因修饰动物的工程部分的免疫球蛋白重链基因进行转录的部分或全部免疫球蛋白;以及来源于基因修饰动物细胞的部分或全部的工程免疫球蛋白。下面更详细地描述本发明的这些和其他的方面、目的及特征。附图说明图1:描述了小鼠igh基因座(上部)[包括v(ighv)、d(ighd)、j(ighj)和c(ighc)基因片段;存在多个ighc外显子用以编码不同的igh链同种型]、igk基因座(igk,中部)[包括v(igkv)、j(igkj)和c(igkc)基因片段]和igλ基因座(下部)[包括v(iglv)、j(iglj)和c(iglc)基因片段]。还示出了(1)pair元件,其是对igh环至关重要的顺式调控序列,用以确保在vdj重排中远端vh基因片段的应用;(2)adam6a雄性育性促成基因;(3)基因间控制区1(igcr1),其含有用于调控igh基因座的有序、谱系特异性重排的位点;(4)eμ、ieκ和eλ2-4、重链、κ和λ轻链内含增强子;(5)3’eκ、eλ和eλ3-1、κ和λ轻链3’增强子;(6)sμ、μ开关区域;和(7)3’调控区(3’rr),其是控制同种型转换的顺式作用元件。图2:(a)跨模(tm)和分泌形式的scvhab。tm形式由b细胞表达作为抗原受体(bcr)。成熟的蛋白质具有结构vh-l1(任选的)-cl-l2-ch1-tm;vh、重链可变区、l1、接头1、cl、κ或λ轻链恒定区、l2、接头2、ch1、重链ch1结构域。tmscvhab与igα/β(cd79a/cd79b,未示出)是相关的。分泌形式的scvhab是由浆母细胞和浆细胞所产生,除了具有71个氨基酸长度的tm区被单个赖氨酸残基取代并且该分子与igα/β无关以外其具有基本上相同的结构。(b)tm和分泌形式的hcab。tm形式由b细胞表达作为bcr。成熟的蛋白具有结构vh-l1(任选的)-cl-l2-ch1-h-ch2-ch3-tm;h,重链铰链区,ch2,重链ch2结构域,ch3,重链ch3结构域。tmhcab也与igα/β(cd79a/cd79b,未示出)相关。分泌形式的hcab由浆母细胞和浆细胞所产生,除了具有71个氨基酸长度的tm区被单个赖氨酸残基取代并且该分子与igα/β无关以外其具有基本上相同的结构。图3:基于抗体fab晶体结构的接头2(l2)长度的设计中的考虑因素。l2将cl连接到ch1。图中所示的fab结构及其组成结构域属于igg1κmab(结构生物信息学蛋白质数据库id的研究合作实验室:2xkn)。连接cκ的cooh末端与ch1的nh2末端的桥接距离为(用虚线表示),但由于cκ和ch1的相对位置,接头最好较长以便连接cooh-与nh2-末端。(ggggs[seqidno:35])4接头的理论长度为这小于两个末端之间距离的覆盖范围的两倍因此,示例性的接头长度为(ggggs[seqidno:35])4-16。图4:用于测试体外细胞表面表达的所产生的重链抗体(hcab)构建体。(1)阳性对照,常规h2l2igg(2和3),已知阳性对照在没有lc的细胞表面上被表达。(2)缺少ch1的骆驼样hcab。(3)其中vl直接地连接到vh且缺少ch1的单链fv(scfv)。(4)本文中所描述的hcab,具有蛋白质结构域结构nh-vh-cκ-l2-ch1-ch2-ch3-tm-cooh。(5)与构建体4相同,除了cl和ch1的顺序被反转,vh-ch1-l2-cκ-ch2-ch3-tm-cooh。(6)阴性对照,没有lc(h2l0)的常规igg。这里所说明的构建体编码小鼠hc(例如,小鼠igg1),其含有用于插入质膜中的跨模区。还产生编码分泌形式的构建体,用于测试hcab分泌。将这些构建体转染到hek293t细胞中。图5:图4中所描绘的hcab构建体的细胞表面表达。使用脂质体2000(lipofectamine2000)(invitrogen)连同编码myc标签人cd4(hcd4)的载体作为转染对照,将图4中所描绘的构建体瞬时地转染到hek293t细胞中。此外,所有hek293t细胞均用表达小鼠cd79a和cd79b(igα/igβ)两者的构建体进行共转染,小鼠cd79a和cd79b(igα/igβ)是b细胞抗原受体的表面表达所需的共受体,包括膜结合形式的hcab(wienandsandengles,int.rev.immunol.,20:679,2001)。在20-24小时后,为了细胞表面hcd4、小鼠igg1(migg1)和小鼠κ轻链(migκ)将细胞染色,并利用流式细胞术进行了分析。附图顶部的数字对应于图4中的构建体编号。每个流程图顶部的数字表示阴性(左)和阳性(右)细胞的频率。图6:用于细胞内hcd4、migg1和migκ的转染细胞的染色。为了hcd4、migg1和migκ的细胞内表达,对来自图5中所示的转染的细胞样品进行固定、通透化和染色。利用流式细胞术验证所有构建体均被表达。图7:(a)为了比较cκ、cλ1和cλ2的体外细胞表面表达效率而生成的重链抗体(hcab)构建体。(1)阳性对照,缺少ch1的骆驼样hcab。(2)本文中所描述的hcab,具有结构vh-cκ-l2-ch1-ch2-ch3-tm。(3)本文中所描述的hcab,具有结构vh-cλ1-l2-ch1-ch2-ch3-tm。[seqidno:47,核苷酸序列;seqidno:48,氨基酸序列]。(4)本文中所描述的hcab,具有结构vh-cλ2-l2-ch1-ch2-ch3-tm。这里所说明的构建体编码用于插入质膜的跨模区。还产生了编码分泌形式的构建体,用以测试hcab分泌。(b)构建体3融合基因,5'至3’的示意图。sp,信号肽;vh3-11、d2-21、jh4是用于重链vdj重排的vh、dh和jh基因片段;cλ1,轻链恒定区;l2,接头2;ch1,编码igg1的ch1结构域的外显子;h,igg1铰链区外显子;ch2,编码igg1的ch2结构域的外显子;ch3-s,编码igg1的ch3结构域和分泌尾的外显子;m1+m2,编码igg1跨模区的外显子。图8:图7a中所描绘的hcab构建体的细胞表面表达。使用脂质体2000(lipofectamine2000)(invitrogen)连同编码myc标签人cd4(hcd4)的载体作为转染对照,将图7a中所描绘构建体瞬时地转染到hek293t细胞中。此外,所有hek293t细胞均用表达小鼠cd79a和cd79b(igα/igβ)两者的构建体进行共转染,小鼠cd79a和cd79b(igα/igβ)是抗原受体的表面表达所需的共受体,包括膜结合形式的hcab(wienandsandengles,int.rev.immunol.,20:679,2001)。在20-24小时后,为了细胞表面hcd4(顶部行)或小鼠migg1(中间行)将细胞染色,或者为了细胞内小鼠migg1(底部行)进行固定和通透化和染色,并且利用流式细胞术进行分析。左边(无hc)的列是仅用hcd4构建体进行转染的细胞。第1-4列是用hcd4加入如图7a中编号的构建体进行转染的细胞。在各流程图顶部的数字表示阴性(左)和阳性(右)细胞的频率。图9:各种hcab构建体的分泌和细胞内表达的蛋白质印迹分析。将用所示构建体转染的细胞培养40-48小时,然后离心。收集上清液以检测hcab的分泌(左),将细胞团块溶解于np-40以检测hcab的细胞内表达(右)。在非还原条件下对样品进行sds-聚丙烯酰胺凝胶电泳(sds-page),印迹到pvdf膜上,然后用hrp标记抗小鼠iggfc抗体进行探测。载体的migg1、ch1和/或fc区编码migg1。载体的migg2a、ch1和/或fc区编码migg2a。如附图中所示,构建体的l2长度(6或10)和cl(cκ、cλ1或cλ2)也不同。空载体、无插入的表达载体。泳道1、vh-fc、骆驼样hcab。分子量标志(kda0在凝胶的左侧和右侧可见。图9表示序列ggggs(seqidno:35)的重复序列。图10:各种hcab构建体的分泌和细胞内表达的蛋白质印迹分析。除了凝胶在还原条件下操作外与图9相同。图10表示序列ggggs(seqidno:35)的重复序列。图11:图9和图10中所示印迹的转染和上样对照(loadingcontrols)。将印迹剥离再用抗myc或抗gapdhmab进行重新探测。hcd4构建体包括myc标签,用作转染对照。gapdh是持家基因,用作上样对照。图11表示序列ggggs(seqidno:35)的重复序列。图12:利用elisa对hcab分泌的定量。利用elisa测定了图9中所描绘的转染子的上清液中分泌的igg1(左)和igg2a(右)hcab的量。图13:通过同源重组将小鼠cl-l2-ch1-h-ch2-ch3_s-tm基因盒导入在ighm上游位置的内源性小鼠igh基因座以产生hcab的策略。在该图和图14和图15中,“内源性小鼠igh基因座”是在wabl和killeen的美国公布号20130219535a1中所描述的含有部分人igh基因座的es细胞中。(a,b)是靶向载体的结构。在附图中被标记为a.和b.并且用虚线连接的片段在靶向载体中是连续的。(c)是待靶向的内源性小鼠igh基因座的区域。(d,e)是所得到的靶向小鼠igh基因座。在附图中被标记为d.和e.并且用虚线连接的片段在体内是连续的。(f,g)是用flp重组酶去除可选择标记后的最终靶向基因座。在附图中被标记为f.和g.并且用虚线连接的片段在体内是连续的。此图及后续的附图中的igcl表示cκ或者cλ(在小鼠的情况下cλ1、cλ2或cλ3)的轻链恒定区。图14和15:通过重组酶介导的盒式交换(rmce)将小鼠cl-l2-ch1-h-ch2-ch3_s-tm基因盒导入在ighm上游位置的内源性小鼠igh基因座中以生产hcab的策略。图14:步骤1,rmce受体等位基因的生成。(a)是rmce靶向载体的结构。(b)是要靶向的内源性小鼠igh基因座的区域。(c)是所得到的靶向小鼠igh基因座。图15:步骤2,用cl-l2-ch1-h-ch2-ch3_s-tm载体靶向rmce修饰的受体等位基因。(a)是rmce靶向载体的结构。(b)是rmce修饰的igh基因座。(c,d)是所得到的靶向igh基因座。在附图中被标记为c.和d.并且用虚线连接的片段在体内是连续的。图16:本文中所引用的序列。具体实施方式除非另外指出或定义,本文中使用的所有术语具有其在本领域中的通常含义,这对于本领域技术人员将是显而易见的。例如参考标准手册,如sambrooketal,“molecularcloning:alaboratorymanual”(2nded.),vols.1-3,coldspringharborlaboratorypress(1989);lewin,"genesiv",oxforduniversitypress,newyork(1990),和janewayetal,“immunobiology”(5thed.,ormorerecenteditions),garlandscience,newyork,2001。如本文中所引述的氨基酸残基在抗体中的位置应理解为对应于imgt编号的位置。(国际免疫遗传学信息)。imgt编号是指天然存在抗体的编号。关于imgt数据库和编号方案的解释可以参见giudicellietal.,nuc.acidsres.,34:d781,2006。本文中被称为scvhab和hcab的抗体构建体是非天然存在的人工构建体。众所周知,本发明的材料、方法和用途(例如,特定地指代分离的核酸序列、氨基酸序列、表达构建体、转化的宿主细胞、转基因动物和重组抗体)是“人造的”或合成的,因此不被视为“自然规律”的结果。本文中所使用的术语“抗体”应指代由采用各种组合或构造的抗体结构域所组成或包含这些抗体结构域的多肽或蛋白质,其被理解为免疫球蛋白的重链和/或轻链的恒定和/或可变结构域,具有或没有接头序列。多肽被理解为抗体结构域,如果包含β-桶状结构则由通过环序列连接的抗体结构域结构的至少两个β链组成。抗体结构域可具有天然结构或者通过诱变或衍生化进行修饰,例如修改抗原结合特性或任何其他特性,例如稳定性或功能特性,例如与fc受体,fcμ、fcα/μ、fcα、fcε和/或fcγ受体(例如,小鼠中的fcrn、fcγri、fcγriib、fcγriii或fcγriv)或者多聚ig受体(pigr)结合。在本文,术语“抗体”和“免疫球蛋白”可互换地使用。如本文所使用的术语“抗体”应尤其指包含vh作为单一可变抗体结构域连同具有一个或多个连接序列或铰链区(例如重链)的恒定抗体结构域的抗体构建体,其由一条或两条单链组成,其中各单链包含连接到恒定结构域的可变重链区(或vh)或者由其组成。示例性的抗体包含本文中进一步描述的任何scvhab或hcab或者由其组成。本文中所描述的抗体可包含属于igg类型(例如,igg1、igg2、igg3或igg4亚型)、iga1、iga2、igd、ige或igm类型的抗体结构域,或它们的鼠类对应物igg1、igg2a/c、igg2b、igg3、iga、igd、ige或igm或者由它们所组成。据此,抗体通常被理解为包括基本上由免疫球蛋白基因或免疫球蛋白基因的片段所编码的一种或多种多肽的蛋白质(或蛋白质复合物)。公认的免疫球蛋白基因包括κ、λ、α、γ、δ、ε和μ恒定区基因,以及免疫球蛋白可变区基因。轻链(lc)被分类为κ或者λ。重链(hc)被分类为γ、μ、α、δ或ε,它们相应地分别定义免疫球蛋白类别igg、igm、iga、igd和ige。在典型的igg抗体结构中,hc或lc各自含有彼此连接以产生一对结合位点结构域的至少两个结构域。在特定的情况下,重链可包含lc恒定域,但仍被认为是hc,例如缺乏轻链可变结构域或区域。抗体的hc可包含将抗体的一个或两个抗原结合臂连接到fc部分的铰链区。尤其是,本文中所描述的scvhab可适当地包含铰链区作为c-末端延伸,例如用以将scvhab连接到包含肽/多肽序列的其他元件。示例性的抗体构建体可含有抗体恒定结构域,例如通过铰链区连接的fc的恒定结构域。铰链区可以是免疫球蛋白的天然存在的重链铰链区,例如igg1或igg3的铰链区,或者是人工铰链区,该人工铰链区包含作为天然存在氨基酸的多个连续氨基酸或由其所组成,这些氨基酸具有大致相同的长度(+/-20%、或+/-10%)。优选的铰链区包含一个或多个(例如2、3或4个)半胱氨酸残基,这些半胱氨酸残基可与另一个铰链区形成二硫桥键,从而获得二聚体构建体。本文中所描述的抗体可包含一个或多个缩短或者延伸的抗体结构域,例如使用连接序列或接头。这种连接具体地是通过重组融合或化学键合。特异性连接可通过将一个结构域的c末端连接到另一个结构域的n末端而实现,例如其中将末端区中的一个或多个氨基酸残基删除以缩短结构域大小、或者延伸以增加结构域的灵活性。具体地,缩短的结构域序列包含c末端和/或n末端区域的删除,例如删除至少1、2、3、4或5、多达6、7、8、9或10个氨基酸。利用接头的结构域延伸可通过来源于免疫球蛋白结构域的n-或c-末端区域的氨基酸序列而实现,其将会天然地位于该结构域附近,例如包含结构域之间的天然衔接点。可取代地,接头可含有来源于铰链区的氨基酸序列。然而,接头也可以是人工序列,例如富含多个gly和ser氨基酸或者由它们组成。术语“抗原结合位点”或“结合位点”是指参与抗原结合的抗体的部分。天然抗体中的抗原结合位点是由重链(“h”)和/或轻链(“l”)的n-末端可变区(“v”)或其可变结构域的氨基酸残基所构成。重链(和任选的轻链)的v区内的三个高度可变延伸(称为“高变区”)介于被称为骨架区的更保守的侧翼延伸之间。抗原结合位点提供与结合的表位或抗原的三维表面互补的表面,并且高变区被称为“互补决定区”或“cdr”。包含于cdr中的结合位点在本文中也称为“cdr结合位点”。术语“cdr区”或各自的序列是指可变抗体结构域的可变抗原结合区,如vh或vhh结构域,其包括能够与抗原发生结合相互作用的变化结构。具有cdr区的抗体结构域可以原样使用或整合在较大的蛋白质构建体中,从而形成具有结合功能的这种构建体的特定区域。变化结构可以来源于结合蛋白(如免疫球蛋白)的天然库,特别是来源于抗体或免疫球蛋白样分子。变化结构也可以通过随机化技术而产生,特别是本文中所描述的。这些变化结构包括抗体可变结构域的诱变cdr环区域,特别是免疫球蛋白的cdr环。通常,具有带特异性cdr结构的抗原结合位点的抗体能够特异性结合靶抗原,即,通过一对vh/vl结构域的cdr环特异性地识别这种靶抗原。在hc抗体中,抗原结合位点以仅由vh-cdr1、vh-cdr2和vh-cdr3环组成的特异性cdr结构为特征。如果由动物(例如本文中所描述的转基因非人动物)产生,这种抗原结合位点应理解为是天然的,或者具有天然结构和/或构象。尽管抗原结合位点可以人工产生,但由于利用合成新结构的重组技术进行工程化,因而将编码各个抗体的各个基因加入转基因非人动物中导致具有天然构象的新合成抗体的产生。可以利用任何体内或体外亲和力成熟技术使这种天然构象进一步亲和力成熟,从而产生包含以天然构象为特征并且进而以特异性结合其靶抗原的高亲和力为特征的人工抗原结合位点的多克隆和/或单克隆抗体。术语“抗体”应适用于动物来源的抗体,例如哺乳动物,包括人、鼠、兔和大鼠,或禽类(例如鸡),该术语应尤其包括基于动物来源序列(例如小鼠序列)的重组抗体。术语“抗体”还适用于完全人抗体。关于免疫球蛋白所使用的术语“完全人”应理解为包括具有来源于人种系免疫球蛋白序列的可变区和恒定区的抗体。人抗体可包含不是由人种系免疫球蛋白序列编码的氨基酸残基(例如,通过体外随机或位点特异性诱变或者通过体内体细胞突变而导入的突变),例如在cdr中。人抗体包括从人免疫球蛋白或抗体文库中分离或者从一种或多种人免疫球蛋白转基因动物中分离的抗体。人免疫球蛋白优选地选自或来源于由lga1、lga2、igd、ige、lgg1、lgg2、lgg3、lgg4和igm组成的组。鼠免疫球蛋白优选地选自或来源于由iga、igd、ige、lgg1、lgg2a、lgg2b、lgg2c、lgg3和igm组成的组。术语“抗体”还适用于嵌合抗体,其具有来源于不同物种的混合序列,如鼠源和人源的序列。具体地,术语“抗体”适用于由转基因非人动物(例如小鼠)产生的抗体,其包含人抗原结合区和非人(例如,鼠)恒定区或骨架序列。关于免疫球蛋白或抗体所使用的术语“嵌合”是指那些分子,其中抗体链的一部分与来源于特定物种或属于特定类别的免疫球蛋白中的相应序列为同源,而该链的其余部分与另一物种或类别中的相应序列为同源。通常,可变区模仿来源于一个哺乳动物物种的免疫球蛋白的可变区,而恒定部分与来源于另一种免疫球蛋白的序列为同源。在一个实例中,可变区可以来源于目前已知的来源,这些已知来源使用来自人宿主生物体的易于获得的b细胞连同来源于例如非人细胞制品的恒定区。术语“抗体”还适用于单克隆抗体,尤其是重组抗体,该术语包括通过重组方法制备、表达、产生或分离的所有类型的抗体和抗体结构,例如来源于动物(例如哺乳动物,包括人)的抗体,其包括含自不同来源的基因或序列,例如嵌合、人源化抗体、或由杂交瘤获得的抗体。