一种抗真菌肽突变体及其制备方法和应用
1.本发明属于生物技术领域,涉及一种抗真菌肽突变体,本发明还涉及该抗真菌肽突变体的制备方法和应用。
背景技术:
2.自从1928年弗莱明发现青霉素,人类进入了抗生素时代。抗生素被用于临床治疗病菌感染,已挽救了亿万人的生命。然而随着抗生素的广泛使用甚至误用,耐药性病菌逐渐显现,使得抗生素的有效性下降,抗生素抗性问题逐渐成为全球公众健康的重要威胁。每年,耐药性感染在全球范围内造成约70万人死亡。估测到2050年,细菌耐药性每年会造成全球1000万人死亡。我国是最大的抗生素生产国和消费国之一,仅2013年,中国抗生素总用量约为16.2万吨,其中48%为人用抗生素,其余为兽用抗生素。滥用抗生素使得我国的抗生素抗性问题日益严重。与此相对的是,新型抗生素的开发速度又滞后于微生物抗性的产生,使得这一现状更为严峻。
3.抗微生物肽(antimicrobial peptides,amps)作为天然免疫的重要效应分子,分布于几乎所有物种中,具有广谱的抗微生物(包括细菌、真菌、病毒和原生生物)活性,此外,还具有免疫调节功能,例如能招募免疫细胞,诱导细胞因子合成与释放,促进伤口修复等。抗微生物肽的作用方式多样,除了能直接与膜相互作用导致内容物泄露或直接裂解细胞外,还可以进入靶细胞,抑制关键生命活动,例如核酸复制,蛋白翻译,酶的活性等,形成代谢抑制效应,最终导致微生物死亡。正是由于这种复杂的作用机制,微生物不易对抗微生物肽产生抗性,这使得其成为药物开发的候选。抗真菌肽为一类具有抗真菌活性的生物活性肽,由包括真菌在内的几乎所有的多细胞生物所产生,参与微生物拮抗和动植物天然免疫防御。这类肽一般含有<100个氨基酸,部分成员含有二硫键用于稳定结构。抗真菌肽可以作为是食品无防腐剂,正在用于开发治疗真菌的药物,用于人和动物疾病的治疗。
4.但是天然形式的抗真菌肽也存在一些不足,限制了其向临床发展,最首要的问题是活性欠佳,及由此带来的高的生产成本和非特异的细胞毒性,另外还包括易受蛋白酶水解。
5.因此,当前对改进活性和性能的抗真菌肽存在需求。
技术实现要素:
6.针对现有技术的不足,本发明的目的是提供一种抗真菌肽突变体,其为线虫来源的抗真菌肽cremycin-5的突变体。相较于野生型肽,本发明获得的突变体具有更广的抗真菌谱和更强的抗真菌活性,更高的热稳定性,更高的血清稳定性,以及更快的杀菌速率。
7.本发明通过如下的发明构思实现,本发明人发现,在进化上,一个蛋白质由保守性和非保守性位点组成。前者因为结构和功能上的重要性,它们的突变往往导致蛋白质生物活性的完全破坏;后者则能够承受广泛的突变而对蛋白质的结构在整体上不造成影响。根据其突变的功能效应,非保守性位点可以划分为三类:1)中性位点,其饱和突变体的功能与
野生型类似;2)可变电阻式位点,其饱和突变体的功能呈现大小可变的行为;3)普遍增强性位点(universally enhanced site,简称ues),其饱和突变体的功能呈现普遍的增强。
8.现有技术中报道了自然界存在一些含普遍增强性位点的蛋白质,例如真菌来源的孢子霉素(第8位谷氨酸),人类抗体igg1(第326位赖氨酸),以及牛胰磷脂酶a2(pla2)(第56位赖氨酸)。本发明人发现,在这些含普遍增强性位点的蛋白中,自然选择没有在ues处选择活性最优的氨基酸残基,而是以次优的氨基酸残基替代。这是因为蛋白质的功能性既是外在的又是内在的。前者是通过与外源的配体(如蛋白,dna和生物膜)相互作用而介导生化反应过程,例如分子识别,结合,催化;而后者则是指蛋白的内部特性,例如稳定性,溶解性,刚柔性和变构性。因此,本发明人认为,从进化的角度,自然选择正是通过引入ues促使内外功能达到平衡,最为典型的例子就是酶活的特异性与热稳定性之间的折衷现象。因此,从进化的角度,我们把普遍增强性位点称为折衷性位点(tradeoff site)。基于折衷性位点是蛋白质的共性,本发明人将其运用到蛋白质工程领域,通过释放这类位点所造成的限制而增强蛋白的活性。
