一种基于流式细胞术定量分析细胞核内转录因子含量的方法与流程
本发明涉及生物技术领域,具体而言,涉及一种基于流式细胞术定量分析细胞核内转录因子含量的方法。
背景技术:
转录因子(transcriptionfactor)是指能够在细胞核内结合在某基因上游特异性核苷酸序列上的蛋白质,这些蛋白质能调控基因的转录。许多核转录因子在活化的情况下会从细胞质中转移到核中启动转录,这些核转录因子在细胞核中的分布情况可以在一定程度上反映其转录活性。而转录因子的活性与细胞的生长,增殖,应激,死亡等过程密切相关,也参与了人体的各种生理和病理过程。
自噬在细胞内容物降解过程中发挥重要作用,其有助于维持真核细胞的稳态和存活,并且涉及初始形成的自噬体和由溶酶体介导的最终降解。自噬功能障碍与多种病理生理过程相关1-2。在许多疾病状况下,如神经退行性疾病、心血管疾病和感染性疾病中,增强自噬或自噬相关过程显示出有益的作用3。此外,抑制自噬被认为是临床治疗癌症的潜在方法。
tfeb(转录因子eb)是调节自噬和溶酶体生物功能的关键靶标。越来越多的证据支持tfeb在细胞和动物模型中的过表达可以改善神经退行性疾病的发展4-6,减轻动脉粥样硬化7,保护机体免受心脏毒性8,缓解庞贝氏症9和其他溶酶体贮积症中的自噬和溶酶体病变10-11。通常情况下,磷酸化的tfeb(失活形式)分布在细胞质中,去磷酸化的tfeb(活性形式)转位至细胞核,随后启动与自噬和溶酶体生物合成有关的基因的转录12-14。
目前报道的活化tfeb的机制主要涉及抑制活性的蛋白激酶,例如抑制mtorc1(雷帕霉素靶蛋白复合物1)12,15和akt/pkb(蛋白激酶b)16,或降低tfeb磷酸化、优先与无活性磷酸化tfeb17相互作用并泛素化,或与tfebn端直接结合以促进其核转位等机制18。
tfeb是调控与自噬和溶酶体生物功能相关基因转录的关键转录因子。在饥饿或抑制mtorc1等条件下,tfeb从细胞质转移至细胞核以启动自噬过程。目前常规使用的显微镜观测和蛋白印迹法来量化tfeb核转位不可避免地会带来偏差,或仅具有较弱的量化分析能力。
鉴于此,特提出本发明。
技术实现要素:
本发明的目的在于提供一种基于流式细胞术定量分析细胞核内转录因子含量的方法。采用该方法可以对细胞核内的转录因子进行定量分析,其具有高通量和更准确的量化分析特点。
本发明是这样实现的:
一种基于流式细胞术定量分析细胞核内转录因子含量的方法,其包括如下步骤:
细胞核分离步骤:将待检测的细胞进行细胞核分离,得到细胞核;
转录因子染色步骤:用转录因子抗体与所述细胞核孵育,然后用荧光素标记的二抗进行孵育,得到荧光染料标记的细胞核。
流式细胞术分析步骤:采用流式细胞术分析染色后的细胞核的荧光强度。
进一步地,在本发明的一些实施方案中,在细胞核分离步骤中,将待检测的细胞与核质分离试剂混合,经静置、离心后,弃去上清胞质,再往沉淀中加入核质分离试剂,经静置、离心后,弃去上清,获得沉淀,即为所述细胞核。
进一步地,在本发明的一些实施方案中,静置的条件为:温度3-6℃,时间4-6min。
进一步地,在本发明的一些实施方案中,离心的条件为:转速:450-550g,时间4-6min。
进一步地,在本发明的一些实施方案中,在转录因子染色步骤中,在用转录因子抗体孵育前,将得到的所述细胞核用多聚甲醛固定,再用tritonx-100透化,接着用山羊血清进行封闭。
进一步地,在本发明的一些实施方案中,所述多聚甲醛的浓度为3%-5%。
进一步地,在本发明的一些实施方案中,所述tritonx-100的浓度为0.1%-0.3%。
进一步地,在本发明的一些实施方案中,所述山羊血清的浓度为4%-6%。
进一步地,在本发明的一些实施方案中,封闭后,载玻片上的所述细胞核用抗褪色封片剂包埋封片。
进一步地,在本发明的一些实施方案中,所述荧光染料选自异硫氰酸荧光素、四甲基罗丹明-5(6)-异硫氰酸、cy2、cy3、cy5、cy7和alexafluor染料。
需要说明的,tfeb是一种核转录因子,主要调控自噬溶酶体通路相关的基因转录。本发明实施例以tfeb为具体应用例子,来说明本发明提供基于流式细胞术定量分析细胞内转录因子含量或分布情况的方法。基于此,本发明提供的方法可以适用于所有的核转录因子的含量检测以及分布情况检测例如细胞质和细胞核中分布情况检测。