其他的例子是指从经转化而表达抗体的宿主细胞中分离的抗体,或者从抗体或抗体结构域的重组组合文库中分离的抗体,或者通过涉及到将抗体基因序列剪接到其他dna序列的任何其他方式所制备、表达、产生或分离的抗体。术语“抗体”应理解为包括新的或现有的(在本文中称为“亲本”)分子(例如天然存在的免疫球蛋白)的功能活性变体。还应理解的是,该术语包括抗体变体,并且也应包括这种分子的衍生物。衍生物是一种或多种抗体和/或融合蛋白的任意组合,其中抗体的任何结构域(例如,包含vh结构域或vh结构域的抗原结合位点的抗体结构域)可在任何位置融合到一种或多种其他蛋白质(如其他抗体,例如包含cdr环、受体多肽的结合结构),但也可融合到其他的配体、酶、毒素等。本文中所描述的抗体可特异性地用作分离多肽或者用作组合分子,例如,通过与其他肽或多肽的重组、融合或偶联技术。抗体的衍生物也可通过各种化学技术(如共价偶联、静电相互作用、二硫键等)与其他物质缔合或结合而获得。与抗体结合的其他物质可以是脂质、碳水化合物、核酸、有机和无机分子或者它们的任意组合(例如,peg、前药或药物)。衍生物还包含具有相同氨基酸序列但完全地或部分地由非天然或化学修饰的氨基酸所制作的抗体。在一具体实施方案中,抗体是包含允许与生物学上可接受的化合物发生特异性相互作用的另外的标签的衍生物。对于可用于本发明的标签没有特定限制,只要它对免疫球蛋白与其靶向的结合没有或具有可接受的负面影响即可。合适的标签的例子包括his标签、myc标签、flag标签、strep标签、钙调蛋白标签、gst标签、mbp标签、和s标签。在另一具体实施方案中,抗体是包含标记的衍生物。本文中所使用的术语“标记”是指直接地或间接地与抗体偶联从而产生“标记”抗体的可检测的化合物或组合物。标签可由自身检测,例如放射性同位素标记或荧光标记,或者在酶标记的情况下,可催化可检测的底物化合物的化学改变。抗体的衍生物例如来源于亲本抗体或抗体序列,如亲本抗原结合(例如cdr)或骨架(fr)序列,例如在硅化或重组工程中或者通过化学衍生化或合成所获得的突变体或变体。术语“变体”应具体地包括功能活性变体。就抗原结合的特异性而言,本文中所描述的抗体的功能变体具有特别的功能。术语“变体”应特别地指代抗体,例如突变体抗体或抗体的片段,例如通过诱变方法获得的,尤其是指将插入物或缺失物删除、交换、导入特定的抗体氨基酸序列或区域中或者化学衍生化氨基酸序列,例如在恒定结构域中,以工程化提高的抗体稳定性、增强的效应子功能或半衰期,或者在可变结构域中以调节抗原结合特性,例如通过本领域可获得的亲和力成熟技术。可采用任何的已知诱变方法,包括在期望位置的点突变,例如通过随机技术获得的突变,或者用于scvhab或hcab工程化的结构域缺失。在某些情况下,例如用任何可能的氨基酸或优选氨基酸的选择而随机地选择位置,以使抗体序列随机化。术语“诱变”是指用于改变多核苷酸或多肽序列的任何本领域公认的技术。优选的诱变类型包括易错pcr诱变、饱和诱变、或其他的位点专一诱变。当在细胞表面上或在细胞内表达抗原时,抗体在抗原结合方面的功能活性通常是在elisa测定、biacore测定、octetbli测定或基于流式细胞术的测定中进行测定。可获得功能活性变体,例如通过改变亲本抗体的序列,例如具有免疫球蛋白的特异性天然结构(如igg1结构)的单克隆抗体,以获得在识别靶抗原时具有相同的特异性但具有与母体结构不同的结构的变体,例如用以修饰任何抗体结构域,以便导入特定的突变或者产生母体分子的片段。特定的功能性活性变体包含一种或多种功能活性cdr变体或亲本抗体,它们各自在亲本cdr序列中包含至少一个点突变,并且包含与亲本cdr序列具有至少60%序列一致性,优选地至少70%、至少80%、至少90%的序列一致性的氨基酸序列或者由其组成。特异性变体是例如亲本抗体的功能活性变体,其中将亲本cdr序列加入到人骨架序列中,其中每个亲本cdr序列的任选地1、2、3或4个氨基酸残基可进一步通过导入点突变而突变,以提高亲本或人源化抗体的稳定性、特异性和亲和力。具体地,该抗体可以包含亲本抗体的任何cdr序列的功能活性cdr变体,其中该功能活性cdr变体包含下列中的至少一种:a)亲本cdr序列中的1、2或3个点突变,优选地其中每个cdr序列中的点突变的数目为0、1、2或3;和/或b)亲本cdr序列的四个c末端或四个n末端或四个中心氨基酸位置的任一个中的1或2个点突变;和/或c)与亲本cdr序列具有至少60%的序列一致性;优选地其中功能性活性变体抗体包含本文中所描述的功能活性cdr变体中的至少一种。具体地,包含一种或多种功能活性cdr变体的功能活性变体抗体具有与亲本抗体相同表位结合的特异性。根据一具体方面,点突变是一个或多个氨基酸的氨基酸置换、缺失和/或插入中的任一种。相对于抗体序列的“氨基酸序列一致性百分比(%)”被定义为在候选序列中与特定多肽序列中的氨基酸残基相同的氨基酸残基的百分比,在根据本领域众所周知的方法比对序列并导入缺口之后,如clustalw(chennaetetal.,nucleicacidsres.,31:3497,2003),如果需要,以获得最大序列一致性百分比,并且不考虑将任何保守取代作为序列一致性的一部分。本领域技术人员可以确定用于测量比对的适当参数,包括在进行比较的序列的全长上实现最大比对所需的任何算法。抗体变体具体地被理解为包括具有特定糖基化模式的同源物、类似物、片段、修饰或变体,例如通过糖基化工程所产生的,其是功能性的并且可以用作功能等同物,例如结合到特定靶向并且具有不同的功能性质。抗体可以是糖基化的或非糖基化的。例如,如本文中所描述的重组抗体可在合适的哺乳动物细胞中被表达以允许由表达抗体的宿主细胞所确定的分子的特异性糖基化,或者在缺少糖基化机制的原核细胞中被表达,从而得到未糖基化的蛋白质。术语抗体结构域(尤其是恒定抗体结构域)的“β折叠”或“β链”,在本文中以下列方式理解。抗体结构域通常由至少两条β链组成,这些β链经由至少两个或三个主链氢键而侧向地连接,从而形成通常被扭曲的折叠片。β链是通常具有3至10个氨基酸长度的单个连续氨基酸延伸,从而采用这种延伸构象并且包括与至少一个其他链结合的主链氢键,使得它们形成β片层。在该β片层中,大部分的β链与其他链相邻地排列,并且与它们的邻域形成广泛的氢键网络,其中一条链骨架中的nh基团与相邻链的骨架中的c=o基团形成氢键。抗体恒定结构域(如cl(cκ,cλ,)、ch1、ch2或ch3结构域)的结构类似于可变结构域的结构,由通过环连接的β-链所组成,其中部分的环含有短α-螺旋状延伸。如由各种fc晶体结构的x射线晶体学b因子可看到的,骨架大多是刚性的并且环相对地更加柔性。抗体恒定结构域通常具有形成β片层(a-b-c-d-e-f-g)的七条β链,其中β链通过环连接,三个环位于结构域(a-b、c-d、e-f)的n末端顶端,另外三个环位于结构域(b-c、d-e、f-g)的n末端顶端。结构域的“环区”是指位于β链的区域之间的蛋白质部分(例如,各ch3结构域包含从n-末端向c-末端取向的7个β片层,a至g)。具体地,通过连接与a、b和e链有关联的结合表面而产生一对抗体结构域,如恒定抗体结构域,例如本文中所描述的在scvhab或hcab中所包含的cl/ch1结构域对,或者fc的任何抗体结构域对(在本文中也称为第一抗体结构域的β片层区域),使第一抗体结构域与第二结构域的β片层区域接触(即,配对)以产生二聚体。在某些实施方案中,抗体结构域可以包含野生型氨基酸序列,例如来源于动物(包括人类)的野生型氨基酸序列、或者包含突变的人工氨基酸序列,例如可以用异源序列取代一条或多条β链的至少一部分,例如包含促进与另一个结构域配对的突变(例如域间二硫桥,如连接两个抗体结构域的β片层区域)、结和/或孔突变、或链交换。特定的结构域突变可以包括新(额外的)氨基酸残基(例如cys残基)的加入,这能够形成额外的域间或链间二硫桥以稳定化a)通过额外的域内二硫键形成抗体结构域,和/或b)例如在本文进一步描述的抗体构建体中,通过cl结构域和ch1结构域之间的结构域间二硫桥形成结构域对;和/或c)通过另外的链间二硫键桥接形成抗体结构域的两条链。二硫键通常是由两个半胱氨酸或其他形成硫醇的氨基酸的硫醇基的氧化而形成,或者通过氨基酸类似物的硫醇基的氧化而形成,从而通过连接氨基酸侧链的s原子形成人工二硫桥。具体地,可在保证另外的半胱氨酸修饰的一对结构域之间插入半胱氨酸(作为额外的氨基酸或氨基酸置换),从而通过二硫键形成而产生稳定的结构域对。一“对”抗体结构域被理解为在特定排列中的一组的两个抗体结构域,其中一个抗体结构域在其表面上或在空腔中具有与在另一个抗体结构域上的区域特异性地结合并因此为互补的区域。抗体结构域可通过β片层区的接触而结合以形成一对结构域。这种结构域对也称为(异源或同源)二聚体,该二聚体例如通过静电相互作用、重组融合或共价键合而缔合,从而使两个结构域直接物理缔合。本文中具体地描述了scvhab的cl/ch1二聚体,该二聚体是cl结构域和ch1结构域的同源对(cognatepair)。出于稳定性的原因,这种cl/ch1对尤其通过肽接头l2进一步连接,从而使该对变成单个共价多肽链。此外,可以在cl结构域和ch1域之间导入共价二硫桥,从而使结构域对相互作用稳定化。关于一对相关结构域或结构域二聚体的术语“同源”被理解为如下的结构域,每个结构域具有相互互补地结合的界面从而在每个结构域上形成域间接触表面。在彼此接触时,通过这些接触表面的缔合而形成结构域对。可通过首先筛选通过将cdr序列折叠在抗体文库的各结合位点中以选择特异性结合物所形成的抗原结合位点而产生抗体。下一步,所选择的文库成员可用作cdr序列(或亲本cdr序列,可对该序列做进一步修饰以调节抗原结合和甚至表型特性)的来源,该序列可用于将任何类型的抗体构建体工程化,例如全长免疫球蛋白或其抗原结合片段。抗体文库(例如,包含识别相同靶抗原的特异性抗体构建体的库,或者由某个动物或品种(例如,本文中所描述的转基因非人动物)产生的天然抗体文库,该文库包含识别不同靶抗原的抗体库)是指抗体的组合或集合(例如,本文中所描述的scvhab或hcab),各抗体在所选的展示系统或容器中适当地展示。特定的展示系统将给定的蛋白质(在本文中是抗体,例如本文中所描述scvhab或hcab)与其编码核酸(例如其编码mrna、cdna或基因)偶联。因此,文库的各成员包含编码在其上展示的抗体的核酸。展示系统包括但不限于细胞、病毒(如噬菌体)、核糖体、真核细胞(如酵母)、包含质粒的dna和mrna。任何抗体基因多样性文库均可用于这种目的,例如包括大量的单独文库成员,以形成抗体序列的多样性,或者采用预选的文库,该预选文库例如富含稳定化的或功能活性的文库成员。例如,展示系统可以富含结合到某个靶的文库成员。可以通过包括例如链替换方法的公知技术来构建文库。就重链替换而言,将抗体克隆到含有例如人vh基因库的载体中,以形成噬菌体抗体文库转化体。其他的方法包括对抗体的cdr的位点专一诱变、或者cdr随机化,其中利用应用含nnk密码子的诱变寡核苷酸使靶向残基进行完全随机化、或使用简约诱变使标向残基部分随机化,使部分或全部的cdr随机化,其中处在编码靶向氨基酸残基的位置的寡核苷酸含有偏向初始核苷酸碱基的混合物。可替代地,可以使用易错pcr,并且应用dntp类似物、易错聚合酶,或者在pcr反应中添加mn2+离子来构建该文库。有各种技术可用于制造编码人抗体文库构建的设计的基因。可以利用完全合成方法来生产dna,其中将序列分为重叠的片段,随后将它们制备成合成寡核苷酸。将这些寡核苷酸混合到一起并通过首先加热到约100℃然后缓慢冷却到环境温度而彼此退火。在该退火步骤后,可以直接地克隆合成组装基因,或者可以在克隆之前利用pcr使其扩增。当需要大的单罐人类文库并且大量资源可用于构建过程时,这尤其可取的。具体的方法是采用噬菌体、噬菌粒和/或酵母文库进行直接结合物选择并且内化噬菌体抗体选择。可以将用于位点专一诱变的其他方法用于文库插入物的形成,例如kunkel法(kunkel,proc.natl.acad.sci.usa.,82:488,1985)或dpni法[weiner,etal.,gene151:119,1994)。“天然文库”是指多核苷酸(或者由这种多核苷酸编码的多肽)的文库,该多核苷酸文库尚未被询问以确定对特定抗原具有特异性的抗体的存在。“天然文库”也指代不局限于对任意一组抗原或对特定抗原具有特异性的抗体序列或者对其不加偏倚或富集的文库。因此,天然文库与“成熟文库”(例如“亲和力成熟文库”)不同。天然文库也可包括“免疫前”文库,该文库是指在天然存在序列已经历抗原选择之前具有与天然存在抗体序列相似的序列多样性的文库。可设计并制备这种免疫前文库,以便反映或模仿该免疫前库,并且/或者可基于收集v、d和j基因以及重链序列(例如,公知的种系序列)的其他大型数据库获知的合理设计而进行设计和制备。在本发明的某些实施方案中,代表在人或非人库中所发现的可能的v、d和j多样性以及连接多样性(即,n1和n2)的组件被从头合成为单链或双链dna寡核苷酸。“成熟文库”是指被设计成增强或改善抗体序列的至少一个特征的文库,该抗体序列在对诸如天然文库或免疫前文库的文库进行询问以确定具有对抗原的特异性的抗体序列是否存在时被鉴定。可通过加入对应于下列的核酸序列而生成这种成熟文库:一个或多个cdr;一个或多个抗原结合区;一个或多个vh区;和/或一个或多个重链;从天然文库的查询中获得或鉴定出来的文库,这些文库被设计用于进一步在体外或体内发生诱变,以生成具有在初始(亲本)抗体的情况下所导入多样性的文库。作为阵列技术的不同例子,可以采用b细胞克隆,其在b细胞阵列中的手动或计算机可寻址位置处产生编码本文中所描述的抗体构建体的基因。可用机器人或手动方法来操纵此阵列,从而仅重新排列表达某种类型抗体的细胞、和/或特异性地识别某个靶的细胞。在某些实施方案中,使用经基因工程化的b细胞克隆,例如从经过适当免疫的非人转基因动物(如本文中所描述的动物)以产生抗体,或者使用哺乳动物细胞表达文库,或者可替代地利用抗体和蛋白质工程的标准方法和机器人工具生成稳定转化的哺乳动物细胞的大群体。可在合适的培养箱中和/或在长期保存条件下排列在平板上的可定位孔中使单独的克隆保持活力,例如可包含在液氮中在-135℃下储存的冷冻细胞悬液或者在允许所储存的细胞系恢复的其他可接受条件。本文中所使用的术语“库”应指代变体的集合,例如以靶表位或抗原特异性的多样性为特征的变体。通常,抗体的结构(也称为“支架”)在这种库中是相同的,但具有多种不同的cdr序列。如在本领域中众所周知的,存在可用于鉴定和分离具有某些结合特性和亲和力的蛋白质的多种展示和选择技术,包括例如展示技术,如细胞和非细胞方法,尤其是移动展示系统。在细胞系统中,可采用噬菌体展示、病毒展示、酵母或其他真核细胞展示,如哺乳动物或昆虫细胞展示。移动系统与采用可溶方式的展示系统有关,如体外展示系统,其中有核糖体展示、mrna展示或核酸展示。可利用任意合适方法对展示能够与靶结合的抗原结合结构的文库成员的文库进行筛选。该筛选步骤可包括一轮或数轮的选择。可采用适合于鉴定能够与靶抗原结合的抗体的任何筛选方法。特别地,各轮的选择可以包含在所述靶的存在下孵育抗体文库,从而选择与所述抗原或其表位结合的抗体。一旦鉴定出具有期望结构的抗体,便可以利用本领域中公知的方法产生这种抗体,包括例如杂交瘤技术或重组dna技术。在杂交瘤方法中,对合适的非人宿主动物(如啮齿动物或小鼠)进行免疫以激活淋巴细胞,这些淋巴细胞产生或者能够产生将特异性地结合到用于免疫的蛋白质的抗体。可替代地,可在体外免疫淋巴细胞。然后使用合适的融合剂(如聚乙二醇)使淋巴细胞与浆细胞瘤细胞发生融合,以形成杂交瘤细胞。对用于生产针对抗原的单克隆抗体的其中生长杂交瘤细胞的培养基进行测定。优选地,利用流式细胞术、免疫沉淀或者利用体外结合测定(如酶联免疫吸附测定法(elisa))对由杂交瘤细胞所产生的单克隆抗体的结合特异性进行测定。根据另一具体实例,可以通过使用共知的重组表达载体,例如包含编码抗体序列的核苷酸序列的质粒或表达组件,通过分离编码所需抗体链的dna并且用用于表达的编码序列来转染重组宿主细胞而产生重组单克隆抗体。重组宿主细胞可以是原核细胞和真核细胞。根据一具体方面,编码核苷酸序列可用于基因操作,以使抗体人源化或改善抗体的亲和力或其他特征。例如,可将恒定区工程化为类似于人恒定区。可能需要对抗体序列进行基因操作,以获得针对靶抗原的更大亲和力。对于本领域技术人员而言显而易见的是,可以对抗体执行一种或多种多核苷酸改变,并且仍然保持其针对靶(表位或抗原)的结合能力。利用各种方法产生抗体分子通常是很好理解的。在例如harlow,etal.,antibodies:alaboratorymanual2nded.,coldspringharborlaboratorypress,coldspringharbor,n.y.,(2014)中提供了与抗体产生有关的各种技术。例如可利用在培养中由连续细胞系产生抗体分子的任何方法产生单克隆抗体。制备单克隆抗体的合适方法的实例包括kohler等人,nature256:495,1975)的杂交瘤方法及人b细胞杂交瘤方法[kozbor,j.immunol.133:3001,1984;和brodeuretal.,1987,inmonoclonalantibodyproductiontechniquesandapplications,lbschook,ed.,(marceldekker,inc.,newyork),pp.51-63]。如本文中所使用的术语“靶”应指代表位或抗原。如本文中所使用的术语“抗原”应尤其包括已证实由于抗原暴露于动物的免疫系统或暴露于抗体文库而被抗体(至少一个互补位)的结合位点所识别的所有抗原和靶分子。具体地,被本文中所描述的抗体所靶向的优选抗原是已证明具有或能够具有免疫或治疗相关性的分子,特别是已对临床疗效进行了测试的分子。术语“抗原”是用于描述整个靶分子或这种分子的片段,特别是亚结构,例如靶的多肽或碳水化合物结构。这种亚结构(经常被称为“表位”,例如b细胞表位、t细胞表位)可以是免疫学相关的,即,也被天然抗体或单克隆抗体所识别。本文中所使用的术语“表位”应尤其指存在于在抗原与特异性抗体之间的界面处的分子结构,其中与表位相互作用的抗体表面被称为“互补位”。在化学上,表位可由在不连续表位中的碳水化合物序列或结构、肽序列或序列组,脂肪酸或寡核苷酸或多核苷酸所组成。如果抗原分子是有机、生物化学或无机物质,它则被称为“半抗原”。表位或半抗原可由上述物质的衍生物或任意组合所组成。如果表位是多肽,则它在肽中通常包括至少3个氨基酸、优选地8至50个氨基酸、更优选地约10-20个氨基酸。表位可以是线性表位或者不连续表位。线性表位由多肽或碳水化合物链的一级序列的单个片段组成。线性表位可以是连续的或重叠的。不连续表位示由通过折叠多肽以形成三级结构而聚集到一起的氨基酸或碳水化合物所组成,并且氨基酸不一定在线性序列中彼此相邻。