9.基于上述的进化思想,本发明提出了一种新的抗真菌肽设计策略,即首先通过蛋白质并系同源的序列和生物活性比较,确定与该活性相关的氨基酸位点,再利用丙氨酸扫描确定多肽的折衷性位点,再向该位点引入饱和突变,最大限度的释放该位点的限制作用,最终获得一系列活性增强的突变体。该方法的工作量小但成功率高,是一种高效的增强蛋白活性的方法。
10.一方面,本发明提供了一种抗真菌肽突变体的工程化设计的方法,所述方法包括以下步骤:
11.1)确定抗真菌肽的折衷性位点;
12.2)对所述折衷性位点进行饱和定点突变,获得突变体;
13.3)体外抑菌实验分析步骤2)获得的突变体,筛选具有期望的活性或功能的抗真菌肽突变体;
14.其中,所述步骤1)是通过以下方法实现的:
15.①
选择抗真菌肽的并系同源的序列,并进行生物活性比较,从而确定与活性相关的氨基酸位点;
16.②
利用丙氨酸扫描确定多肽的折衷性位点;
17.优选地,所述抗真菌肽选自crem-5野生型肽、果蝇素(drosomycin)、昆虫防御肽(insect defensin)、天蚕素(cecropin)、刺肩蝽素(thanatin)和棉铃虫素(heliomicin)。
18.另一方面,本发明提供了一种抗真菌肽突变体,所述突变体为crem-5野生型肽的突变体,所述突变体包含如seq id no:1所示的氨基酸序列、其生物活性功能片段、变体或衍生物;
19.seq id no:1dvksghykgpcyhdxncngvcrdegyksghcsrwggacwcdt。
20.其中,所述x为lys、arg、his、met、asn、tyr、phe、gln、ser、thr、ala、ile、val、pro、d-lys、d-arg、d-his、d-met、d-asn、d-tyr、d-phe、d-gln、d-ser、d-thr、d-ala、d-ile、d-val或d-pro;
21.优选地,所述x为赖氨酸。
22.优选地,所述抗真菌肽突变体还包括一种或多种低免疫原性或无免疫原性的聚合
物的修饰,从而获得一种低/无免疫原性的聚合物-抗真菌肽突变体。
23.所述无免疫原性的聚合物可以是天然来源的,优选地为明胶或葡聚糖;也可以是人工合成的,优选地为聚乙二醇。
24.在本发明的实施方案中,所述聚乙二醇的分子量为从约5000到40,000优选从约10,000到40,000,最优选地为20,000。
25.上述聚合物可以通过连接基团与抗真菌肽突变体结合,例如乙酰化、甲基化和磷酸化等。所述连接基团可以是任何生物相容性基团,包括但不限于:酯基、醛基、酰胺基、氨基甲酸酯基、顺丁烯二酰亚胺基团或羟琥珀酰亚胺基团。在本发明的实施方案中,所述生物相容性基团是羟琥珀酰亚胺基团或醛基。
26.优选地,所述突变体具有如seq id no:2-15中任一项所示的氨基酸序列:
27.seq id no:2dvksghykgpcyhdkncngvcrdegyksghcsrwggacwcdt。
28.seq id no:3dvksghykgpcyhdrncngvcrdegyksghcsrwggacwcdt。
29.seq id no:4dvksghykgpcyhdhncngvcrdegyksghcsrwggacwcdt。
30.seq id no:5dvksghykgpcyhdmncngvcrdegyksghcsrwggacwcdt。