本发明的转录因子包括但不限于tfeb、nf-kappab、keap1、beta-catenin、hgncsymbol、nfya、arx、hoxa11、sox8、zfx、dlx6、etv1、pax6、tead3、e2f2、tfap2b、tfap2d、nfix、pax7、mlxipl、etv7、znf200、hivep2、nr1h4、isl1、snai2、runx3、hsf2、nr1h3、pou2f2、arntl2、nr2e3、hoxc8、sox30、barx2、zic2、nfe2l3、alx4、foxc1、prdm1、mxd1、csda、gsc2、dlx3、dmrt3、mef2b、pou1f1、znf76、foxj2、ybx1、nfyc、spi1、klf6、zfy、cbfb、mef2a、tfe3、pitx1、rora、hes2、mnt、tcf3、srebf1、nfatc3、tp63、tbx21、gli2、tead2、znf37a、fosl2、pax2、rarb、nfkb2、hoxa9、tp73、cic、rfx3、tcf7、mef2c、pgr、nfe2l1、grhl2、rest、nfx1、tfap2c、rfx2、tbx5、rcor1、tfap4、znf10、pou4f3、esr1、cebpe、crem、hivep1、cdc5l、mzf1、sox10、xbp1、six4、hif1a、yy1、nfatc4、vsx1、mybl2、hnf4a、nfatc2、tcfl5、gmeb2、e2f1、elf4、gata1、klf8、klf5、tsc22d1、hsf4、nfat5、ctcf、znf174、sall1、maz、trps1、relb、lyl1、crx、meis3、znf175、dbp、klf1、etv2、usf2、znf14、erf、hbp1、sp4、dlx5、tfec、hoxa1、hoxa2、hoxa3、hoxa5、hoxa6、hoxa13、pax4、ezh2、meox2、pou6f2、gli3、lhx2、lhx6、creb3、lhx3、gata3、tlx1、pitx3、ebf3、tfam、ubtf、hoxb6、hnf1b、mlx、ezh1、dlx4、foxn1、phox2b、nfkb1、elf2、whsc1、sox6、prdm4、myf6、myf5、gli1、elk3、foxm1、cux2、crebl2、vdr、nanog、ppard、bach2、znf451、e2f3、nr2e1、tfeb、srf、hand1、irx4、nr3c1、smad5、cdx1、bcl6、tfdp2、hes1、foxp1、tfcp2l1、tlx2、stat1、otx1、id2、cebpz、dlx2、atf2、epas1、vax2、nfe2l2、prrx1、ash1l、mef2d、prdm2、zbtb17、tfap2e、nr5a2、etv3、id3、irf6、prox1、atf6、creb1、plagl1、myb、tcf21、klf12、satb2、klf9、nr4a3、onecut2、prox2、vsx2、esrrb、znf410、nkx2-3、mxi1、hoxb8、hoxb5、hoxb7、hoxb3、hoxb1、msx2、elf1、smad9、egr1、nr2c1、nfyb、tbx2、tbx4、tshz3、lhx4、myog、znf205、znf644、hoxa7、twist1、neurog3、egr2、atf1、nr4a1、hoxc13、hoxc11、nfe2、hoxc12、mxd4、ctcfl、znfx1、snai1、rbpjl、hif3a、trerf1、spdef、sox4、rreb1、runx2、gcm2、sox21、irf1、sox9、barhl1、bhlhb4、pax8、fosb、foxa2、pax1、tcf15、max、id1、plagl2、thra、nr1d1、irf3、stat5a、dach2、znf384、elk1、six1、prdm7、hivep3、bcl11b、klf2、atf4、foxp2、irf5、hoxd1、hoxd3、hoxd9、hoxd10、hoxd11、hoxd13、myod1、sox15、foxa1、nrl、klf16、jund、gata5、npas1、znf236、nfatc1、cdx4、nr1h2、barx1、rara、rfx1、pparg、e2f5、arntl、dpf2、gsc、bhlhb2、meis2、grhl1、mycn、rax、sox5、sox3、clock、ets1、hnf1a、tbx3、dmtf1、ehf、elf5、tfcp2、hey2、pax3、nkx2-1、gata4、skil、hlx、klf4、nr5a1、lmx1b、myc、dmrt1、tfap2a、irf4、gcm1、crebzf、pou2f3、six3、lbx1、stat4、prdm5、lef1、etv6、ascl1、pdx1、esr2、tcf12、zfhx3、irf8、gata6、tp53、foxk2、smad4、nfic、spib、znf593、barhl2、rxrg、tbx19、pou2f1、mael、nr1i3、lhx9、rorc、rfx5、arnt、creb3l4、sox13、meis1、dlx1、nr1i2、rbak、creb5、znf157、znf182、st18、znf7、nfib、nr6a1、zeb1、vax1、tcf7l2、nkx6-2、znf215、tbx6、thrb、pou4f2、nr3c2、fli1、pou4f1、tcf7l1、hhex、zic1、nr4a2、ubp1、tgif2lx、hoxa10、jazf1、cebpg、gabpa、klf10、batf、alx3、bach1、znf19、tsc22d3、erg、ets2、creb3l1、ski、grhl3、elk4、usf1、hoxb13、runx1、irx6、isl2、znf230、nr2f6、pknox1、aire、zbtb7b、foxh1、ikzf3、znf385a、bcl6b、gmeb1、nfia、znf281、lmx1a、atf3、rel、neurod1、znf2、msx1、elf3、fev、eomes、nkx6-1、hesx1、znf148、klf15、heyl、pitx2、hand2、ebf1、tlx3、t、neurod6、znf12、hey1、sox17、hnf4g、zhx1、phox2a、znf22、cdx2、otx2、sohlh1、dach1、snapc4、gfi1b、sall2、spic、znf202、znf143、znf3、stat6、smad3、nkx3-1、tef、pbx3、prrx2、sp2、znf91、znf23、znf180、znf146、znf444、rcor2、tbx10、e4f1、batf2、rbpj、ptf1a、foxi1、irf2、stat3、znf30、shox2、sox14、e2f6、ar、atf5、nr0b1、zeb2、bnc1、hic2、znf32、gsx1、onecut1、klf13、zfpm2、znf35、hoxd4、hoxd12、znf282、fos、smad1、emx2、sp7、npas2、irx1、irx2、six2、stat2、foxa3、scrt1、znf16、atf7、hoxb9、foxn2、znf160、znf350、sox7、junb、neurod2、znf274、etv5、foxb1、klf11、cebpb、atoh1、mizf、arnt2、znf24、hoxc5、rarg、sp3、rela、esrra、dmrt2、insm1、znf622、stat5b、hoxb2、zbtb4、zhx3、glis1、nr1d2、ddit3、prop1、smad2、znf25、fosl1、mlxip、nr2f1、etv4、hoxd8、ascl3、hsf5、foxg1、zscan2、tgif2ly、sox11、deaf1、six5、zbtb38、hic1、tgif1、nr2c2、zbtb33、irx3、jun、sox12、znf260、lcorl、maf、zhx2、msc、foxe1、zbtb7a、hes7、gata2、egr3、znf238、zfpm1、atoh7、alx1、bhlhb8、gsx2、hoxc9、hoxc10、bhlhb5、znf322a、sox2、plag1、neurog1、znf708、creb3l2、satb1、hoxb4、mafa、sox1、nkx2-5、figla、ascl2、tbx1、olig1、pou6f1、six6、foxo4、pou3f2、sry、wt1、maff、hsf1、mixl1、irf7、nr2f2、sp1、pbx1、pou3f1、mybl1、ikzf1、znf267、shox、rxra、znf197、znf396、znf395、ppara、tead1、mitf、foxd3、ascl4、mesp2、lbxcor1、mtf1、znf322b、sp6、pax5、myt1、lcor、esrrg、tcf4、pou3f4、znf398、mafg、hoxa4、hoxc6、znf181、znf44、tead4