具体地,表位至少是诊断相关分子的一部分,即,样品中表位的不存在或存在与疾病或患者的健康状况或者制造中的工艺状态或者环境和食物状况定性地或定量地相关。表位也可以是治疗相关分子的至少一部分,即,可以被改变疾病进程的特异性结合结构域所靶向的分子。如本文所使用的术语“特异性”或“特异性结合”是指决定异源分子群体中的目标同源配体的结合反应。因此,在指定的条件下(例如,免疫测定条件),抗体结合到其特定靶并且不以显著的量结合到样品中存在的其他分子。特异性结合表示该结合在选择的靶识别,高、中或低结合亲和力或亲合力方面是选择性的。如果结合常数或结合动力学与样品中的竞争靶相差至少10倍、优选地相差至少100倍、更优选地至少1000倍,通常实现选择性结合。特异性结合不排除与相似抗原、或者不同物种(类似物)的相同抗原的某些程度的交叉反应。例如,结合实体也可优选地与类似于人靶的啮齿动物或灵长类动物靶发生交叉反应,以便于临床前动物研究。如本文所使用的术语“基因座”是指编码被表达产物的dna的dna编码序列或片段,即基因组序列,如宿主生物体的基因组的一部分、或载体的一部分,例如整合于靶位点,例如在所限定的限制位点或同源性的区域。可以设计限制位点以确保将表达组件插入适当的阅读框中。通常,将外来(在本文中也称为外源的)dna插入载体dna的一个或多个限制位点,然后由载体连同可传播载体dna运载到宿主细胞中。通常,基因座包含至少一个基因。术语“基因座”并不意味着基因被主动转录或完整。可能包含已灭活的基因。在本文中所描述的具体实施方案中,转基因动物的内源性κ和λ轻链基因座通过一种或多种修饰(例如功能丢失突变,或内源性κ和/或λ轻链基因座或其一部分的缺失)而失去功能。示例性的适当修改如下地理解。为了使κ链基因座失活,通过重组酶介导的盒式交换(rmce)策略删除vκ2-137、最cκ末端vκ基因片段和jκ5、最cκ近端jκ之间的整个3.2mb基因组区域。这是通过在vκ2-137下游和jκ5下游插入适当的靶向序列,然后是体外cre介导的干预基因组区域缺失来实现的。使用类似的策略来灭活λ链基因座。rmce删除了含有小鼠λv基因片段(iglv)的整个194kb区。在这种情况下,将适当的靶向序列插入iglv2的上游和iglv1的下游,然后进行体外cre介导的干预基因组区域缺失。可将基因座工程化以表达编码抗体的外显子,例如本文进一步描述的。可以使用各种常规技术针对位点特异性编辑和/或重组产生重组基因座。优选地,通过将含有编码例如cl-l2-ch1的基因片段的dna片段(在本文中称为“供体dna”)插入非人动物免疫球蛋白基因座的修饰形式中来产生修饰基因座,例如宿主生物体的重链基因座(在本文中称为“受体等位基因”)。受体等位基因可含有针对位点特异性dna重组酶(例如cre重组酶)的识别位点(loxp位点和loxp位点的突变版本)。供体dna的两侧可以是相同的cre重组酶识别位点(在5'端和3'端的两端,例如,一侧有一个loxp位点,而另一侧有突变版本的loxp位点)。cre重组酶可用于催化供体dna插入受体等位基因。在可选择的实施方案中,基因片段主要(如果不是排他性的话)通过同源重组被导入免疫球蛋白基因座。在这种实施方案中,采用靶向序列或载体,其由基因组靶向同源性臂组成,所述基因组靶向同源性臂侧接包含编码基因片段的抗体的核酸序列(即,5'端和3'端的核苷酸序列,其与靶序列同源并能够与靶序列杂交)。这些基因组同源性臂有助于将编码dna的抗体插入免疫球蛋白基因座,例如编码免疫球蛋白重链的dna。术语“靶向序列”是指与细胞基因组中的dna序列同源的序列,其在待修饰的免疫球蛋白遗传基因座区域的两侧或与其相邻存在。两侧或相邻序列可以在基因座本身内或在宿主细胞基因组中编码序列的上游或下游。将靶向序列插入重组dna载体中以用于细胞转染,使得待插入细胞基因组中的序列(例如重组位点的序列)位于载体的靶向序列的两侧。在许多情况下,其中采用同源重组完成基因组中的遗传变化,例如插入或缺失,进一步的修饰将牵涉到工程化的位点特异性核酸內切酶的使用,以增加可实现预计结果的可能性。这种核酸内切酶是有价值的,因为它们可以被工程化为对靶基因组中的独特序列具有高度特异性,并且因为它们在它们识别的位点引起双链dna断裂。双链断裂促进与靶向载体的同源重组,所述靶向载体在断裂的紧邻区域携带与dna的靶向同源性。因此,与使用单独的靶向载体相比,靶向载体和在载体靶向的区域内或附近切割dna的位点特异性核酸内切酶的组合通常导致更高的同源重组效率。此外,有可能通过使用一种或多种位点特异性核酸内切酶和由两个靶向同源臂组成的靶向载体来促进基因组缺失的产生,其中一个臂靶向待缺失区域的一侧而另一臂靶向另一侧。位点特异性重组与一般同源重组的不同之处在于,重组酶识别所需要的短特异性dna序列是发生重组的唯一位点。位点特异性重组需要专门的重组酶来识别位点并催化这些位点处的重组。已经显示出许多噬菌体和酵母来源的位点特异性重组系统,每个都包含重组酶和特异性同源靶位点,为了dna整合的目的在真核细胞中起作用,因此适用于本文中所描述中的用途。这些包括噬菌体p1cre/lox,酵母flp-frt系统和位点特异性重组酶酪氨酸家族的dre系统。这种系统和使用方法在现有技术中已被很好地描述。通过使用野生型和突变型loxp(或frt等)位点以及适当的重组酶(例如,cre或flp)以及阴性和/或阳性选择的组合促进重组酶介导的盒式交换(rmce)程序。当使用的位点彼此相同和/或没有选择时,将发生rmce,但是由于有利于切除而不是插入反应,降低了工艺效率,并且(不包括阳性选择)将没有富集用于适当突变的细胞。酪氨酸家族的其他系统,例如噬菌体λint整合酶、hk2022整合酶,以及另外属于单独的重组酶丝氨酸家族的系统,例如噬菌体phic31、r4tp901整合体,已知使用它们各自的重组位点在哺乳动物细胞中起作用,并且也适用于本文所述的用途。本文中所描述的方法具体地利用位点特异性重组位点,这些位点利用相同的重组酶,但不促进位点之间的重组。例如,可以将loxp位点和突变的loxp位点整合到宿主的基因组中,但将cre导入宿主中将不导致两个位点发生重组;相反,loxp位点将与另一个loxp位点发生重组,而突变位点将仅与另一个同样突变的loxp位点发生重组。两种类型的变体重组酶位点可用于促进重组酶介导的盒式交换。一个在该位点的8bp间隔区内具有突变,而另一个在13bp的反向重复序列中具有突变。间隔突变体,如lox511(hoess,etal.,nucleicacidsres.,14:2287,1986)、lox5171和lox2272(leeandsaito,gene,216:55,1998)、m2、m3、m7和mil(langeretal.,nucleicacidsres.,30:3067,2002)容易与自身发生重组,但具有显著降低的与野生型位点发生重组的速率。通过重组酶介导的盒式交换将此类的突变位点用于dna插入的用途的实例可以参见baerandbode,curr.opin.biotechnol.,12:473,2001。反向重复突变体代表第二类的变体重组酶位点。例如,loxp位点可在左反向重复序列(le突变体)或右反向重复序列(re突变体)中含有改变的碱基。le突变体lox71在左反向重复序列的5'末端具有5个bp,它们从野生型序列变为taccg(araki,nucleicacidsres.,25:868,1997)。类似地,re突变体lox66具有变为cggta的5个最靠近3'的碱基。反向重复突变体可以用于将质粒插入物整合到染色体dna中。例如,可以将le突变体用作“供体”re突变体重组入其中的“靶”染色体loxp位点。在重组后,将发现含有re位点的供体dna片段被插入基因组中,该基因组的一侧是双突变位点(同时含有le和re反向重复突变),而另一侧是野生型位点(leeandsadowski,prog.nucleicacidres.mol.biol.,80:1,2005)。该双突变体与野生型位点充分不同,即它不能被cre重组酶所识别,因此插入的片段不能被在两个位点之间的cre介导的重组所切除。在某些方面,与编码核酸区或调控序列相反,可以将位点特异性重组位点导入内含子或基因间区域。这可避免在将位点特异性重组位点插入动物细胞的基因组中时无意破坏正确基因表达所必需的任何调控序列或编码区。可以利用常规的同源重组技术来实现位点特异性重组位点的导入。这种技术描述于参考文献中,例如,sambrookandrussell(2001)molecularcloning:alaboratorymanual,3ded.(coldspringharbor,n.y.:coldspringharborlaboratorypress)和nagy,(2003)manipulatingthemouseembryo:alaboratorymanual,3ded.(coldspringharbor,n.y.:coldspringharborlaboratorypress)。可以使用本领域已知的被设计用于阳性或阴性选择的载体来促进进入基因组中的特异性重组。为了便于对已经历置换反应的细胞的鉴定,可采用适当的遗传标志系统,并且通过例如选择培养基的使用来选择细胞。然而,为了确保基因组序列在置换区间的两个端点处或相邻位置基本上没有外源核酸序列,可取地可以在选择含有被置换核酸的细胞之后除去该标志系统/基因。重组酶可以以纯化蛋白的形式而提供,或者可由在细胞内瞬时表达的构建体所表达,从而提供重组酶活性。可替代地,该细胞可用于形成转基因动物,该转基因动物可与表达所述重组酶的动物杂交,从而产生缺少标志基因和相关的重组位点的后代。在本文中,关于基因的术语“内源的”表示该基因是产自于细胞,即该基因存在于未修饰细胞的基因组中的特定基因座。内源基因可以是存在于野生型细胞中的该基因座上的野生型基因(如在自然界中所发现)。如果内源基因存在于与野生型基因相同的基因组中的基因座,则内源基因可以是修饰内源基因。这种修饰内源基因的实例是将外来核酸插入其中的基因。内源基因可存在于核基因组、线粒体基因组等中。在可选择实施方案中,通过使用非同源末端连接方法的crispr/cas9技术,将基因片段导入免疫球蛋白基因座中(例如参见heetal.,nuc.acidsres.,44:e85,2016),而不是通过通常用于该系统的同源定向修复。本文中所使用的“载体”被定义为对于克隆重组核苷酸序列的转录所需的dna序列,即重组基因的转录和它们mrna在合适宿主生物体中的转译。载体包括质粒和病毒以及任何dna或rna分子(无论是否自我复制),它们可以用于转化、转导或转染细胞。载体可包含自主复制的核苷酸序列以及整合有核苷酸序列的基因组。表达载体可另外地包含用于在宿主细胞或基因组整合位点中的自主复制的起点、一种或多种可选择标记(例如,氨基酸合成基因或赋予抗生素(如嘌呤霉素、zeocintm、卡那霉素、g418或潮霉素)耐药性的基因)、许多限制酶切割位点、合适的启动子序列和转录终止子,将这些组分可操作地连接到一起。常见类型的载体是“质粒”,其通常是双链dna的自含分子,该双链dna可以容易地接受另外的(外来)dna并且可以容易地导入合适的宿主细胞中。质粒常常含有编码dna和启动子dna,并且具有适于插入外来dna的一个或多个限制性位点。具体地,术语“质粒”是指可以将dna或rna序列(例如外来基因)导入宿主细胞中从而转化宿主并促进导入序列的表达(例如,转录和转译)的载体。本文所使用的术语“宿主细胞”应指经转化以产生特定重组蛋白(如本文中所描述的抗体)的原代受试细胞、及其任何后代。应该理解的是,并非所有后代都与亲本细胞完全相同(由于故意或无意的突变或环境差异),然而这种改变的后代包括在这些术语中,只要该后代保留与最初转化的细胞相同的功能。术语“宿主细胞系”是指用于表达重组基因以产生重组多肽(如重组抗体)的宿主细胞的细胞系。本文中所使用的术语“细胞系”是指已获得在长时间内增殖能力的特定细胞类型的已建立克隆。可将这种宿主细胞或宿主细胞系维持在细胞培养物中并且/或者进行培养以产生重组多肽。本文中所使用的关于核酸、抗体或其他化合物的术语“分离的”或“分离”应指与其天然缔合的环境充分地分离从而以“大体上纯的”形式而存在的这种化合物。“分离的”不一定表示将与其他化合物或材料的人工或合成混合物,或者存在不干扰基本活性的杂质排除,这些杂质可能存在,例如由于不完全的纯化。尤其是,本文中所描述的分离的核酸分子还表示包括化学合成的那些。就本文中所描述的核酸而言,有时使用术语“分离的核酸”。当应用于dna时,该术语是指与其中其来源的生物体的天然存在基因组中与其紧密邻接的序列相分离的dna分子。例如,“分离的核酸”可包含被插入载体(如质粒或病毒载体)中或者被整合到原核或真核细胞或宿主生物的基因组dna中的dna分子。当应用于rna时,术语“分离的核酸”主要是指由如上定义的分离dna分子所编码的rna分子。可替代地,该术语可指代已从将会以其天然状态(即,在细胞或组织中)与之缔合的其他核酸充分地分离的rna分子。“分离的核酸”(dna或者rna)还可代表利用生物方法或合成方法直接产生并且与在其产生期间所存在的其他组分分离的分子。就多肽或蛋白质(如分离的抗体)而言,术语“分离的”应具体地指没有或基本上没有与它们天然地缔合的物质的化合物,例如在其天然环境或当这种制备是通过体外或体内实施的重组dna技术时其中制备它们的环境(例如,细胞培养)中与它们一起发现的其他化合物。分离的化合物可以与稀释剂或佐剂一起配制,并且仍然为了实际目的而分离,例如当用于诊断或治疗时可以将多肽或多核苷酸与药学上可接受的载体或赋形剂混合。本文中所描述的抗体尤其是以分离的形式提供,其基本上没有针对不同靶抗原的其他抗体并且/或者包含不同结构排列的抗体结构域。但是,可将分离的抗体包括在组合制剂中,该组合制剂含有分离抗体的组合,例如与具有至少一种其他抗体(如具有不同特异性的单克隆抗体或抗体片段)的组合。具体地,本文中所描述的抗体是以大体上纯的形式提供。本文中所使用的术语“大体上纯的”或“纯化的”应指包含至少50%(w/w),优选地至少60%、70%、80%、90%或95%的化合物(如核酸分子或抗体)的制剂。利用适合于该化合物的方法(例如,色谱法、聚丙烯酰胺凝胶电泳、hplc分析等)测量纯度。本文中所描述的抗体可具体地使用于药物组合物。因此,提供一种药物组合物,该药物组合物包含本文中所描述的抗体及药学上可接受的载体或赋形剂。根据本发明,这些药物组合物可以以推注或输注的方式或者通过连续输注而给药。适于促进这种给药方法的药物载体在本领域中是公知的。药学上可接受载体通常包括与本发明所提供的免疫球蛋白生理学相容的任何和所有合适的溶剂、分散介质、包衣、等渗剂和吸收延迟剂等。药学上可接受载体的其他实例包括无菌水、生理盐水、磷酸盐缓冲盐水、右旋糖、甘油、乙醇等,以及它们的任意组合。其它的药学上可接受载体在本领域中是已知的,并且在例如remington’spharmaceuticalsciences(gennaro,ar,ed.,mackprintingco)中有描述。液体制剂可以是溶液剂、乳剂或混悬剂,并且可以包括赋形剂(如悬浮剂、增溶剂、表面活性剂、防腐剂和螯合剂)。用于肠胃外给药的示例性制剂包括适合于皮下、肌内或静脉内注射的制剂,例如溶液剂、乳剂或混悬剂。就化合物(例如本文中所描述的抗体)的给药而言,本文所用的术语“治疗有效量”是指当施用于受试者时足以获得有益或期望的结果(包括临床结果)的量或活性。因此,其有效量或同义的数量取决于应用它的上下文。有效量是用来表示足以治疗、预防或抑制这种疾病或病症的化合物的量。在疾病的情况下,如本文中所描述的治疗有效量的抗体具体地是用于治疗、调节、缓解、逆转、或影响得益于抗体与其靶抗原相互作用的疾病或病症。对应于这种有效量的化合物的量将根据各种因素而变化,如给定的药物或化合物、药物制剂、给药途径、疾病或病症的类型、待治疗的受试者或宿主的身份等,但可以由本领域技术人员按常规方法确定。术语“重组的”是指在宿主细胞中不天然存在的多核苷酸或多肽。重组分子可含有以非天然存在的方式连接在一起的两个或多个天然存在序列。重组细胞含有重组多核苷酸或多肽。如果细胞接受了重组核酸,核酸对细胞则是“外源的”。术语“重组的”尤其表示“利用基因工程制备或者由基因工程得到”。可替代地,使用术语“工程化”。例如,可通过将各个亲本序列工程化以产生工程抗体或结构域,来修饰抗体或结构域以产生变体。重组宿主具体地包含表达载体或克隆载体,或者它已经过基因工程化而含有重组核酸序列,尤其是使用对宿主为外来的核苷酸序列。通过在宿主中表达各个重组核酸而产生重组蛋白。本文所使用的术语“重组抗体”包括免疫球蛋白及尤其是利用重组方法而制备、表达、产生或分离的抗体,诸如:a)从用于人免疫球蛋白基因或由其所制备的杂交瘤的转基因或转染色体的动物(例如非人动物,如小鼠)中分离的抗体,b)从转化以表达抗体的宿主细胞分离的抗体,例如从转染瘤中,c)从重组、组合抗体库中分离的抗体,和d)利用包含将例如人免疫球蛋白基因序列剪接到其他dna序列的任何其他方法所制备、表达、产生、或分离的抗体。这种重组抗体包含经工程化而包括重排和突变的抗体,这些重排和突变例如在抗体成熟期间发生。“位点特异性重组”是指在两个相容的重组位点之间的重组过程,包括以下三个事件中的任何事件:a)位于重组位点侧面的预选核酸的删除;b)位于重组位点侧面的预选核酸的核苷酸序列的转化,和c)紧邻位于不同核酸分子上的重组位点的核酸区域的相互交换。应当理解的是,如果一个或两个核酸分子是圆形的,核酸片段的这种相互交换则导致整合事件。本文中所使用术语“转基因”是用来描述已经或将要被人工插入细胞(尤其是宿主动物细胞)的基因组中的遗传物质。如本文中所使用的术语“转基因”是指核酸分子,例如采用表达构建体和/或靶向载体形式的核酸。“转基因动物”表示非人动物,通常是哺乳动物或禽类,例如啮齿动物,尤其是小鼠或大鼠,尽管其他哺乳动物是可想到的,具有在一部分的其细胞中以染色体或染色体外元件的形式而存在或者稳定地整合到其种系dna中(即,在其大部分或全部细胞的基因组序列中)的外源核酸序列。在实施方案的某些方面,本发明的转基因动物进一步包含人免疫球蛋白区域。例如,已开发出许多方法,所述方法采用人免疫球蛋白序列置换内源性小鼠免疫球蛋白区域以产生用于药物发现目的的部分或完全人抗体。这种小鼠的实例包括在例如美国专利号7,145,056;7,064,244;7,041,871;6,673,986;6,596,541;6,570,061;6,162,963;6,130,364;6,091,001;6,023,010;5,593,598;5,877,397;5,874,299;5,814,318;5,789,650;5,661,016;5,612,205;和5,591,669中所描述的小鼠。在特别有利的方面,本发明的转基因动物包括嵌合免疫球蛋白片段,如在wabl和killeen的美国公布号2013/0219535中所描述的。