31.seq id no:6dvksghykgpcyhdnncngvcrdegyksghcsrwggacwcdt。
32.seq id no:7dvksghykgpcyhdyncngvcrdegyksghcsrwggacwcdt。
33.seq id no:8dvksghykgpcyhdfncngvcrdegyksghcsrwggacwcdt。
34.seq id no:9dvksghykgpcyhdqncngvcrdegyksghcsrwggacwcdt。
35.seq id no:10dvksghykgpcyhdsncngvcrdegyksghcsrwggacwcdt。
36.seq id no:11dvksghykgpcyhdtncngvcrdegyksghcsrwggacwcdt。
37.seq id no:12dvksghykgpcyhdancngvcrdegyksghcsrwggacwcdt。
38.seq id no:13dvksghykgpcyhdincngvcrdegyksghcsrwggacwcdt。
39.seq id no:14dvksghykgpcyhdvncngvcrdegyksghcsrwggacwcdt。
40.seq id no:15dvksghykgpcyhdpncngvcrdegyksghcsrwggacwcdt。
41.还一方面,本发明提供一种编码本发明所述的抗真菌肽突变体的前体或编码本发明所述的抗真菌肽突变体的dna序列。
42.再一方面,本发明提供一种含有所述dna序列的表达载体。
43.另一方面,本发明提供了编码所述的抗真菌肽前体或编码抗真菌肽突变体的dna序列的制备方法,包括体外重组dna技术、dna合成技术、体内重组技术。所述dna序列可有效连接到表达载体中的适当启动子上,以指导mrna合成。所述表达载体可以是本领域熟知的细菌质粒、噬菌体、酵母质粒、植物细胞病毒、哺乳动物细胞病毒如腺病毒、逆转录病毒或其他载体。此外,所述表达载体优选地包含一个或多个选择性标记基因,以提供用于选择转化的宿主细胞的表型性状。本发明实施例具体使用了pgex-4t-1作为载体。
44.还一方面,本发明提供了一种重组的宿主细胞,所述宿主细胞含有所述表达载体、或者在染色体中整合有所述dna序列。
45.所述宿主细胞可以是原核细胞,如细菌细胞;或是低等真核细胞,如酵母细胞;或是高等真核细胞,如哺乳动物细胞。代表性例子有:大肠杆菌,链霉菌属;真菌细胞如酵母;cho等。用重组表达载体转化宿主细胞可用本领域技术人员熟知的常规技术进行。当宿主为原核生物如大肠杆菌时,能吸收dna的感受态细胞可在指数生长期后收获,用如cacl2等法
处理,所用的步骤在本领域众所周知。如果需要,转化也可用电穿孔的方法进行。当宿主是真核生物,可选用如下的dna转染方法:磷酸钙共沉淀法,常规机械方法如显微注射、电穿孔、脂质体包装等。本发明实施例具体列举了大肠杆菌作为宿主细胞。
46.还一方面,本发明提供了一种制备所述抗真菌肽的方法,该方法包括:
47.1)用含有编码抗真菌肽突变体的dna序列的重组表达载体转化或转导合适的宿主细胞;
48.2)在合适的培养基中培养宿主细胞;
49.3)从培养基或细胞中分离、纯化蛋白质。
50.宿主细胞可以用常规方法培养表达本发明的抗真菌肽突变体。根据所用的宿主细胞,培养中所用的培养基可选自各种常规培养基。在适于宿主细胞生长的条件下进行培养。当宿主细胞生长到适当的细胞密度后,用合适的方法诱导选择的启动子,将细胞再培养一段时间。在上面的方法中的重组多肽可在细胞内、或在细胞膜上表达、或分泌到细胞外。如果需要,可利用其物理的、化学的和其它特性通过各种分离方法分离和纯化重组的蛋白。这些方法是本领域技术人员所熟知的。这些方法的例子包括但并不限于:常规的复性处理、用蛋白沉淀剂处理、离心、渗透破菌、分子筛层析、吸附层析、离子交换层析、高效液相层析和其它各种液相层析技术及这些方法的结合。