、znf273、zfp161、mier1、tfdp1、hoxc4、znf26、mafk、znf43、znf28、yy2、pax9、srebf2、pou3f3、rorb、sox18、mycl2、mafb、rxrb、pbx2、sp5、znf551、pou5f1、zbtb48、e2f4、olig2、、znf254、mxd3、crebl1、irf9、noto、zbed1、hmx1、、meox1、znf195、znf263、znf207、dpf1、wiz、zbtb32、znf221、znf582、zfp64、znf839、phf20、ikzf2、zzz3、foxp3、foxn3、znf280c、creb3l3、prdm6、zfp112、znf275、zbtb11、zfat、zxdc、znf532、znf510、znf93、znf324、znf671、znf416、znf586、znf446、znf8、znf264、mynn、evi1、znf213、znf343、ebf4、lhx5、znf302、zbtb25、zfhx4、tbx15、ikzf5、znf184、patz1、zbed4、sall4、adnp、znf516、adnp2、rhoxf1、znf629、znf423、znf500、foxf1、znf90、znf550、znf419、pbx4、znf574、znf85、evx1、zkscan1、nobox、glis3、camta2、hlf、nkx3-2、klf3、dbx1、zbtb16、rfx4、prdm13、zbtb24、tbx18、bbx、zbtb47、znf142、gon4l、mycl1、rlf、znf684、dr1、rcor3、znf541、klf7、znf271、znf430、foxo3、zbtb45、bcl11a、zfp30、znf706、znf549、zscan18、znf211、znf639、zscan20、tbx22、bhlhb3、neurod4、ikzf4、esx1、batf3、znf137、znf576、znf45、gzf1、nkx2-4、nkx2-2、znf133、ovol2、znf436、glis2、sebox、zbtb1、znf484、foxj1、zscan10、mllt1、znf557、zbtb46、mnx1、znf337、prdm12、znf317、znf426、znf331、casz1、znf341、znf227、znf141、rhoxf2、znf304、zscan5a、znf132、znf20、lhx1、toe1、znf414、egr4、emx1、znf670、sp110、nkx2-8、zfhx2、vezf1、tbr1、zfp37、znf189、foxp4、znf193、foxf2、znf280d、znf365、aebp2、foxn4、znf740、zic5、jdp2、zscan29、znf710、znf434、znf287、znf750、zim3、prdm15、dmrtc2、zfp14、znf473、znf432、rere、prdm16、dmrta2、dmrtb1、znf697、znf687、osr1、znf514、znf385b、znf660、znf300、znf92、znf41、znf711、hmbox1、prdm14、znf462、ciz1、znf214、mkx、foxo1、sall3、ventx、znf385d、znf268、znf547、znf256、zscan1、zfp36l2、prdm8、znf117、znf599、lass3、znf18、mier3、zkscan2、c14orf43、znf689、zic3、znf618、zscan12、znf276、znf222、znf225、znf234、znf233、znf235、znf486、znf208、znf761、znf714、znf394、znf333、cux1、znf382、znf577、znf653、znf75a、tal1、lhx8、znf326、gfi1、znf496、en1、znf513、znf691、znf589、znf474、prdm9、fam170a、foxq1、tbx20、sp8、znf704、en2、uncx、gbx1、osr2、nfil3、hdx、znf488、pknox2、drgx、znf503、znf219、znf319、zscan21、znf606、znf592、zbtb39、tsc22d4、znf226、znf229、znf668