这种转基因动物具有包含被导入的人免疫球蛋白区域的基因组,其中被导入的区域包含基于非人脊椎动物的内源基因组的人可变区编码序列和非编码可变序列。优选地,本发明的转基因细胞和动物具有其中部分或全部的内源性免疫球蛋白区域被去除的基因组。在另一有利方面,对动物的基因组含量进行修饰,使得它们的b细胞能够在每个细胞中表达多于一个的功能性vh结构域,即,这些细胞产生双特异性抗体,如wo2017035252a1中所描述。参考以下实施例,将更全面地理解前面的描述。然而,这些实施例仅是实施本发明一个或多个实施方案的方法的代表,而不应理解为限制本发明的范围。实施例实施例1:基于iggfab片段的晶体结构的最小接头2长度的估计。接头2(l2,图2)将cl(cκ或cλ)连接到ch1。基于图3中所示的fab结构[pdb2xkn,egfr(表皮生长因子受体)抗体7a7的fab片段的晶体结构,其是igg1κmab],连接cκ的cooh末端和ch1的nh2末端的桥键的距离为(在图3中用虚线表示)。由于cκ和ch1的相对位置,接头必须更长以便连接c末端和n末端。(ggggs[seqidno:35])4接头的理论长度为其小于两个末端之间距离的覆盖范围的两倍因此,在这种情况下,建议的最小接头长度为(ggggs[seqidno:35])4并且最大为(ggggs[seqidno:35])16。实施例2:编码hcab的表达载体的体外测试。在生成表达scvhab或hcab的转基因小鼠之前,构建了编码跨模和分泌形式的hcab的多种表达载体,并测试了体外表达。这些不同的构建物在若干方面有所不同:l2长度、cl(cκ、cλ1或cλ2)的组成、ch(igg1或igg2a)的组成、以及编码hcab蛋白(nh-vh-cκ-l2-ch1-ch2-ch3-tm-cooh相对于nh-vh-ch1-l2-cκ-ch2-ch3-tm-cooh)中的cl和ch1结构域的顺序。还构建并测试了若干阳性和阴性对照载体:已知在细胞表面上表达的阳性对照-常规的h2l2igg抗体、缺少ch1结构域和任何lc的骆驼样igg、具有连接的vκ和vh但没有cl或ch1的scfvigg抗体。不在细胞表面上表达的阴性对照-常规的igg抗体但没有lc(h2l0)。使用脂质体2000(lipofectamine2000)(invitrogen)将表达载体转染到hek293t细胞中。将编码具有myc标签的人cd4的表达载体进行共转染,并将hcd4表达用作测定转染效率的对照。此外,将所有hek293t细胞与表达小鼠cd79a和cd79b(igα/igβ)两者的构建体进行了共转染,小鼠cd79a和cd79b是抗原受体的表面表达所需的共受体,包括膜结合形式的hcab(wienands和engles,int.rev.immunol.,20:679,2001)。20-24小时后,为了检测细胞表面hcd4、小鼠igg1(migg1)和小鼠κ轻链(migκ)而将细胞染色,通过流式细胞术进行分析以检测细胞表面及细胞内hcab和hcd4蛋白表达。在与细胞相关的myc、gapdh和hcab及分泌的hcab的免疫印迹(wb)检测中,将细胞裂解,在转染后的40-48小时收集上清液。相同的上清液也用于利用elisa对分泌的hcab进行定量。含有cκ的hcab在细胞表面上被表达。图4描述了在第一轮实验中所使用的表达载体。阳性对照:1.常规的h2l2igg抗体;2.缺少ch1结构域和lc的骆驼样igg;3.scfvigg抗体,具有连接的vκ和vh但没有cl或ch1。本文中所描述的hcab:4.nh-vh-cκ-l2-ch1-ch2-ch3-tm-cooh。本文中所描述的hcab,但在hcab中cκ和ch1结构域的顺序颠倒:5.nh-vh-ch1-l2-cκ-ch2-ch3-tm-cooh。阴性对照:没有lc(h2l0)的常规igg抗体。图5描绘了利用流式细胞术对目标蛋白质的细胞表面表达的分析。在所有转染的细胞系中,表达hcd4的细胞的频率相似(范围为61-64%),从而表明在所有情况下(上排)具有相似的转染效率。正如预计(中间行)的,阳性对照、常规的migg1(1)、骆驼样ab(2)和scfv(3)均在细胞表面上表达。本文中所描述的hcab(4)相似地表达。然而,如果hcab中cκ和ch1结构域的顺序颠倒(5),则不再有任何细胞表面表达。与表达水平相关的细胞表面migg1染色的平均荧光强度(mfi)随着不同的构建体而变化(表1)。作为阴性对照的没有轻链的常规igg1(6)不在细胞表面上表达。正如预计的,仅在使用构建体1和4的情况下观察到表面migκ的表达,因为它们是仅有的含cκ的表面表达的hcab。为确保细胞表面migg1表达的缺乏不是因为蛋白质在细胞内部被降解,而将转染子固定、透化再用同一组的抗体对细胞内蛋白进行染色(图6,将这些样品的细胞内migg1染色的mfi示于表2)。用构建体5和6转染的细胞含有大量的细胞内migg1重链,但不在细胞表面上表达。因此,某些主动机制必须将这些分子保留在细胞内。在由构建体6编码的h2l0hcab中,已知该保留是由于部分未折叠的ch1结构域与er分子伴侣bip的缔合所导致(haasandwabl,nature,306:387,1983;bole,etal.,jcellbiol.102:1558,1986)。表1.细胞表面migg1染色的平均荧光强度(mfi)。数据来自图5中的流式细胞术分析。构建体编号mfi13841227293531042965表2.细胞内migg1染色的平均荧光强度(mfi)。数据来自图6中的流式细胞术分析。构建体编号mfi1569624240360034164675159065736含有cλ1或cλ2的hcab也在细胞表面上表达。还使用图7a中所示构建体测试了在hcab中改变cl结构域的效果。阳性对照构建体1编码骆驼样igg1,构建体2-4编码分别含有cκ、cλ1和cλ2的本文中所描述的hcab。尽管mfi有变化,但所有hcab均在细胞表面上表达(图8,中间行)(表3)。这里未对cλ3进行测试,但预计结果与cλ2相同,这是因为cλ2和cλ3的氨基酸序列几乎相同(99%的氨基酸一致性)。将编码构建体3的融合基因的示意图示于图7b。该构建体中的vh外显子由vh3-11、dh2-21和jh4所编码,cl外显子是由cλ1所编码,并且ch和铰链(h)外显子是来自igg1。本领域技术人员将认识到,可以将任何vh、dh、jh、cl或ch基因插入以取代这里所描述的构建体的各个成分。表3.细胞表面migg1染色的平均荧光强度(mfi)。数据来自图8中的流式细胞术分析。构建体编号mfi13072221693456543400hcab的分泌也用图7中所示的分泌形式的构建体转染hek293t细胞,以便利用wb(图9和图10)和酶联免疫测定(elisa,图12)对hcab分泌进行测试。在这些实验中也检查了将l2长度从6个重复序列变更为10个重复序列以及将ch结构域从igg1变更为igg2a的效果。将用于转染效率的wb对照(抗myc抗体)、和上样对照(抗gapdh抗体)示于图11中。如利用wb所评估的,骆驼样ab显示最佳分泌(图9左侧,泳道1,非还原性条件;图10左侧,泳道1,还原条件)。在被检查的hcab构建体中,编码具有cλ1的migg1和10个ggggs(seqidno:35)重复序列的接头的一个hcab构建体显示最佳分泌(图9左,泳道4,非还原条件;图10左,泳道4,还原条件)。将接头长度从6个重复序列增加到10个也改善vh-cκhcab的分泌(比较图9中的泳道2和3,左图),推测这是通过改善杂hcab分子的折叠及其从er分子伴侣中的释放而实现。甚至可通过进一步增大接头长度而更加地改善hcab分泌。与骆驼样ab相比相对较低的hcab分泌水平可能是由于hcab的细胞内蛋白复合物的形成所致。可以看出,这些高分子量带在表达migg1hcab的转染子的细胞裂解物中相比在骆驼样裂解物中更加丰富(图9,在右图中比较了泳道1与泳道2-5)。这些是二硫键连接的复合物而不是非特异性聚集体,因为它们在还原条件下消失(图10,右图)。利用elisa测定对用wb进行分析的相同上清液中的hcab进行了定量(图12)。与wb数据一致,具有cλ1和10个ggggs(seqidno:35)重复序列的接头的migg1显示出hcab中的最佳分泌(约840ng/ml),这仅比骆驼样抗体少约3.8倍。实施例3:利用同源重组将小鼠cl-l2-ch1-h-ch2-ch3_s-tm基因盒导入在ighm上游位置的内源小鼠igh基因座中以产生hcab。图13中示出了用于导入cl-l2-ch1-h-ch2-ch3_s-tm基因盒以产生hcab的示例性方法。图13a和图13b中描绘了靶向载体(注意到,在此附图中被标记为a.和b.且用虚线连接的片段在靶向载体中是连续的)。盒的cl成分可以编码cκ或者cλ(cλ1、cλ2或cλ3)。(注意到,本文中也描述的含有接头1的盒l1-cl-l2-ch1-h-ch2-ch3_s-tm可以相同的方式导入。)同源重组靶向载体的两个基本成分是短同源臂(sha)和长同源臂(lha),它们与位于被修饰的内源基因座区域侧面的同源dna片段共有序列一致性(图13c)。在这种情况下,sha由位于相应的小鼠jh非编码序列(seqidno:2)侧面的人jh2-jh6基因片段组成。lha由完整的ighm基因组成,起始于5'内含子并终止于3'utr(seqidno:21)。起始于5’端的靶向载体的其他显著特征包括:1)pgk_tk_pa(seqidno:1),其是由磷酸甘油酸酯激酶启动子(pgk)所驱动的单纯疱疹病毒(hsv)胸苷激酶(tk)基因并且包括聚a位点(pa)。该因素是用于使用更昔洛韦对已整合靶向载体但未经过同源重组的细胞进行阴性选择;这种细胞将保留hsv-tk基因并被杀死。2)t3噬菌体rna聚合酶的t3启动子(seqidno:3)。该dna依赖性rna聚合酶对t3噬菌体启动子具有高度特异性。99kd酶催化体外rna合成,这允许从少量b细胞或杂交瘤中对vdj重排进行快速克隆。3)cag_puror_pa,其是由强cag启动子所驱动的嘌呤霉素耐药基因并且包括polya位点(seqidno.:5)。该因素是用于基于嘌呤霉素耐药性对已整合靶向载体的细胞进行阳性选择。4)注意到,已将靶向载体稳定地整合到其基因组中的细胞将对更昔洛韦和嘌呤霉素两者具有耐药性。5)注意到,cag_puror_pa元件位于frt位点(seqidno:4和seqidno:6)的侧面,frt位点可在鉴定正确靶向的es细胞克隆之后用于通过提供flp重组酶而在体外或体内去除该元件。eμ增强子(seqidno:7)被包括在cl-l2-ch1-h-ch2-ch3_s-tm基因盒的上游位置,用以促进基因座的转录。在靶向载体中接在其后的是ighmlha(seqidno:21)。靶向载体缺少在内源性igh基因座中存在的μ开关(s)区,因而靶向基因座也将缺少s区,因此不能进行同种型转换。通过电穿孔将靶向载体导入es细胞中。使细胞在增补有更昔洛韦和嘌呤霉素的培养基中生长。然后使用广泛实行的基因靶向策略,用位于新导入cl-l2-ch1-h-ch2-ch3_s-tm基因盒的5'和3'内部的引物,利用基因组pcr监测存活的分离es细胞克隆是否成功进行了基因靶向。通过使用映射到位于sha的5'侧的侧面的dna序列的探针、映射到位于lha的3'侧的侧面的dna序列的第二探针、和映射在载体中基因组一致性的两个臂之间的新型dna内的第三探针,利用基因组southern印迹对靶向盒的正确整合做进一步验证。(在图13d和e中描绘了正确靶向的基因座的结构。注意到,标记为d.和e.且在附图中用虚线连接的片段在es细胞的igh基因座中是连续的。)使用原位荧光杂交法对经pcr和southern印迹法验证的es细胞克隆的核型进行分析,该原位荧光杂交法被设计用于辨别在小鼠es细胞中发生的最常发生的染色体畸变。将具有这种畸变的克隆排除不再使用。基于pcr和southern印迹数据判断具有预期的正确基因组结构,并且基于核型分析也没有可检测染色体畸变的es细胞克隆被选择用于进一步使用。将在小鼠重链基因座中携带正确靶向的cl-l2-ch1-h-ch2-ch3_s-tm基因盒的es细胞克隆显微注射到取自dba/2品系的小鼠胚泡中,根据标准程序形成部分地由es细胞得到的嵌合体小鼠。选择在其皮毛中具有最高水平的es细胞源性贡献的雄性嵌合体小鼠与雌性小鼠交配。这里所选择的雌性小鼠属于c57b1/6ntac品系,该品系在其种系中携带编码flp重组酶的转基因。分析了这些交配的后代是否存在cl-l2-ch1-h-ch2-ch3_s-tm基因盒,以及是否缺失位于frt侧面的嘌呤霉素耐药性基因(图13f和图13g。注意到,在体内的igh基因座中,标记为f.和g.且在附图中用虚线连接的片段是连续的)。将正确靶向的小鼠用于建立小鼠群体。实施例4:利用同源重组将小鼠cl-l2-ch1_s-tm基因盒导入在ighm上游位置的内源小鼠igh基因座中以产生scvhab。本实施例在所有方面与实施例3相同,除了基因修饰的小鼠产生scvhab而不是hcab以外。这通过修饰基因盒(靶向载体)使其具有结构cl-l2-ch1_s-tm而不是cl-l2-ch1-h-ch2-ch3_s-tm来实现。靶向载体的结构和序列与实施例3相同,用于将载体导入es细胞以选择和分析正确的同源重组并建立小鼠群体的方法与实施例3是相同的(注意到,可以将本文中也描述的含有接头1的盒l1-cl-l2-ch1_s-tm以相同的方式导入)。实施例5:利用重组酶介导的盒式交换(rmce)将小鼠cl-l2-ch1-h-ch2-ch3_s-tm基因盒导入在ighm上游位置的内源小鼠igh基因座中以产生hcab。实施例5a:rmce受体等位基因的形成。这里的目的是将puro_tk融合基因导入在eμ下游位置和在ighm上游位置的内源igh基因座的区域中,该puro_tk融合基因在上游位于突变frt位点和突变loxp位点的侧面并且在下游位于wtfrt和loxp位点的侧面(图14b)。在此构型中,突变体和wtfrt或loxp位点在flp或cre重组酶的存在下不能重组,但可以整合供体dna的片段,该供体dna在其5'和3'末端分别具有相应的突变和wt位点(图14a)。使用与图13中相同的sha和lha通过同源重组导入puro_tk融合基因。靶向载体的另一个特征是在sha上游5'端存在白喉毒素a(dta)基因(seqidno:34)。该因素用于针对已整合了靶向载体但未经过同源重组的细胞进行阴性选择;这些细胞保留了dta基因,并在表达毒素时被杀死。通过电穿孔将靶向载体导入es细胞中,使细胞在增补有嘌呤霉素的培养基中生长。然后使用广泛实行的基因靶向策略,用位于新导入的puro_tk融合基因盒5'和3'内部的引物,利用基因组pcr监测存活的分离es细胞克隆是否成功进行了基因靶向。使用映射到位于sha的5'侧的侧面的dna序列的探针、映射到位于lha的3'侧的侧面的dna序列的第二探针、和映射在载体中基因组一致性的两个臂之间的新dna内部的第三探针,通过基因组southern印迹,对靶向基因座的结构做进一步验证(图14c中描绘了正确靶向基因座的结构)。利用原位荧光杂交法对es细胞的pcr和southern印迹法阳性克隆的核型进行分析,该原位荧光杂交法被设计用于辨别在小鼠es细胞中发生的最常发生的染色体畸变。将具有这种畸变的克隆排除不再使用。基于pcr和southern印迹数据判断具有预期的正确基因组结构,并且基于核型分析也没有可检测染色体畸变的es细胞克隆被选择用于如下所述的进一步使用。实施例5b:利用重组酶介导的盒式交换(rmce)导入cl-l2-ch1-h-ch2-ch3_s-tm基因盒。rmce靶向载体(图15a)的序列与图13a和图13b中所示的同源区域是相同的,除了该载体位于突变frt和loxp位点的5'端的侧面及wtfrt和loxp位点的3'端的侧面外(注意到,可以将本文中也描述的l1-cl-l2-ch1-h-ch2-ch3_s-tm盒以相同的方式导入)。载体被导入到实施例5a中产生的rmce修饰的es细胞中(图15b)与用于瞬时表达cre或flp重组酶的载体和靶向载体一起被整合,得到图15c、15d中所示的基因组结构。(注意到,标记为c.和d.并附图中通过虚线连接的片段在体内的igh基因座是连续的)。未通过rmce正确整合靶向载体的es细胞保留了puro_tk基因并通过将更昔洛韦添加到生长培养基中被杀死。然后使用广泛采用的基因靶向策略,通过位于新导入的cl-l2-ch1-h-ch2-ch3_s-tm基因盒5'和3'内的引物,通过基因组pcr监测存活的分离的es细胞克隆是否成功进行了基因靶向。通过使用映射到5'侧、3'侧的侧面以及新型dna内的dna序列的探针的基因组southern印迹对靶向基因座的结构做进一步验证。使用原位荧光杂交方法对es细胞的pcr和southern印迹法阳性克隆的的核型进行分析,该原位荧光杂交法被设计用于区分小鼠es细胞中发生的最常发生的染色体畸变。将具有这种畸变的克隆排除不再使用。基于pcr和southern印迹数据判断具有预期的正确基因组结构,并且基于核型分析也没有可检测的染色体畸变的es细胞克隆被选择用于进一步使用。将在小鼠重链基因座中携带正确靶向的cl-l2-ch1-h-ch2-ch3_s-tm基因盒的es细胞克隆显微注射到dba/2品系的小鼠胚泡中,根据标准程序创建部分es细胞衍生的嵌合体小鼠。选择在其皮毛中具有最高水平的es细胞源性贡献的雄性嵌合体小鼠与雌性小鼠交配。分析这些交配的后代中是否存在cl-l2-ch1-h-ch2-ch3_s-tm基因盒。使用正确靶向的小鼠建立小鼠群体。实施例6:利用重组酶介导的盒式交换(rmce)将小鼠cl-l2-ch1_s-tm基因盒导入到在ighm上游位置的内源小鼠igh基因座中以产生scvhab。本实施例在所有方面与实施例5相同,除了经遗传修饰的小鼠产生scvhab而不是hcab以外。这通过将靶向载体修饰为具有结构cl-l2-ch1_s-tm而不是cl-l2-ch1-h-ch2-ch3_s-tm来实现。靶向载体的结构和序列与实施例5相同,用于将载体导入es细胞以便选择和分析正确的同源重组并且建立小鼠群体的方法也与实施例5相同。(注意到,可以将本文中也描述的l1-cl-l2-ch1_s-tm盒以相同的方式导入)。序列表<110>特里安尼公司(trianni,inc.)