51.再另一方面,本发明提供了一种抗真菌药物组合物,所述药物组合物含有治疗或预防有效量的本发明的抗真菌肽突变体(修饰或未修饰聚合物)以及药学上可接受的载体或辅料。
52.优选地,所述抗真菌肽突变体的致死浓度为2-20μm,优选地为3-10μm。
53.更优选地,所述载体和/或辅料包括水溶性填充剂、ph调节剂、稳定剂、注射用水或渗透压调节剂中的一种或多种。
54.优选地,所述水溶性填充剂选自甘露醇、低分子右旋糖酐、山梨醇、聚乙二醇、葡萄糖、乳糖或半乳糖等中的一种或多种;所述ph调节剂选自枸橼酸、磷酸、乳酸、酒石酸、盐酸等有机或无机酸,以及氢氧化钾、氢氧化钠、氢氧化铵、碳酸钠、碳酸钾、碳酸铵、碳酸氢钾、碳酸氢钠或碳酸氢铵盐等生理上可接受的无机碱或盐中的一种或多种;所述稳定剂选自edta-2na、硫代硫酸钠、焦亚硫酸钠、亚硫酸钠、磷酸氢二钾、碳酸氢钠、碳酸钠、精氨酸、赖氨酸、谷氨酸、天冬氨酸、聚乙二醇、聚乙烯醇、聚乙烯吡咯烷酮、羧基/羟基纤维素或其衍生物如hpc、hpc-sl、hpc-l或hpmc、环糊精、十二烷基硫酸钠或三羟甲基氨基甲烷等中的一种或多种;所述渗透压调节剂为氯化钠和/或氯化钾。
55.又一方面,本发明提供一种本发明所述的抗真菌肽突变体、编码所述抗真菌肽突变体前体或编码本发明所述的抗真菌肽突变体的dna序列、本发明所述的表达载体或宿主细胞或本发明所述的药物组合物在制备用于抗真菌的药物中的应用。
56.与现有技术相比,本发明具有以下优点:
57.1)本发明提供了一种新的改造抗真菌肽活性的方法,相比传统的工程化技术,该方法可快速高效地提高抗真菌肽的活性,同时不仅局限于对抗真菌肽的改造,还可扩展到其他的蛋白质药物活性的改善,具有广阔的应用前景。为快速筛选到折衷性位点,该方法还可以和体外翻译系统相结合以提高效率。
58.2)本发明提供了一种抗真菌肽突变体,与野生型相比,本发明提供的突变体具有
更广的抗真菌谱和更强的抗真菌活性,一些突变体活性提高近10倍。本发明提供的突变体还具有高度的热稳定性,在100c
°
加热5分钟,结构和功能没有改变。本发明提供的突变体还具有高度的血清稳定性。进一步地,本发明提供的突变体不溶小鼠血细胞,并且本发明提供的突变体还具有比临床真菌治疗药物克霉唑更快的杀菌速率。
附图说明
59.以下,结合附图来详细说明本发明的实施方案,其中:
60.图1示出为折衷性位点突变策略流程图。蛋白的氨基酸残基根据进化保守性的不同,可分为保守性位点和非保守性位点。前者因为结构和功能上的重要性,突变通常导致生物活性的破坏。后者的突变具有不同的效应,包括,1)所有的饱和突变体的功能不发生显著的改变;2)所有的饱和突变体的功能呈现可变电阻式的变化;3)几乎所有饱和突变体的活性均增强。首先,利用并系同源蛋白功能发生分化的特点,通过序列-生物活性的比较确定特定功能相关的氨基酸位点,然后对这些位点进行丙氨酸扫描确定折衷性残基(即这里的普遍增强位点),再对折衷性残基进行饱和突变,进而得到一系列增效突变体。
61.图2示出鉴定的crem-5的候选折衷性位点。通过比较crem-5与它的三个并系同源肽(crem-3,crem-9和crem-11)的序列和抗真菌活性关系首先鉴定出crem-5抗真菌活性相关的氨基酸位点。然后对这些位点完成丙氨酸扫描,发现e15a突变体活性增强,为候选的折衷性位点。