、znf646、znf641、znf610、znf701、znf526、znf345、znf83、znf558、znf232、znf597、insm2、gbx2、zbtb5、znf507、znf509、znf608、znf354a、zbtb43、znf280a、znf747、foxd4、znf212、prdm10、znf296、znf415、znf672、znf692、znf439、znf440、znf581、znf524、zbtb26、znf562、znf318、znf561、otp、znf584、zik1、camta1、nhlh1、znf540、znf570、mllt3、klf17、znf217、znf57、znf556、znf554、etv3l、znf131、zmat3、znf596、ovol1、znf621、znf680、bnc2、znf483、znf449、znf295、znf417、rax2、mga、znf80、evx2、znf497、klf14、znf266、zic4、znf654、znf408、znf519、isx、drap1、znf77、znf169、znf645、znf611、znf613、znf683、znf404、znf283、znf135、foxr1、dmrta1、kcmf1、znf575、foxl1、foxc2、irx5、zbtb34、ccdc79、olig3、nhlh2、znf491、znf620、znf518a、znf619、zbtb41、znf354c、znf114、znf366、znf454、znf543、znf354b、znf223、znf713、tprx1、znf552、zfp42、znf664、lbx2、foxs1、znf154、znf648、fiz1、znf771、tshz1、znf48、nkx2-6、znf609、znf571、zscan4、zfp3、znf443、znf572、zfp82、znf707、znf746、znf467、znf678、zbtb2、zfp41、hkr1、zbtb20、rfx7、znf329、znf101、zscan22、tshz2、znf721、znf662、znf320、znf623、tfdp3、znf438、znf530、foxl2、znf703、znf285a、foxd4l1、zfp1、znf56、zbtb40、zbtb7c、znf479、znf445、znf74、zbtb37、dbx2、zfp36l1、snai3、zbtb3、znf696、znf566、znf224、znf529、znf284、tal2、argfx、zbtb6、znf749、znf17、znf555、znf75d、znf501、myt1l、foxd2、zfp91、foxe3、znf546、foxd4l3、znf286a、zkscan4、znf70、znf643、znf642、zscan23、znf490、znf626、znf383、hes4、znf669、znf559、hmx3、znf177、znf548、znf563、znf292、znf567、znf573、znf527、znf33a、znf600、zkscan3、foxr2、znf699、znf250、znf79、znf681、znf107、znf471、znf493、zbtb44、znf167、znf565、znf34、znf140、znf124、tsc22d2、znf569、znf777、znf605、znf136、zkscan5、znf502、zfp62、znf33b、znf431、znf418、znf700、znf239、zscan16、znf709、zfp28、znf252、znf705a、znf585a、znf138、znf470、znf100、znf542、znf498、znf441、znf420、znf187、znf679、znf682、znf257、znf688、znf165、znf655、znf517、znf675、znf695、znf628、znf665、znf624、dmbx1、znf615、znf433、znf323、znf595、znf460、znf81、znf585b、znf461、hes5、znf677、znf823、znf311、znf347、znf71、znf121、znf734、znf335、znf560、znf84、znf667、znf649、znf248、znf544、znf251、znf607、znf334