<120>单链vh和重链抗体<130>redl-1111<160>49<170>patentinversion3.5<210>1<211>2182<212>dna<213>人工序列<220><223>pgk_tk_pa<400>1agtcagcttctgatggaattagaacttggcaaaacaatactgagaatgaagtgtatgtgg60aacagaggctgctgatctcgttcttcaggctatgaaactgacacatttggaaaccacagt120acttagaaccacaaagtgggaatcaagagaaaaacaatgatcccacgagagatctataga180tctatagatcatgagtgggaggaatgagctggcccttaatttggttttgcttgtttaaat240tatgatatccaactatgaaacattatcataaagcaatagtaaagagccttcagtaaagag300caggcatttatctaatcccaccccacccccacccccgtagctccaatccttccattcaaa360atgtaggtactctgttctcacccttcttaacaaagtatgacaggaaaaacttccatttta420gtggacatctttattgtttaatagatcatcaatttctgcagacttacaggacggatcgat480cccctcagttagcctcccccatctcccgggcaaacgtgcgcgccaggtcgcatatcgtcg540gtatggagccgggggtggtgacgtgggtctggaccatcccggaggtaagttgcagcaggg600cgtcccggcagccggcgggcgattggtcgtaatccaggataaagacgtgcatggaacgga660ggcgtttggccaagacgtccaaggcccaggcaaacacgttatacaggtcgccgttggggg720ccagcaactcgggggcccgaaacagggtaaataacgtgtccccgatatggggtcgtgggc780ccgcgttgctctggggctcggcaccctggggcggcacggccgtccccgaaagctgtcccc840agtcctcccgccacgacccgccgcactgcagataccgcaccgtattggcaagtagcccgt900aaacgcggcgaatcgcagccagcatagccaggtccagccgctcgccggggcgctggcgtt960tggccaggcggtcgatgtgtctgtcctccggaagggccccaagcacgatgttggtgccgg1020gcaaggtcggcgggatgagggccacgaacgccagcacggcctggggggtcatgctgccca1080taaggtaccgcgcggccgggtagcacaggagggcggcgatgggatggcggtcgaagatga1140gggtgagggccgggggcggggcatgtgagctcccagcctcccccccgatatgaggagcca1200gaacggcgtcggtcacggcataaggcatgcccattgttatctgggcgcttgtcattacca1260ccgccgcgtccccggccgatatctcaccctggtcgaggcggtgttgtgtggtgtagatgt1320tcgcgattgtctcggaagcccccagcacccgccagtaagtcatcggctcgggtacgtaga1380cgatatcgtcgcgcgaacccagggccaccagcagttgcgtggtggtggttttccccatcc1440cgtggggaccgtctatataaacccgcagtagcgtgggcattttctgctccgggcggactt1500ccgtggcttcttgctgccggcgagggcgcaacgccgtacgtcggttgctatggccgcgag1560aacgcgcagcctggtcgaacgcagacgcgtgttgatggccggggtacgaggccatgatgg1620caaggacggtgcattggctgcaggtcgaaaggcccggagatgaggaagaggagaacagcg1680cggcagacgtgcgcttttgaagcgtgcagaatgccgggcctccggaggaccttcgggcgc1740ccgccccgcccctgagcccgcccctgagcccgcccccggacccaccccttcccagcctct1800gagcccagaaagcgaaggagcaaagctgctattggccgctgccccaaaggcctacccgct1860tccattgctcagcggtgctgtccatctgcacgagactagtgagacgtgctacttccattt1920gtcacgtcctgcacgacgcgagctgcggggcgggggggaacttcctgactaggggaggag1980tagaaggtggcgcgaaggggccaccaaagaacggagccggttggcgcctaccggtggatg2040tggaatgtgtgcgaggccagaggccacttgtgtagcgccaagtgcccagcggggctgcta2100aagcgcatgctccagactgccttgggaaaagcgcctcccctacccggtagaattgacctg2160ccggggccctcgaatcctgcag2182<210>2<211>2508<212>dna<213>小鼠<400>2gggtctataattactctgatgtctaggaccagggggctcaggtcactcaggtcaggtgag60tcctgcatctggggactgtggggttcaggtggcctaaggcaggatgtggagagagtttta120gtataggaacagaggcagaacagagactgtgctactggtacttcgatgtctggggcacag180ggaccacggtcaccgtctcctcaggtaagctggcttttttctttctgcacattccattct240gaaacgggaaaagatattctcagatctccccatgtcaggccatctgccacactctgcatg300ctgcagaagcttttctgtaaggatagggtcttcactcccaggaaaagaggcagtcagagg360ctagctgcctgtggaacagtgacaatcatggaaaataggcatttacattgttaggctaca420tgggtagatgggtttttgtacacccactaaaggggtctatgatagtgtgactactttgac480tactggggccaaggcaccactctcacagtctcctcaggtgagtccttacaacctctctct540tctattcagcttaaatagattttactgcatttgttgggggggaaatgtgtgtatctgaat600ttcaggtcatgaaggactagggacaccttgggagtcagaaagggtcattgggagccctgg660ctgacgcagacagacatcctcagctcccatacttcatggccagagatttatagggatcct720ggccagcattgccgctaggtccctctcttctatgctttctttgtccctcactggcctcca780tctgagatcatcctggagccctagccaaggatcatttattgtcaggggtctaatcattgt840tgtcacaatgtgcctggtttgcttactggggccaagggactctggtcactgtctctgcag900gtgagtcctaacttctcccattctaaatgcatgttggggggattctgggccttcaggacc960aagattctctgcaaacgggaatcaagattcaacccctttgtcccaaagttgagacatggg1020tctgggtcagggactctctgcctgctggtctgtggtgacattagaactgaagtatgatga1080aggatctgccagaactgaagcttgaagtctgaggcagaatcttgtccagggtctatcgga1140ctcttgtgagaattaggggctgacagttgatggtgacaatttcagggtcagtgactgtct1200ggtttctctgaggtgaggctggaatataggtcaccttgaagactaaagaggggtccaggg1260gcttctgcacaggcagggaacagaatgtggaacaatgacttgaatggttgattcttgtgt1320gacaccaggaattggcataatgtctgagttgcccaggggtgattctagtcagactctggg1380gtttttgtcgggtatagaggaaaaatccactattgtgattactatgctatggactactgg1440ggtcaaggaacctcagtcaccgtctcctcaggtaagaatggcctctccaggtctttattt1500ttaacctttgttatggagttttctgagcattgcagactaatcttggatatttgtccctga1560gggagccggctgagagaagttgggaaataaactgtctagggatctcagagcctttaggac1620agattatctccacatctttgaaaaactaagaatctgtgtgatggtgttggtggagtccct1680ggatgatgggatagggactttggaggctcatttgaagaagatgctaaaacaatcctatgg1740ctggagggatagttggggctgtagttggagattttcagtttttagaataaaagtattagt1800tgtggaatatacttcaggaccacctctgtgacagcatttatacagtatccgatgcatagg1860gacaaagagtggagtggggcactttctttagatttgtgaggaatgttccgcactagattg1920tttaaaacttcatttgttggaaggagagctgtcttagtgattgagtcaagggagaaaggc1980atctagcctcggtctcaaaagggtagttgctgtctagagaggtctggtggagcctgcaaa2040agtccagctttcaaaggaacacagaagtatgtgtatggaatattagaagatgttgctttt2100actcttaagttggttcctaggaaaaatagttaaatactgtgactttaaaatgtgagaggg2160ttttcaagtactcatttttttaaatgtccaaaattcttgtcaatcagtttgaggtcttgt2220ttgtgtagaactgatattacttaaagtttaaccgaggaatgggagtgaggctctctcata2280acctattcagaactgacttttaacaataataaattaagtttcaaatatttttaaatgaat2340tgagcaatgttgagttggagtcaagatggccgatcagaaccagaacacctgcagcagctg2400gcaggaagcaggtcatgtggcaaggctatttggggaagggaaaataaaaccactaggtaa2460acttgtagctgtggtttgaagaagtggttttgaaacactctgtccagc2508<210>3<211>19<212>dna<213>人工序列<220><223>t3启动子<400>3cctttagtgagggttaatt19<210>4<211>48<212>dna<213>人工序列<220><223>5'frt位点<400>4gaagttcctattccgaagttcctattcttcaaaaggtataggaacttc48<210>5<211>2711<212>dna<213>人工序列<220><223>cag_puror_pa<400>5tcattctggcctccccctcccccaaggtcattctggcctccccctcccccaaggtcattc60tggcctccccctcccccttggccgcgcctcattctggcctccccctcccccaaggcacac120aaaaaaccaacacacagatctattgaaaataatgatcttttattgatccgcgcctggatc180tcgaaccggttcaggcaccgggcttgcgggtcatgcaccaggtgcgcggtccttcgggca240cctcgacgtcggcggtgacggtgaagccgagccgctcgtagaaggggaggttgcggggcg300cggaggtctccaggaaggcgggcaccccggcgcgctcggccgcctccactccggggagca360cgacggcgctgcccagacccttgccctggtggtcgggcgagacgccgacggtggccagga420accacgcgggctccttgggccggtgcggcgccaggaggccttccatctgttgctgcgcgg480ccagccgggaaccgctcaactcggccatgcgcgggccgatctcggcgaacaccgcccccg540cttcgacgctctccggcgtggtccagaccgccaccgcggcgccgtcgtccgcgacccaca600ccttgccgatgtcgagcccgacgcgcgtgaggaagagttcttgcagctcggtgacccgct660cgatgtggcggtccgggtcgacggtgtggcgcgtggcggggtagtcggcgaacgcggcgg720cgagggtgcgtacggcccgggggacgtcgtcgcgggtggcgaggcgcaccgtgggcttgt780actcggtcatggaaggctaggcggatcggcgatcgctccggaggccgccctgggagtgga840cacctgtggagagaaaggcaaagtggatgtcaatgtcactcaagtgtatggccagatctc900aagcctgccacacctcaagcttgacaacaaaaagattgtcttttctgaccagatggacgc960ggccaccctcaaaggcatcaccgcgggccaggtgaatatcaaatcctcctcgtttttgga1020aactgacaatcttagcgcagaagtcatgcccgcttttgagagggagtactcaccccaacg1080cttctagagccgccggtcacacgccagaagccgaaccccgccctgccccgtcccccccga1140aggcagccgtccccccgcggacagccccgaggctggagagggagaaggggacggcggcgc1200ggcgacgcacgaaggccctccccgcccatttccttcctgccggcgccgcaccgcttcgcc1260ccgcgcccgctagagggggtgcggcggcgcctcccagatttcggctccgcacagatttgg1320gacaaaggaagtccctgcgccctctcgcacgattaccataaaaggcaatggctgcggctc1380gccgcgcctcgacagccgccggcgctccgggggccgccgcgcccctcccccgagccctcc1440ccggcccgaggcggccccgccccgcccggcacccccacctgccgccaccccccgcccggc1500acggcgagccccgcgccacgccccgtacggagccccgcacccgaagccgggccgtgctca1560gcaactcggggaggggggtgcagggggggttgcagcccgaccgacgcgcccacaccccct1620gctcacccccccacgcacacaccccgcacgcagcctttgttcccctcgcagccccccccg1680caccgcggggcaccgcccccggccgcgctcccctcgcgcacactgcggagcgcacaaagc1740cccgcgccgcgcccgcagcgctcacagccgccgggcagcgcggagccgcacgcggcgctc1800cccacgcacacacacacgcacgcaccccccgagccgctccccccgcacaaagggccctcc1860cggagcccctcaaggctttcacgcagccacagaaaagaaacaagccgtcattaaaccaag1920cgctaattacagcccggaggagaagggccgtcccgcccgctcacctgtgggagtaacgcg1980gtcagtcagagccggggcgggcggcgcgaggcggcggcggagcggggcacggggcgaagg2040cagcgcgcagcgactcccgcccgccgcgcgcttcgctttttatagggccgccgccgccgc2100cgcctcgccataaaaggaaactttcggagcgcgccgctctgattggctgccgccgcacct2160ctccgcctcgccccgccccgcccctcgccccgccccgccccgcctggcgcgcgccccccc2220cccccccgcccccatcgctgcacaaaataattaaaaaataaataaatacaaaattggggg2280tggggaggggggggagatggggagagtgaagcagaacgtggggctcacctcgaccatggt2340aatagcgatgactaatacgtagatgtactgccaagtaggaaagtcccataaggtcatgta2400ctgggcataatgccaggcgggccatttaccgtcattgacgtcaatagggggcgtacttgg2460catatgatacacttgatgtactgccaagtgggcagtttaccgtaaatactccacccattg2520acgtcaatggaaagtccctattggcgttactatgggaacatacgtcattattgacgtcaa2580tgggcgggggtcgttgggcggtcagccaggcgggccatttaccgtaagttatgtaacgcg2640gaactccatatatgggctatgaactaatgaccccgtaattgattactattaataactagt2700caataatcaat2711<210>6<211>48<212>dna<213>人工序列<220><223>3'frt位点<400>6gaagttcctattccgaagttcctattcttcaaaaggtataggaacttc48<210>7<211>313<212>dna<213>小鼠<400>7ctgcagcagctggcaggaagcaggtcatgtggcaaggctatttggggaagggaaaataaa60accactaggtaaacttgtagctgtggtttgaagaagtggttttgaaacactctgtccagc120cccaccaaaccgaaagtccaggctgagcaaaacaccacctgggtaatttgcatttctaaa180ataagttgaggattcagccgaaactggagaggtcctcttttaacttattgagttcaacct240tttaattttagcttgagtagttctagtttccccaaacttaagtttatcgacttctaaaat300gtatttagaattc313<210>8<211>1000<212>dna<213>小鼠<400>8ttaaaaaaaaaaaaaaaggaaagggacttctctgtgtttggcaacacaagtgcgatgcac60aggcaggaagatcaaatctgtcccaacaatacaggggacagagggtcaacctacaaaagg120aaagaacctggggcagtgtgaagacaacactgtagaagccaaggctgagttcactgagct180ctcgttagtgagactacacagcaaggaggtggcgggcactgagcagtgaggccccgggaa240gtgggggtgatggtggtgacggtggtaactgttaagaactgggggaaagaattgtggaga300accaagctaaatagttatgtcaaaccacatgtttaggagcctgggttgacttcataggga360gtaggcatggaggctaatctagaggtttgtgtataggcaagaagtgaatcctgacccaag420aatagagagtgctaaacggacttagttcaaagacaactgaaaaagacaatgcctgcaaaa480caaagctaaggccagagctcttggactatgaagagttcagggaacctaagaacagggacc540atctgtgtacaggccaaggccggtagaagcagcctaggaagtgtcaagagccaacgtggc600tgggtgggcaaagacaggaagggactgttaggctgcagggatgtgccgacttcaatgtgc660ttcagtattgtccagattgtgtgcagccatatggcccaggtataagaggtttaacagtgg720aacacagatgcccacatcagacagctggggggcgggggtgaacacagatacccatactgg780aaagcaggtggggcattttcctaggaacgggactgggctcaatggcctcaggtctcatct840ggtctggtgatcctgacattgataggcccaaatgttggatatcacctactccatgtagag900agtcggggacatgggaagggtgcaaaagagcggccttctagaaggtttggtcctgtcctg960tcctgtctgacagtgtaatcacatatactttttcttgtag1000<210>9<211>64<212>dna<213>小鼠<400>9ataatacactgaaatggagcccttccttgttacttcataccatcctctgtgcttccttcc60tcag64<210>10<211>320<212>dna<213>小鼠<400>10gggctgatgctgcaccaactgtatccatcttcccaccatccagtgagcagttaacatctg60gaggtgcctcagtcgtgtgcttcttgaacaacttctaccccaaagacatcaatgtcaagt120ggaagattgatggcagtgaacgacaaaatggcgtcctgaacagttggactgatcaggaca180gcaaagacagcacctacagcatgagcagcaccctcacgttgaccaaggacgagtatgaac240gacataacagctatacctgtgaggccactcacaagacatcaacttcacccattgtcaaga300gcttcaacaggaatgagtgt320<210>11<211>15<212>dna<213>人工序列<220><223>接头2<400>11ggaggaggggggtcc15<210>12<211>289<212>dna<213>小鼠<400>12aaaacgacacccccatctgtctatccactggcccctggatctgctgcccaaactaactcc60atggtgaccctgggatgcctggtcaagggctatttccctgagccagtgacagtgacctgg120aactctggatccctgtccagcggtgtgcacaccttcccagctgtcctgcagtctgacctc180tacactctgagcagctcagtgactgtcccctccagcacctggcccagccagaccgtcacc240tgcaacgttgcccacccggccagcagcaccaaggtggacaagaaaattg289<210>13<211>96<212>prt<213>小鼠<400>13lysthrthrproproservaltyrproleualaproglyseralaala151015glnthrasnsermetvalthrleuglycysleuvallysglytyrphe202530progluprovalthrvalthrtrpasnserglyserleusersergly354045valhisthrpheproalavalleuglnseraspleutyrthrleuser505560serservalthrvalproserserthrtrpproserglnthrvalthr65707580cysasnvalalahisproalaserserthrlysvalasplyslysile859095<210>14<211>39<212>dna<213>小鼠<400>14tgcccagggattgtggttgtaagccttgcatatgtacag39<210>15<211>321<212>dna<213>小鼠<400>15tcccagaagtatcatctgtcttcatcttccccccaaagcccaaggatgtgctcaccatta60ctctgactcctaaggtcacgtgtgttgtggtagacatcagcaaggatgatcccgaggtcc120agttcagctggtttgtagatgatgtggaggtgcacacagctcagacgaaaccccgggagg180agcagatcaacagcactttccgttcagtcagtgaacttcccatcatgcaccaggactggc240tcaatggcaaggagttcaaatgcagggtcaacagtgcagctttccctgcccccatcgaga300aaaccatctccaaaaccaaag321<210>16<211>320<212>dna<213>小鼠<400>16gcagaccgaaggctccacaggtgtacaccattccacctcccaaggagcagatggccaagg60ataaagtcagtctgacctgcatgataacaaacttcttccctgaagacattactgtggagt120ggcagtggaatgggcagccagcggagaactacaagaacactcagcccatcatggacacag180atggctcttacttcgtctacagcaagctcaatgtgcagaagagcaactgggaggcaggaa240atactttcacctgctctgtgttacatgagggcctgcacaaccaccatactgagaagagcc300tctcccactctcctggtaaa320<210>17<211>132<212>dna<213>小鼠<400>17gggctgcaactggacgagacctgtgctgaggcccaggacggggagctggacgggctctgg60acgaccatcaccatcttcatcagcctcttcctgctcagtgtgtgctacagcgctgctgtc120acactcttcaag132<210>18<211>81<212>dna<213>小鼠<400>18gtaaagtggatcttctcctcggtggtggagctgaagcagacactggttcctgaatacaag60aacatgattgggcaagcaccc81<210>19<211>1340<212>dna<213>小鼠<400>19taggccacctcctgtaatggcatttcccaggccccgaaggaccctgtccaatatgccaag60cagcacaactgagatcacactgtctgctcatctcgctttcctccgaccccgagactcagc120tactctcaaattttccctctctgaaggaccatgtggacattacattgctccaggccacag180ccaccaggacctaaaacaccatcacagcagcaccaaagacactggatagacccacaaggg240caatagtttcctcaacagtatatccaaactgttgggacaaacgagcaatcactgaagaag300tgacaagttcccacaatgtcagtgtccagctgagaagggacaaaaagtggtaccagccct360gtccacaccaccttctaattcacaggaatacgtgatagaagaggcaggttgtagatccga420aagatgagacagattttatcaactccagaaagagctgggtccaactgaattattctagcg480accttggcattgtcatgacctgccatgaccttcctccttaacacttcgataaaccctggg540atatggaaaatgcctgtgtttctcagggtttgggaaagaaccatccatgttgggattctt600gtgtagatcctccttctggtcacagatgcaatacactggattttcaggcaaaggagcaaa660ttcacagacaactctggccctacagccctaagacctagacaccaccatctccttggaatt720atcaaatttaacacccggcacacaacaaagaaggactgggactttgaggcctttgtgtag780ccctagagggggcagaggccactgagcagggattgggtgatcacaaggacctcctggaga840gggacctgaggagcaggttccaattgggccaaagaaagaagaacaacaatagagatgaag900gatgctggaaagagccatggtacagcagtcttgtccttcagacatgactcttacagccca960ggactcttacagtagctagctggagcagaagtccaagggattaccatgccctagggccac1020aggctactggagggtggagtgagtctactacacaggtccaatgcctgtttctccattgct1080tctcagccaatgagaaatcagagtctccacctccaagaaaaaggaaggtggaaatgaaag1140gtgagcacctgccttcccgtgactggcagaaagatctccacggactcaaggctttgactt1200caaacttcccgtagatccctgatgtctttaaggactctgtctgctttgttggttttgttt1260gtttgttttgtactttgtcaataaaacatttttcaaatattctacaattgctgtgctctt1320cctatgcaaactggccctgg1340<210>20<211>660<212>dna<213>小鼠<400>20cacttcaaaacatggcacagttaagttgaccagtgggccatgcagagcatactacccctc60ctggtcctgtgtcctaccagatatcctctaagtgtcccttcactgctgggctccagttct120gctgccctgaaccacacaggtcctcagtctgtcctccctatgggcagtttcatcccagcc180agaagctgccctgtggcccctaggctgcccaggcatggtctcccacaccaaccacacaaa240ctaagaagcctgtcccatacattgacctcaggctcagtgataacatttctatagaaaaag300gaaaggaataagaaaaaaaaaactccatattacatgggggctgcctaccttgctgatgct360tattgtctcaagcaagtttggagaagttccaaatcaggcactgacagggtgggttctcag420ccatgctcttcttacctggtaggtgccttctcccccatggcacctcacaggctctccatc480tgtgtgtgtctgggtcctgatctcttctcataagtacaaagtcaggctggaagaggtaca540ccctagccctcattataacttaccagtttatgatcctgtctgcaaacatctgaggtccct600gtggctcaaatgtcaattagtgagtcttctggggacagaatttagtccacattagctctc660<210>21<211>4566<212>dna<213>小鼠<400>21ggtgttcatgcccctagagttggctgaagggccagatccacctactctagaggcatctct60ccctgtctgtgaaggcttccaaagtcacgttcctgtggctagaaggcagctccatagccc120tgctgcagtttcgtcctgtataccaggttcacctactaccatatctagccctgcctgcct180taagagtagcaacaaggaaatagcagggtgtagagggatctcctgtctgacaggaggcaa240gaagacagattcttacccctccatttctcttttatccctctctggtcctcagagagtcag300tccttcccaaatgtcttccccctcgtctcctgcgagagccccctgtctgataagaatctg360gtggccatgggctgcctggcccgggacttcctgcccagcaccatttccttcacctggaac420taccagaacaacactgaagtcatccagggtatcagaaccttcccaacactgaggacaggg480ggcaagtacctagccacctcgcaggtgttgctgtctcccaagagcatccttgaaggttca540gatgaatacctggtatgcaaaatccactacggaggcaaaaacaaagatctgcatgtgccc600attccaggtaagaaccaaaccctcccagcaggggtgcccaggcccaggcatggcccagag660ggagcagcggggtggggcttaggccaagctgagctcacaccttgacctttcattccagct720gtcgcagagatgaaccccaatgtaaatgtgttcgtcccaccacgggatggcttctctggc780cctgcaccacgcaagtctaaactcatctgcgaggccacgaacttcactccaaaaccgatc840acagtatcctggctaaaggatgggaagctcgtggaatctggcttcaccacagatccggtg900accatcgagaacaaaggatccacaccccaaacctacaaggtcataagcacacttaccatc960tctgaaatcgactggctgaacctgaatgtgtacacctgccgtgtggatcacaggggtctc1020accttcttgaagaacgtgtcctccacatgtgctgccagtgagtggcctgggctaagccca1080atgcctagccctcccagattagggaagtcctcctacaattatggccaatgccacccagac1140atggtcatttgctccttgaactttggctccccagagtggccaaggacaagaatgagcaat1200aggcagtagaggggtgagaatcagctggaaggaccagcatcttcccttaagtaggtttgg1260gggatggagactaagcttttttccaacttcacaactagatatgtcataacctgacacagt1320gttctcttgactgcaggtccctccacagacatcctaaccttcaccatccccccctccttt1380gccgacatcttcctcagcaagtccgctaacctgacctgtctggtctcaaacctggcaacc1440tatgaaaccctgaatatctcctgggcttctcaaagtggtgaaccactggaaaccaaaatt1500aaaatcatggaaagccatcccaatggcaccttcagtgctaagggtgtggctagtgtttgt1560gtggaagactggaataacaggaaggaatttgtgtgtactgtgactcacagggatctgcct1620tcaccacagaagaaattcatctcaaaacccaatggtaggtatccccccttcccttcccct1680ccaattgcaggacccttcctgtacctcatagggagggcaggtcctcttccaccctatcct1740cactactgtcttcatttacagaggtgcacaaacatccacctgctgtgtacctgctgccac1800cagctcgtgagcaactgaacctgagggagtcagccacagtcacctgcctggtgaagggct1860tctctcctgcagacatcagtgtgcagtggcttcagagagggcaactcttgccccaagaga1920agtatgtgaccagtgccccgatgccagagcctggggccccaggcttctactttacccaca1980gcatcctgactgtgacagaggaggaatggaactccggagagacctatacctgtgttgtag2040gccacgaggccctgccacacctggtgaccgagaggaccgtggacaagtccactggtaaac2100ccacactgtacaatgtctccctgatcatgtctgacacaggcggcacctgctattgaccat2160gctagcgctcaaccaggcaggccctgggtgtccagttgctctgtgtatgcaaactaacca2220tgtcagagtgagatgttgcattttataaaaattagaaataaaaaaaatccattcaaacgt2280cactggttttgattatacaatgctcatgcctgctgagacagttgtgttttgcttgctctg2340cacacaccctgcatacttgcctccaccctggcccttcctctaccttgccagtttcctcct2400tgtgtgtgaactcagtcaggcttacaacagacagagtatgaacatgcgattcctccagct2460acttctagatatatggctgaaagcttgcctaacctggtgcaggcagcattcaggcacata2520tatagacacacatgcatttatacatagatatataggtacacatgtgtagacacatacatg2580aatgtgtattcatggacacacagacaaaggtacacatatatacacatgagttcatgcgca2640cacacatgcatggacacttacaaacgccttcagagacaaataggcatagacacacaacca2700ctcacagaaacagataccaatatgcatggtcctgtgtacacagaaacagactataggcaa2760atatacacaaataaactatatagatacaaagatatgcatatacacacatgtacagaaaca2820tcttcacatgtgtacactaacatgtgaacaggtatagcacacagatacacctggactctg2880accagggctgtaatctccaaggctcacggctcagagagcctacactaggctgggtcactg2940atactcctcaggagcccactctatgattgggagagataaccccaggtacaaagtatgcct3000atctgtctcaacaccatggggcagaagatactccactaaccacccatgacagaaagttag3060ccttggctgtgtctccattaatagaacacctcagaagaccaatgtgaaattgcctaaccc3120actcacacccaccctgatctccagttcaaaatgcagaaaacataatgcagttgtccaaaa3180gatgccccaaccacacacacacacacacacacacacacacacacacacacacacacacac3240acacacacacaccatcaaggagcctctgtaaggagtcaccacccaataacactgcctctt3300tgggctcatatcctggacattcttcatattcatatccatttggggcctaggctttagata3360tccccaagggctcatctttacagggatcagagatcccaataaatgccctggtcccacagc3420ctccctcaggtatctgtctgtttatctcttggtaccaagacccaacattgctggcagggg3480taggacaagcaacgcacgggaactctgatcaaagaaagtcatgagatgcctgagtccttc3540aggaagtaaggagggacaacctctggtatccctgttcttattgctaaagcccaagagaca3600gggagacctgctctaaattctcagtctaaacagcaccgatggcaccacctgctcagggaa3660agtccagagcacaccaatatcattttgccacagttcctgagtctgcctttacccaggtcc3720atacattgcatctgtcttgcttgctctgctgccccagggctcctggaacaaaggctccaa3780attagtgtgtcctacagcttggcctgttctgtgcctccgtctagcttgagctattagggg3840accagtcaatactcgctaagattctccagaaccatcagggcaccccaacccttatgcaaa3900tgctcagtcaccccaagacttggcttgaccctccctctctgtgtcccttcatagaggggg3960aggtgaatgctgaggaggaaggctttgagaacctgtggaccactgcctccaccttcatcg4020tcctcttcctcctgagcctcttctacagcaccaccgtcaccctgttcaaggtagtgtggt4080tgtggggctgaggacacagggctgggacagggagtcaccagtcctcactgcctctacctc4140tactccctacaagtggacagcaattcacactgtctctgtcacctgcaggtgaaatgactc4200tcagcatggaaggacagcagagaccaagagatcctcccacagggacactacctctgggcc4260tgggatacctgactgtatgactagtaaacttattcttacgtctttcctgtgttgccctcc4320agcttttatctctgagatggtcttctttctagactgaccaaagactttttgtcaacttgt4380acaatctgaagcaatgtctggcccacagacagctgagctgtaaacaaatgtcacatggaa4440ataaatactttatcttgtgaactcactttattgtgaaggaatttgttttgtttttcaaac4500ctttcctgcggtgttgacagcccaaggattatctgaatagagcttaggaactggaaatgg4560aacagt4566<210>22<211>45<212>dna<213>智人<400>22gagcccaaatcttgtgacaaaactcacacatgcccaccgtgccca45<210>23<211>36<212>dna<213>智人<400>23gagcgcaaatgttgtgtcgagtgcccaccgtgccca36<210>24<211>51<212>dna<213>智人<400>24gagctcaaaaccccacttggtgacacaactcacacatgcccacggtgccca51<210>25<211>36<212>dna<213>智人<400>25gagtccaaatatggtcccccatgcccatcatgccca36<210>26<211>57<212>dna<213>智人<400>26gttccctcaactccacctaccccatctccctcaactccacctaccccatctccctca57<210>27<211>18<212>dna<213>智人<400>27gttcccccacctccccca18<210>28<211>39<212>dna<213>小鼠<400>28tgcccagggattgtggttgtaagccttgcatatgtacag39<210>29<211>48<212>dna<213>小鼠<400>29gagcccagagggcccacaatcaagccctgtcctccatgcaaatgccca48<210>30<211>66<212>dna<213>小鼠<400>30gagcccagcgggcccatttcaacaatcaacccctgtcctccatgcaaggagtgtcacaaa60tgccca66<210>31<211>63<212>dna<213>小鼠<400>31gagcccagagtgcccataacacagaacccctgtcctccactcaaagagtgtcccccatgc60gca63<210>32<211>49<212>dna<213>小鼠<400>32gagcctagaatacccaagcccagtacccccccaggttcttcatgcccac49<210>33<211>33<212>dna<213>小鼠<400>33ggtcctactcctcctcctcctattactattcct33<210>34<211>573<212>dna<213>白喉杆菌<400>34gatgatgttgttgattcttctaaatcttttgtgatggaaaacttttcttcgtaccacggg60actaaacctggttatgtagattccattcaaaaaggtatacaaaagccaaaatctggtaca120caaggaaattatgacgatgattggaaagggttttatagtaccgacaataaatacgacgct180gcgggatactctgtagataatgaaaacccgctctctggaaaagctggaggcgtggtcaaa240gtgacgtatccaggactgacgaaggttctcgcactaaaagtggataatgccgaaactatt300aagaaagagttaggtttaagtctcactgaaccgttgatggagcaagtcggaacggaagag360tttatcaaaaggttcggtgatggtgcttcgcgtgtagtgctcagccttcccttcgctgag420gggagttctagcgttgaatatattaataactgggaacaggcgaaagcgttaagcgtagaa480cttgagattaattttgaaacccgtggaaaacgtggccaagatgcgatgtatgagtatatg540gctcaagcctgtgcaggaaatcgtgtcaggcga573<210>35<211>5<212>prt<213>人工序列<220><223>接头<400>35glyglyglyglyser15<210>36<211>318<212>dna<213>小鼠<400>36ggccagcccaagtcttcgccatcagtcaccctgtttccaccttcctctgaagagctcgag60actaacaaggccacactggtgtgtacgatcactgatttctacccaggtgtggtgacagtg120gactggaaggtagatggtacccctgtcactcagggtatggagacaacccagccttccaaa180cagagcaacaacaagtacatggctagcagctacctgaccctgacagcaagagcatgggaa240aggcatagcagttacagctgccaggtcactcatgaaggtcacactgtggagaagagtttg300tcccgtgctgactgttcc318<210>37<211>315<212>dna<213>小鼠<400>37ggtcagcccaagtccactcccactctcaccgtgtttccaccttcctctgaggagctcaag60gaaaacaaagccacactggtgtgtctgatttccaacttttccccgagtggtgtgacagtg120gcctggaaggcaaatggtacacctatcacccagggtgtggacacttcaaatcccaccaaa180gagggcaacaagttcatggccagcagcttcctacatttgacatcggaccagtggagatct240cacaacagttttacctgtcaagttacacatgaaggggacactgtggagaagagtctgtct300cctgcagaatgtctc315<210>38<211>313<212>dna<213>小鼠<400>38gtcagcccaagtccactcccacactcaccatgtttccaccttcccctgaggagctccagg60aaaacaaagccacactcgtgtgtctgatttccaatttttccccaagtggtgtgacagtgg120cctggaaggcaaatggtacacctatcacccagggtgtggacacttcaaatcccaccaaag180aggacaacaagtacatggccagcagcttcttacatttgacatcggaccagtggagatctc240acaacagttttacctgccaagttacacatgaaggggacactgtggagaagagtctgtctc300ctgcagaatgtct313<210>39<211>107<212>prt<213>智人<400>39argthrvalalaalaproservalpheilepheproproseraspglu151015glnleulysserglythralaservalvalcysleuleuasnasnphe202530tyrproargglualalysvalglntrplysvalaspasnalaleugln354045serglyasnserglngluservalthrgluglnaspserlysaspser505560thrtyrserleuserserthrleuthrleuserlysalaasptyrglu65707580lyshislysvaltyralacysgluvalthrhisglnglyleuserser859095provalthrlysserpheasnargglyglucys100105<210>40<211>105<212>prt<213>智人<400>40glnprolysalaasnprothrvalthrleupheproproserserglu151015gluleuglnalaasnlysalathrleuvalcysleuileseraspphe202530tyrproglyalavalthrvalalatrplysalaaspglyserproval354045lysalaglyvalgluthrthrlysproserlysglnserasnasnlys505560tyralaalasersertyrleuserleuthrprogluglntrplysser65707580hisargsertyrsercysglnvalthrhisgluglyserthrvalglu859095lysthrvalalaprothrglucysser100105<210>41<211>105<212>prt<213>智人<400>41glnprolysalaalaproservalthrleupheproproserserglu151015gluleuglnalaasnlysalathrleuvalcysleuileseraspphe202530tyrproglyalavalthrvalalatrplysalaaspserserproval354045lysalaglyvalgluthrthrthrproserlysglnserasnasnlys505560tyralaalasersertyrleuserleuthrprogluglntrplysser65707580hisargsertyrsercysglnvalthrhisgluglyserthrvalglu859095lysthrvalalaprothrglucysser100105<210>42<211>105<212>prt<213>智人<400>42glnprolysalaalaproservalthrleupheproproserserglu151015gluleuglnalaasnlysalathrleuvalcysleuileseraspphe202530tyrproglyalavalthrvalalatrplysalaaspserserproval354045lysalaglyvalgluthrthrthrproserlysglnserasnasnlys505560tyralaalasersertyrleuserleuthrprogluglntrplysser65707580hislyssertyrsercysglnvalthrhisgluglyserthrvalglu859095lysthrvalalaprothrglucysser100105<210>43<211>106<212>prt<213>智人<400>43glyglnprolysalaalaproservalthrleupheproproserser151015glugluleuglnalaasnlysalathrleuvalcysleuileserasp202530phetyrproglyalavallysvalalatrplysalaaspglyserpro354045valasnthrglyvalgluthrthrthrproserlysglnserasnasn505560lystyralaalasersertyrleuserleuthrprogluglntrplys65707580serhisargsertyrsercysglnvalthrhisgluglyserthrval859095glulysthrvalalaproalaglucysser100105<210>44<211>106<212>prt<213>智人<400>44glyglnprolysalaalaproservalthrleupheproproserser151015glugluleuglnalaasnlysalathrleuvalcysleuvalserasp202530pheasnproglyalavalthrvalalatrplysalaaspglyserpro354045vallysvalglyvalgluthrthrlysproserlysglnserasnasn505