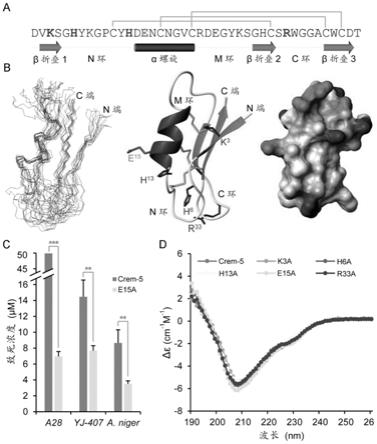
62.图3示出crem-5的氨基酸序列及空间结构以及鉴定的折衷性位点。其中,图a为crem-5的氨基酸序列。粗体表示暴露在分子表面的电荷氨基酸。灰色线示意二硫键的连接方式;蛋白质二级结构元件从解析的实验性结构提取。图b为crem-5的实验性结构。左图表示15个具有最低能量形式的α-碳原子骨架结构的叠加体;中间图表示crem-5空间结构的带状模型;右图表示crem-5空间结构的表面电势图。图c为crem-5及其突变体e15a对构巢曲霉a28,烟曲霉yj-407和黑曲霉的致死浓度。图d为crem-5及其5个突变体的圆二色谱分析。
63.图4示出crem-5折衷性位点突变体的液体抑菌测定及15个突变体的圆二色谱分析。
64.图5示出突变体e15k对白色念珠菌b16的效应测定及治疗潜能评估。其中,图a为突变体e15k对白色念珠菌b16处理后扫描电镜观察结果,箭头指出了胞质分裂异常的细胞。图b为e15k对白色念珠菌b16的杀菌动力学,其中克霉唑和两性霉素b是抗真菌药物,作为阳性对照。图c为e15k的热稳定性,插图显示的是100℃处理5min前后抑菌活性的差异,ns指差异不显著。图d为e15k的血清稳定性。ns指差异不显著。图e为crem-5及其突变体对小鼠血红细胞的溶血测定。
65.图6示出crem-5及其突变体的分子动力学模拟。其中,图a为40ns内crem-5与e15k的α-碳原子的均方根偏差。图b为在20到40ns内crem-5与e15k的α-碳原子的均方根波动。图c为在20到40ns内crem-5与e15k的2001个构象的“香肠模型”。图d为在20到40ns内crem-5与e15k的20个r33构象的叠加。图e为在20到40ns内crem-5与突变体的c11或y12与g36之间氢键的距离。图f为crem-5与e15d的c11或y12上氢原子与g36上氧原子之间的距离。
具体实施方式
66.下面结合具体实施例对本发明作进一步的说明。本实施例仅为解释本发明,不意味以任何方式限制本发明内容。
67.氨基酸缩写的说明:
68.gly:甘氨酸(g)
69.ala:丙氨酸(a)
70.val:缬氨酸(v)
71.leu:亮氨酸(l)
72.phe:苯丙氨酸(f)
73.trp:色氨酸(w)
74.ser:丝氨酸(s)
75.thr:苏氨酸(t)
76.glu:谷氨酸(e)
77.gln:谷氨酰胺(q)
78.asp:天冬氨酸(d)
79.asn:天冬酰胺(n)
80.tyr:苯丙氨酸(y)
81.arg:精氨酸(r)
82.lys:赖氨酸(k)
83.his:组氨酸(h)
84.实施例1使用本发明的方法鉴定抗真菌肽crem-5的折衷性位点
85.crem-5是来源于水果线虫(caenorhabditis remanei)的抗真菌肽,如图3a所示,由42个氨基酸组成,含3对二硫键,灰色线示意二硫键的连接方式,对包括多种丝状真菌及白色念珠菌的临床分离株具有活性。为研究其结构与功能关系,发明人首先通过核磁共振技术解析了其结构,如图3b所示。从实验性结构中提取的二级结构元件被标注在图1a中。图3b左图显示了15个具有最低能量形式的α-碳原子骨架结构的叠加体;图3b中间图显示了由分子动力学模拟得到的平均性结构,以带状模型展示;图3b右图显示了crem-5空间结构的表面电势图。实验性结构证实,crem-5是典型的csαβ折叠形式,即含有一个α-螺旋及三个β折叠,α-螺旋区域是从第14位到第21位的氨基酸残基,三个β折叠分别是从第2到第4位,第29位到第31位以及第38到第41位的氨基酸残基。
86.为寻找crem-5的折衷性位点,发明人按照如下方法进行:
87.1)确定抗真菌肽的折衷性位点;
88.2)对所述折衷性位点进行饱和定点突变,获得突变体;
89.3)体外抑菌实验分析步骤3)获得的突变体,筛选期望活性或功能的抗真菌肽突变体;
90.具体地,如图1和图2所示,通过对crem-5和它的三个并系同源肽(crem-3,crem-9和crem-11)的序列和生物学活性比较发现,k3、h6、h13、e15及r33为crem-5所独有。发明人首先对其进行丙氨酸扫描突变替换。
91.通过pcr策略,使用本领域技术人员已知的方法,获得了5个突变体(k3a,h6a,
h13a,e15a和r33a)。突变体的氨基酸序列如下表1所示:
92.表1序列信息
[0093][0094]
所述序列使用的引物如表2所示:
[0095]
表2使用的引物
[0096][0097]
注:突变的密码子使用粗体表示,并且限制性核酸内切酶位点(bam hi和sal i)使用下划线表示。
[0098]
利用大肠杆菌表达策略,发明人获得了所有重组蛋白,包括有大肠杆菌重组制备的野生型rcrem-5,其中,所述突变体由以下方法制备:
[0099]
1.利用pcr策略,首先构建了pgex-4t-1-crem-5重组表达载体,所用内切酶为bam hi和sal i;
[0100]
2.将测序正确的pgex-4t-1-crem-5质粒转化到大肠杆菌bl21细胞;
[0101]
3.选取过夜培养的单克隆接种到lb培养基(胰蛋白胨10g,酵母提取物5g,nacl 10g),37℃培养至od600=0.6,加入iptg至终浓度0.1mm,诱导4小时;
[0102]
4.离心收集菌体,超声波破菌后,上清由谷胱甘肽琼脂糖凝胶柱进行亲和层析,用5倍体积的pbs缓冲液冲洗杂蛋白;用1-5倍体积的含10mm gsh的tris-hcl洗脱融合蛋白至超滤管中,浓缩蛋白至合适体积。
[0103]
5.采用适量ek酶室温酶切4h,酶切缓冲液采用50mm tris-hcl(ph 8.0),100mm nacl。
[0104]
6.酶切产物用rp-hplc分离纯化。
[0105]
使用maldi-tof测定重组蛋白的分子量,与理论分子量完全吻合,如表3所示。其中,maldi-tof分析在中国科学院微生物所完成。
[0106]
表3 rcrem-5及突变体的分子量
[0107][0108]
通过平板抑菌实验,发明人比较了rcrem-5及其突变体对构巢曲霉a28,烟曲霉yj-407和黑曲霉的活性,如图3c所示。其中,所述平板抑菌实验方法如下:
[0109]
1.真菌平板置30℃培养一周待真菌孢子有较大密度,用灭菌水洗涤收集孢子制成悬液;
[0110]
2.取10μl真菌孢子悬液加入6ml预热的0.8%的pga固体培养基,充分混匀后倒入一个ф90mm平皿中;
[0111]
3.待培养基凝固后,用胶头滴管打孔ф2mm,设置倍比稀释的浓度梯度,每孔2μl上样,30℃培养24小时;
[0112]
4.测量抑菌圈直径,以多肽用量的对数值为自变量,抑菌圈直径的平方为因变量绘制散点图,计算致死浓度c
l
=2.93/(ak10
m/k
),m代表截距intercept,k代表斜率slope,a为打孔的深度为1mm。
[0113]
结果表明,相比于野生型rcrem-5,k3a、h6a、h13a和r33a基本丧失或仅保留微弱的活性(致死浓度高于50μm),而e15a对构巢曲霉a28的活性至少增强了6.19倍,对烟曲霉yj-407的活性增强了0.88倍,对黑曲霉活性增强了1.46倍。
[0114]