、zxda、znf485、zim2、znf192、znf442、znf69、znf583、znf568、zxdb、znf480、znf587、znf280b、znf512、znf511、znf627、znf658、znf536、znf652、znf830、znf521、foxj3、znf358、znf277、zfp2、、zbtb22、zbtb12、duxa、dprx、znf468、znf616、zfp57、foxd4l6、znf155、znf783、znf425、zbtb10、onecut3、znf409、、znf580、znf625、zbtb9、znf134、leutx、znf99、znf253、、homez、znf705f、znf705d、znf718、scrt2、znf407、znf663、、znf579、znf564、bclaf1、tcf20、gabpb1、nrf1、ahr、sim1、bmp2、preb、tcf25、aff3、arid5b、cxxc1、sim2、grlf1、ifi16、tub、npas4、pawr、cebpa、nkrf、rfxank、arid3a、tcf19、pcgf6、nfrkb、phb2、cramp1l、zc3h3、slc30a9、matr3、thap3、znf800、rc3h2、zc3h11a、unkl、zc3h15、sp100、zfyve26、aff4、mbnl3、zcchc6、sohlh2、ncoa1、c20orf194、tox4、cenpt、znf821、tox3、zfp106、dnajc2、tax1bp1、akna、znf346、plek、mll、kiaa1549、ncoa3、tox2、znf831、znf391、c11orf9、znf835、znf780b、fezf1、thap10、e2f8、c14orf106、znf428、thap1、terf2、zbed3、zcchc11、hmga1、prkrir、scaper、znf787、zmat2、ferd3l、cpxcr1、terf1、npas3、msgn1、znf827、setbp1、znf773、znf776、znf837、carhsp1、ahctf1、fezf2、zufsp、gatad1、znf362、thap8、mysm1、bola3、ankzf1、atxn7、tcf23、yeats2、c5orf41、foxk1、kiaa0415、zmat4、nkx6-3、znf367、e2f7、mesp1、mll2、znf598、znf834、thap9、thap11、dnajc21、atoh8、znf764、znf768、znf778、znf804a、nfxl1、id4、lass6、aff1、noc3l、thap2、hes3、znf791、thap6、kiaa2018、znf843、thap4、zbed2、thap5、bola1、znf518b、neurog2、znf784、znf725、znf321、znf594、znf487、znf792、znf716、znf804b、thap7、noc4l、znf733、znf717、znf829、foxi2、znf385c、tcf24、helt、znf793、znf788、hmx2、bsx、zfp92、znf676、znf766、znf836、znf781、znf774、znf775、znf799、znf782、znf846、znf429、znf763、znf772、zscan5b、znf720、znf833、znf98、znf786、znf841、znf780a、znf790、znf770、znf506、znf808、wdhd1、znf789、znf828、zscan5c、、znf316、znf845、、、rbm5、plxnd1、rbm6、dvl2、ybx2、csde1、akap8l、depdc1、brd9、hmgb3、depdc1b、cul3、fam44a、trit1、depdc2、rad51、cul1、mll3、zfr、mta3、riok2、parp12、pms1、hmg20b、mbd3、smarce1、wnt8b、znf638、mkrn2、baz2a、rgs11、plxna2、ube2k、sp140、rapgef3、srcap、dusp12、myst3、nufip1、pds5b、ssh1、mrpl28、rbm22、lass4、rbm27、rapgef4、dsp、depdc5、dmc1、hmg2l1、zmat5、zc3h7b、plek2、zc3h14、smad7、rnf125、smarca1、eea1、fgd1、myst1、chrac1、ppp1r13l、dot1l、sf3a2、trmt1、akap8、zfr2、mier2、plekha4、cbll1、zc3hav1、met、edf1、dvl1、larp5、cul2、tada2l、rgs9、psmd11、trim3、exoc2、grm6、trim23、plxna1、pip5k3、pole4、farsb、rpa2、casp8ap2、tgif2、trim32、c1orf25、depdc7、gtf3a、zc3h7a、zc3h13、nrk、baz2b、hmg1l1、chd6、rnf114、zbp1、hist1h1d、hist1h1a、dek、rnf113a、ttf1、slc2a4rg、gtf2f1、cenpb、prr12、ahdc1、yeats4、ubr4、hp1bp3、macf1、zfp36、mbd4、kiaa1683、zc3h4、plxna3、zfyve20、nr0b2、lin28、unk、mkrn1、mbd2、prb4、rnf138、dhx34、dzip1、trafd1、cd36、phf21a、zc3h10、slc26a10、rc3h1、plxnc1、setdb2、rapgef5、nupl2、myst2、dnajc1、xpa、aptx、smad6、ssb、cops4、larp2、smarcc2、lass5、rbm26、mbnl2、rnf113b、cul4a、hmg20a、ncoa2、adamts17、pcsk6、tigd7、ncor1、cops3、ssh2、mink1、mbd1、dus3l、pogk、setdb1、lass2、pogz、gatad2b、zc3h8、slc4a10、fam171b、hes6、bdp1、gtf2h2、adamts19、scml4、shprh、mrrf、pole3、tut1、ssrp1、mta2、hmga2、kin、mbnl1、smarca5、chd1、sorbs2、larp1、depdc6、myst4、brpf1、fmnl2、dzip1l、cul4b、zc3h18、rnf166、u2af1、adar、cpsf4、dvl3、u2af1l4、larp4、c17orf49、bpnt1、fbxo41、vps72、dhx57、gpr155、tiparp、pbrm1、zdhhc19、plxnb1、mst1r、hmgb2、tigd6、rell2、fbn1、depdc4、larp6、cops2、cul5、c12orf28、rag1、zmat1、atmin、terf2ip、ppp2r3b、gatad2a、znf528、hist1h1e、usp39、mecp2、zrsr2、rnase2、tigd4、mgmt、pkhd1、chd7、hopx、csdc2、znf738、ssh3、kat5、smarcc1、chd2、tigd3、cbx2、gtf2ird2b、larp7、znf674、anapc2、hmgb4、chd9、ccdc71、polr2l、dmap1、znf826、suz12、h1foo、eif3k、farsa、hla-dqb1、mkrn3、tigd5、tigd2、c10orf140、yod1、kiaa1618、tceal8、rgs6、rbm10、rgs7、leng9、otop3、ewsr1、mta1、jrkl、sf3a3、ep400、hist1h1b、kntc1、h1fx、zfp90、rfx6、prmt3、gpatch8、hist1h1t、lin28b、hist1h1c、cpsf4l、zc3h6、gtf2f2、hils1、zdhhc11、h1f0、arid2、hmgb1、plxna4、sema4a、gtf2ird2、ncor2、plxnb2、znf512b、slc39a10、zgpat、psmd12、gpr123、slc22a4、gtf2e2、prb3、helz、znf765、plxnb3、tox、krtap5-9、inf2、rbm20、rhoxf2b、foxo6、rpa4、hla-dqb2、znf805、ppp1r10、prr3、foxb2、ptchd2、foxd4l5、pkhd1l1、lgr4、dpf3、krtap5-1、hla-drb3、ighm、zrsr1、znf578和repin1。