560lystyralaalasersertyrleuserleuthrprogluglntrplys65707580serhisargsertyrsercysargvalthrhisgluglyserthrval859095glulysthrvalalaproalaglucysser100105<210>45<211>98<212>prt<213>智人<400>45alaserthrlysglyproservalpheproleualaproserserlys151015serthrserglyglythralaalaleuglycysleuvallysasptyr202530pheprogluprovalthrvalsertrpasnserglyalaleuthrser354045glyvalhisthrpheproalavalleuglnserserglyleutyrser505560leuserservalvalthrvalproserserserleuglythrglnthr65707580tyrilecysasnvalasnhislysproserasnthrlysvalasplys859095lysval<210>46<211>113<212>prt<213>智人<400>46procysproalaprogluleuleuglyglyproservalpheleuphe151015proprolysprolysaspthrleumetileserargthrprogluval202530thrcysvalvalvalaspvalserhisgluaspprogluvallysphe354045asntrptyrvalaspglyvalgluvalhisasnalalysthrlyspro505560argglugluglntyrasnserthrtyrargvalvalservalleuthr65707580valleuhisglnasptrpleuasnglylysglutyrlyscyslysval859095serasnlysalaleuproalaproileglulysthrileserlysala100105110lys<210>47<211>107<212>prt<213>智人<400>47glyglnproarggluproglnvaltyrthrleuproproserargasp151015gluleuthrlysasnglnvalserleuthrcysleuvallysglyphe202530tyrproseraspilealavalglutrpgluserasnglyglnproglu354045asnasntyrlysthrthrproprovalleuaspseraspglyserphe505560pheleutyrserlysleuthrvalasplysserargtrpglnglngly65707580asnvalphesercysservalmethisglualaleuhisasnhistyr859095thrglnlysserleuserleuserproglylys100105<210>48<211>2055<212>dna<213>人工序列<220><223>构建体<400>48atggagtttgggctgagctgggttttccttgttgctattataaaaggtgtccagtgtcag60gtgcagctggtggagtctgggggaggcttggtcaagcctggagggtccctgagactctcc120tgtgcagcctctggattcaccttcagtgaccactacatgagctggatccgccaggctcca180gggaaggggctggagtgggtttcatacattagtagtagtggtagtatcatatactacgca240gactctgtgaagggccgattcaccatctccagggacaacgccaagaactcactgtatctg300caaatgaacagcctgagagccgaggacacggccgtgtattactgtgcgagaggcgtgccg360gcgaccgatagctggggccagggaaccctggtcaccgtctcctcaggccagcccaagtct420tcgccatcagtcaccctgtttccaccttcctctgaagagctcgagactaacaaggccaca480ctggtgtgtacgatcactgatttctacccaggtgtggtgacagtggactggaaggtagat540ggtacccctgtcactcagggtatggagacaacccagccttccaaacagagcaacaacaag600tacatggctagcagctacctgaccctgacagcaagagcatgggaaaggcatagcagttac660agctgccaggtcactcatgaaggtcacactgtggagaagagtttgtcccgtgctgactgt720tccggtggtggaggatccggtggagggggatcaggaggaggcggtagtgggggaggtggc780agtggcggtgggggatctggaggaggtgggtctggtggcggcgggtcaggcgggggcggg840tcaggaggggggggttctggaggtggagggagcgctaaaacgacacccccatctgtctat900ccactggcccctggatctgctgcccaaactaactccatggtgaccctgggatgcctggtc960aagggctatttccctgagccagtgacagtgacctggaactctggatccctgtccagcggt1020gtgcacaccttcccagctgtcctgcagtctgacctctacactctgagcagctcagtgact1080gtcccctccagcacctggcccagccagaccgtcacctgcaacgttgcccacccggccagc1140agcaccaaggtggacaagaaaattgtgcccagggattgtggttgtaagccttgcatatgt1200acagtcccagaagtatcatctgtcttcatcttccccccaaagcccaaggatgtgctcacc1260attactctgactcctaaggtcacgtgtgttgtggtagacatcagcaaggatgatcccgag1320gtccagttcagctggtttgtagatgatgtggaggtgcacacagctcagacgaaaccccgg1380gaggagcagatcaacagcactttccgttcagtcagtgaacttcccatcatgcaccaggac1440tggctcaatggcaaggagttcaaatgcagggtcaacagtgcagctttccctgcccccatc1500gagaaaaccatctccaaaaccaaaggcagaccgaaggctccacaggtgtacaccattcca1560cctcccaaggagcagatggccaaggataaagtcagtctgacctgcatgataacaaacttc1620ttccctgaagacattactgtggagtggcagtggaatgggcagccagcggagaactacaag1680aacactcagcccatcatggacacagatggctcttacttcgtctacagcaagctcaatgtg1740cagaagagcaactgggaggcaggaaatactttcacctgctctgtgttacatgagggcctg1800cacaaccaccatactgagaagagcctctcccactctcctgggctgcaactggacgagacc1860tgtgctgaggcccaggacggggagctggacgggctctggacgaccatcaccatcttcatc1920agcctcttcctgctcagtgtgtgctacagcgctgctgtcacactcttcaaggtaaagtgg1980atcttctcctcggtggtggagctgaagcagacactggttcctgaatacaagaacatgatt2040gggcaagcaccctag2055<210>49<211>684<212>prt<213>人工序列<220><223>构建体<400>49metglupheglyleusertrpvalpheleuvalalaileilelysgly151015valglncysglnvalglnleuvalgluserglyglyglyleuvallys202530proglyglyserleuargleusercysalaalaserglyphethrphe354045serasphistyrmetsertrpileargglnalaproglylysglyleu505560glutrpvalsertyrileserserserglyserileiletyrtyrala65707580aspservallysglyargphethrileserargaspasnalalysasn859095serleutyrleuglnmetasnserleuargalagluaspthralaval100105110tyrtyrcysalaargglyvalproalathraspsertrpglyglngly115120125thrleuvalthrvalserserglyglnprolysserserproserval130135140thrleupheproproserserglugluleugluthrasnlysalathr145150155160leuvalcysthrilethraspphetyrproglyvalvalthrvalasp165170175trplysvalaspglythrprovalthrglnglymetgluthrthrgln180185190proserlysglnserasnasnlystyrmetalasersertyrleuthr195200205leuthralaargalatrpgluarghissersertyrsercysglnval210215220thrhisgluglyhisthrvalglulysserleuserargalaaspcys225230235240serglyglyglyglyserglyglyglyglyserglyglyglyglyser245250255glyglyglyglyserglyglyglyglyserglyglyglyglysergly260265270glyglyglyserglyglyglyglyserglyglyglyglyserglygly275280285glyglyseralalysthrthrproproservaltyrproleualapro290295300glyseralaalaglnthrasnsermetvalthrleuglycysleuval305310315320lysglytyrpheprogluprovalthrvalthrtrpasnserglyser325330335leuserserglyvalhisthrpheproalavalleuglnseraspleu340345350tyrthrleuserserservalthrvalproserserthrtrpproser355360365glnthrvalthrcysasnvalalahisproalaserserthrlysval370375380asplyslysilevalproargaspcysglycyslysprocysilecys385390395400thrvalprogluvalserservalpheilepheproprolysprolys405410415aspvalleuthrilethrleuthrprolysvalthrcysvalvalval420425430aspileserlysaspaspprogluvalglnphesertrpphevalasp435440445aspvalgluvalhisthralaglnthrlysproargglugluglnile450455460asnserthrpheargservalsergluleuproilemethisglnasp465470475480trpleuasnglylysgluphelyscysargvalasnseralaalaphe485490495proalaproileglulysthrileserlysthrlysglyargprolys500505510alaproglnvaltyrthrileproproprolysgluglnmetalalys515520525asplysvalserleuthrcysmetilethrasnphepheprogluasp530535540ilethrvalglutrpglntrpasnglyglnproalagluasntyrlys545550555560asnthrglnproilemetaspthraspglysertyrphevaltyrser565570575lysleuasnvalglnlysserasntrpglualaglyasnthrphethr580585590cysservalleuhisgluglyleuhisasnhishisthrglulysser595600605leuserhisserproglyleuglnleuaspgluthrcysalagluala610615620glnaspglygluleuaspglyleutrpthrthrilethrilepheile625630635640serleupheleuleuservalcystyrseralaalavalthrleuphe645650655lysvallystrpilepheserservalvalgluleulysglnthrleu660665670valproglutyrlysasnmetileglyglnalapro675680权利要求书(按照条约第19条的修改)1.一种单链vh抗体(scvhab),按照从n末端到c末端:vh-l1-cl-l2-ch1的顺序包含由vh结构域及免疫球蛋白恒定结构域cl和ch1所组成的抗原结合部分,其中l1和l2各自独立地是肽接头;并且其中cl通过β片层接触与ch1配对,从而获得cl/ch1二聚体。2.根据权利要求1所述的scvhab,其中所述cl是cκ或者cλ,优选地其中cλ选自由cλ1、cλ2和cλ3组成的组。3.根据权利要求1或2所述的scvhab,其中l1是长度为3-40个氨基酸的氨基酸序列,优选由下列所组成a)任意组合的甘氨酸和/或丝氨酸的序列;或b)vh框架序列;和/或其中l2是长度为25-50个氨基酸的氨基酸序列,优选地由任意组合的甘氨酸和/或丝氨酸的序列所组成。4.根据权利要求1至3中任一项所述的scvhab,其中所述抗原结合部分的c末端融合到铰链区和其他的免疫球蛋白恒定结构域,所述免疫球蛋白恒定结构域以按照从n末端到c末端的顺序至少包含ch2-ch3。5.一种包含根据权利要求4所述的两个scvhab的重链抗体(hcab),其中使第一scvhab的ch2-ch3结构域与第二scvhab的ch2-ch3结构域配对,从而获得fc区。6.根据权利要求1至4中任一项所述的scvhab或者根据权利要求5所述的hcab,其用于医学用途。7.一种核酸分子,其编码根据权利要求1至4中任一项所述的scvhab。8.一种核酸分子,其编码根据权利要求5所述的hcab。9.一种包含根据权利要求1至4中任一项所述的scvhab或者根据权利要求5所述的hcab的抗体库,包含多种抗体,各自以不同的抗原结合位点为特征,其中所述抗体库通过从b细胞中克隆编码抗体的基因或通过由多种哺乳动物浆细胞分泌抗体来获得。10.根据权利要求9所述的抗体库,其中所述浆细胞具有啮齿动物来源。11.根据权利要求10所述的抗体库,其中所述浆细胞具有小鼠来源。12.一种免疫球蛋白重链基因座,包含:a)可变重链区,其包含每个vh、dh和jh基因片段中的一个或多个,b)恒定重链区,包含编码所述cl和ch1结构域的恒定外显子,任选地进一步包含编码ch2和ch3结构域的外显子,和c)连接区,对这些区域进行工程化并定位成表达根据权利要求1至4中任一项所述的scvhab或根据权利要求5所述的hcab。13.一种转基因非人动物,包含根据权利要求10所述的基因座。14.根据权利要求13所述的转基因非人动物,其是小鼠、大鼠、兔或鸡。15.根据权利要求13或14所述的转基因非人动物,其包含功能丢失突变,该突变使任何内源性轻链基因座κ或λ中的任一个或两者内的某个基因座或基因失活或缺失。16.一种用于形成转基因非人动物的方法,包括:a)提供非人动物细胞;b)提供一种或多种包含编码根据权利要求1至4中任一项所述的scvhab或者根据权利要求5所述的hcab的外显子的载体;c)将所述一种或多种载体导入所述非人动物细胞中;d)将所述外显子整合到所述非人动物细胞的基因组中,并选择转基因细胞,其中所述外显子已在靶位点处被整合到所述非人动物细胞的细胞基因组中,所述靶位点是在内源性免疫球蛋白重链基因座中,即在所述内源性免疫球蛋白重链基因座中的第一ch1外显子的5’;和e)利用所述转基因细胞形成包含所述转基因细胞的转基因非人动物。17.根据权利要求16所述的方法,其中所述非人动物细胞是胚胎干细胞。18.一种产生抗体的方法,包含:a)在非人动物中表达异源免疫球蛋白重链基因座,该基因座包含i)可变重链区,其包含每个vh、dh和jh基因片段中的一个或多个基因片段,ii)恒定重链区,其包含编码所述cl和ch1结构域的恒定外显子,和iii)连接区,对这些区域进行工程化并定位成表达根据权利要求1至4中任一项所述的scvhab或根据权利要求5所述的hcab,其中所述非人动物不表达内源性κ和/或λ基因座;和b)产生抗体,其是根据权利要求1至4中任一项所述的scvhab或根据权利要求5所述的hcab,或者其至少包含所述scvhab或所述hcab的vh结构域。19.根据权利要求13至15中任一项所述的转基因非人动物的用途,a)用于产生scvhab或hcab抗体或者其包括所述vh结构域的片段;或者b)用于产生scvhab或hcab抗体、或者其包含vh结构域的片段的天然文库、或者编码或表达所述天然文库的核酸序列的文库。20.根据权利要求19所述的用途,其进一步包含产生抗体或其片段,所述抗体包含包括在所述scvhab或hcab抗体中的所述vh结构域。当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1