使用圆二色谱技术证实了这些突变体的空间结构与野生型高度相似,排除了突变可能对功能造成的影响,如图3d所示。其中,圆二色谱技术在中科院生物物理所完成。cd仪器为chirascanplus spectropolarimeter(uk),测量蛋白质浓度为0.1mg/ml。波长范围190-260nm。石英比色皿光程为1毫米。
[0115]
综上结果证实,α螺旋上的e15为折衷性残基,位于n环上的h6,h13,邻近的k3及c环
上的r33为crem-5的功能性残基。
[0116]
实施例2抗真菌肽crem-5折衷性位点的饱和突变及功能检测
[0117]
为最大程度的提高crem-5的活性,发明人对第15位折衷性位点进行了饱和替代,即除谷氨酸和半胱氨酸之外的其他18种天然氨基酸(e15k、e15r、e15h、e15m、e15n、e15y、e15f、e15q、e15s、e15t、e15a、e15i、e15l、e15v、e15p、e15g、e15w和e15d)。突变体的氨基酸序列如下表4所示:
[0118]
表4序列信息
[0119]
[0120][0121]
所述序列使用的引物如表5所示:
[0122]
表5使用的引物
[0123][0124]
注:突变的密码子使用粗体表示
[0125]
通过大肠杆菌表达了除e15l,e15g和e15w之外的15个突变体。其中,e15l、e15g和e15w由于ek酶的非特异切割或形成包涵体而未成功获得。
[0126]
使用maldi-tof测定了重组蛋白的分子量,与理论分子量吻合,如表6所示。
[0127]
表6 rcrem-5突变体的分子量
[0128][0129]
通过平板抑菌实验,发明人比较了rcrem-5及其突变体对两种丝状真菌和四种白色念珠菌的临床分离株的活性,如表7所示。
[0130]
表7 rcrem-5及其突变体的对真菌的致死浓度
[0131]
[0132][0133]
结果表明,相比于野生型crem-5,14个突变体对所有6种菌株有至少1倍的活性增强。整体来看,40%的突变体(e15k、e15r、e15h、e15m、e15n和e15y)对6种菌株活性增强,绝大多数有至少4倍的提升;33%的突变体(e15f,e15q,e15s,e15t,和e15a)对5种菌株活性增强;13%的突变体(e15i和e15v)对4种菌株活性增强;7%的突变体(e15p)对3种菌株活性增强;只有保守性替代突变体(e15d)对菌株活性未增强。其中,e15k对5种受试菌活性增强在9.5到15.8倍不等。
[0134]
为验证上述测定的致死浓度,发明人对crem-5和突变体e15q,e15n,e15m和e15k做了液体抑菌实验,如图4a所示。其中,所述液体抑菌实验方法如下:
[0135]
1.挑白色念珠菌克隆于pgm培养基,30℃培养,菌液稀释至1
×
103cfu/ml。
[0136]
2.10μl倍比稀释的多肽分别加入90μl稀释后的菌液中,在30℃孵育24小时,样品在pga平板划线以确定没有真菌生长的最小浓度,即为最小抑制浓度mic。
[0137]
液体抑菌结果表明,crem-5和突变体的最小抑制浓度与致死浓度基本一致。e15q,e15n,e15m与e15k相比于crem-5活性也分别增强了1,3,7,和15倍。
[0138]
圆二色谱技术证实了这些突变体的空间结构与野生型高度相似,排除了突变可能对功能造成的影响,如图4b所示。
[0139]
实施例3突变体e15k对白色念珠菌b16的效应测定及治疗潜能评估
[0140]
发明人研究了活性提升最明显的突变体e15k的杀菌效应并评估了治疗潜能,如图5所示。其中,所述研究方法如下:
[0141]
扫描电镜观察:白色念珠菌b16与5倍c
l
浓度的e15k在30℃孵育6小时,2.5%戊二醛固定1小时,然后pbs洗涤三次。细胞经乙醇梯度脱水,然后由临界点干燥仪(leica em cpd300,austria)干燥,再喷金(leica em scd050,austria)处理。由环境电子显微镜fei quanta 450拍摄。
[0142]
杀菌动力学:1
×
106cfu/ml的白色念珠菌b16与5倍c
l
浓度的e15k在30℃孵育6小时。期间,每间隔1小时,取样,稀释涂平板计数。以培养基组和抗真菌药物克霉唑和两性霉素b分别作为阴性和阳性对照。
[0143]
稳定性评估:为评估血清稳定性,e15k分别在水和小鼠血清中37℃孵育0,12和24小时。然后通过平板抑菌试验研究不同孵育条件下e15k对白色念珠菌b16的活性。为评估热稳定性,e15k分别在50℃孵育5天,80或100℃孵育5分钟。用圆二色谱分析不同孵育温度下
e15k的二级结构变化。其中,对100℃孵育5分钟前后进行平板抑菌试验。
[0144]
溶血实验:小鼠眼静脉取血,加入edta,混匀,细胞沉淀用pbs洗涤3遍后加入pbs,制成10%(细胞沉淀体积/总体积)的血细胞悬液;然后加入终浓度为6.25,12.5,25和50μm多肽。30℃静置15min;以pbs为阴性对照a
blank
,加入1%triton x-100(完全溶血)为阳性对照a
tot
,溶血比率为(a
pep-a
blank
)/(a
tot-a
blank
)
×
100%。
[0145]
结果表明,与野生型crem-5类似,e15k也能抑制白色念珠菌的胞质分裂,如图5a所示。e15k的杀菌动力学表明其杀菌速率介于克霉唑(代谢抑制型抗真菌剂)和两性霉素b(破膜型抗真菌剂)之间,如图5b所示。e15k具有很强的热稳定性,甚至在100℃孵育5分钟后,其结构及抑菌活性基本保持不变,如图5c所示。另外e15k具有很好的治疗潜能。如图5d,5e所示,其在小鼠血清中至少能稳定24小时,且在50μm浓度时也基本不溶血。
[0146]
实施例4 crem-5与其突变体的分子动力学模拟
[0147]
为解释折衷性残基的替换能显著增强crem-5的活性,发明人通过分子动力学模拟分析了突变对功能位点动态性的影响。其中,分子动力学软件:gromacs 5.1.4.
[0148]
在40ns内的模拟过程,如图6a所示,e15k的α-碳原子的均方根偏差(rmsd)均低于野生型crem-5,表明e15k的刚性更强。如图6b,6c所示,e15k在三个环区的刚性均较crem-5强。分析分子间作用力发现,e15k c环的第36位甘氨酸能分别与n环上的第11位半胱氨酸及第12位酪氨酸形成氢键,而crem-5在该位置没有氢键形成。这就解释了crem-5与e15k刚性的差异,e15k通过氢键限定了第33位的功能性位点精氨酸的构象,如图6d所示。如图6e所示,这一现象在其他增效突变体中得到进一步证实,增效突变体在n环第11位半胱氨酸及第12位酪氨酸与c环的第36位甘氨酸之间至少能形成一对氢键,但是e15d则与crem-5类似,该位置没有氢键形成。因此,e15位的增效突变体很可能是通过促使在n环第11位半胱氨酸及第12位酪氨酸与c环的第36位甘氨酸之间形成氢键进而限定了第33位的功能性位点精氨酸的构象,使其适合与真菌相互作用,从而增强了抑菌活性。
[0149]
最后说明的是,以上优选实施例仅用以说明本发明的技术方案而非限制,尽管通过上述优选实施例已经对本发明进行了详细的描述,但本领域技术人员应当理解,可以在形式上和细节上对其作出各种各样的改变,而不偏离本发明权利要求书所限定的范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1