利用本发明的方法,检测细胞在用化学试剂处理前和处理后的细胞核和胞浆的荧光强度,对比前后两次的检测结果,即可确定细胞中发生核转移(即从细胞质中转移到细胞核)的转录因子含量或分布情况,采用该方法可以对细胞核内的转录因子含量进行定量分析或分布情况检测,该方法具有高通量和更准确的量化分析特点。
附图说明
为了更清楚地说明本发明实施例的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,应当理解,以下附图仅示出了本发明的某些实施例,因此不应被看作是对范围的限定,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他相关的附图。
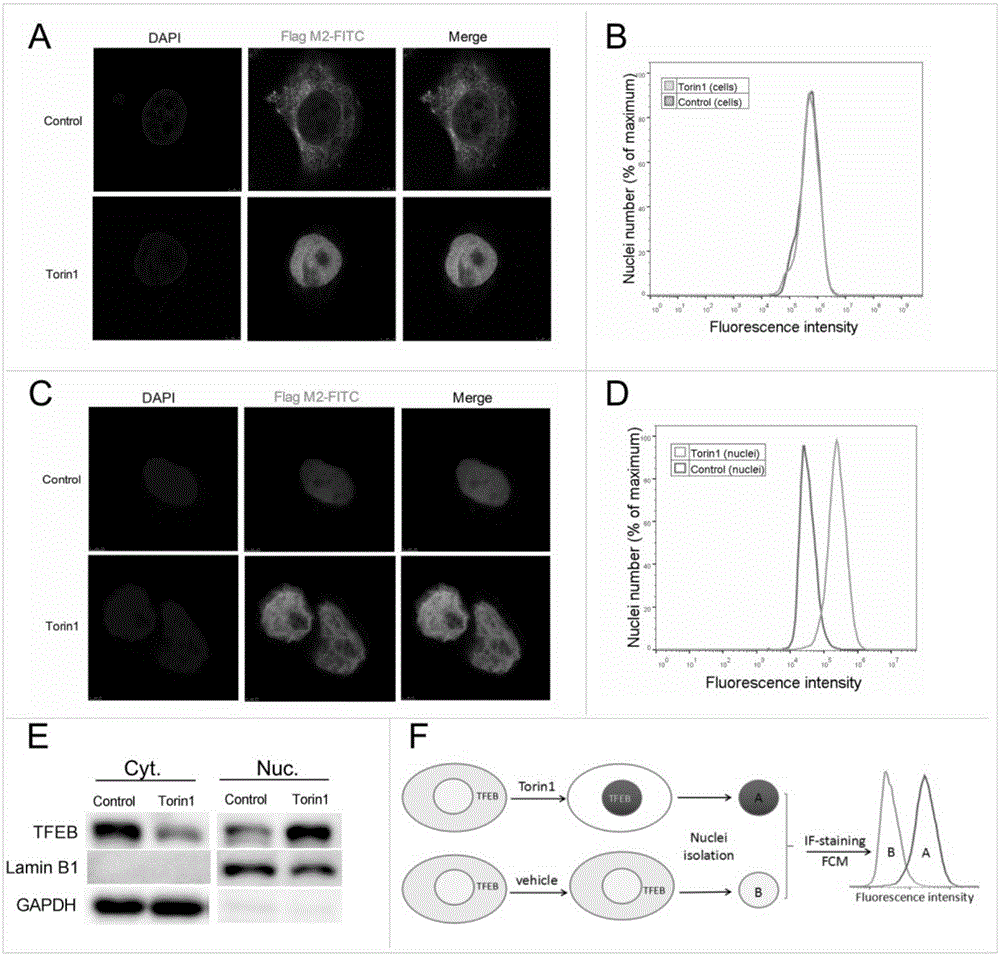
图1为本发明实验例1中通过fcm定量tfeb核转位;
图中:a至e用溶媒(0.1%dmso)和torin1(1μm)处理过表达3xflag-tfeb的hela细胞4小时。(a,c)共聚焦显微镜分析显示tfeb的亚细胞定位。用抗flagm2-fitc抗体和dapi孵育细胞和纯化的细胞核。(b,d)通过fcm进行tfeb亚细胞定位的定量。将细胞和纯化的细胞核与抗flagm2-fitc抗体孵育。(e)westernblot印迹分析tfeb胞浆(cyt.)和胞核(nuc.)的亚细胞定位。gapdh和laminb1作为蛋白内参。(f)通过fcm定量分析tfeb核转位的示意图。
图2为本发明实验例2中基于fcm的测定与传统方法在评估小分子化合物诱导的tfeb核转位中的比较;(0.1%dmso,对照组,ctrl),torin1(500nm),c1(1μm),海藻糖(100mm,tre),5g7(5μm)和6d6(5μm)处理过表达3xflaghela细胞12小时。
图中:(a,b)将细胞与抗flag抗体和dapi一起孵育。通过免疫荧光显微镜对tfeb亚细胞定位进行定量分析。分析来自3次独立实验的至少300个细胞。图a中的比例尺是10微米。(c,d)westernblot印迹分析tfeb胞浆(cyt.)和胞核(nuc.)的亚细胞定位。gapdh和laminb1作为蛋白内参。(e,f)通过fcm进行核tfeb的定量。纯化的细胞核与抗flagm2-fitc抗体孵育。数据显示为平均值±标准偏差。
图3为本发明实验例3中通过fcm定量内源性tfeb核转位;
图中:(a,c,e)hela细胞和(b,d,f)n2a细胞用0.1%dmso和torin1(1μm)处理4小时。(a,b)由incellanalyzer2000拍摄的图片显示了tfeb的不同亚细胞定位。比例尺为20微米。(c,d)通过fcm分析进行tfeb亚细胞定位的定量。纯化的细胞核使用内源性tfeb抗体染色。(e,f)westernblot印迹分析tfeb胞浆(cyt.)和胞核(nuc.)的亚细胞定位。gapdh和laminb1作为蛋白内参。*与对照相比,p<0.05。
图4为本发明实验例4中通过fcm定量分析小鼠肝细胞中内源性tfeb的核转位。通过腹膜内注射溶媒或torin1(20mg/kg,6小时)处理c57bl/6小鼠。图中:(a)westernblot印迹分析tfeb胞浆(cyt.)和胞核(nuc.)的亚细胞定位。gapdh和laminb1作为蛋白内参。(b)通过fcm进行核tfeb的定量。纯化的细胞核与内源性tfeb抗体孵育。*与对照相比,p<0.05。
具体实施方式
为使本发明实施例的目的、技术方案和优点更加清楚,下面将对本发明实施例中的技术方案进行清楚、完整地描述。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市售购买获得的常规产品。
以下结合实施例对本发明的特征和性能作进一步的详细描述。
实验例1
通过免疫染色测试了持续表达3xflag-tfeb(tfeb-hela)13的hela细胞中的tfeb易位。如先前报道,torin1显著诱导tfeb的核转位(图1a)。用含有0.1%dmso(溶媒)的正常培养基培养的对照细胞显示大多数tfeb分布在细胞质中,而torin1处理的细胞(1μm,4h)显示tfeb在细胞核中显著积累。然而,当用fcm分析时,两组之间的平均荧光强度(mfi)没有明显差异(图1b)。数据表明torin1主要改变tfeb的亚细胞定位而不影响其在细胞中的总量。通过使用核质分离试剂盒,来自两个组的细胞核被纯化并用抗flagm2-fitc抗体染色。与载体相比,torin1显著诱导tfeb在细胞核中的积累(图1c)。同时torin1处理组细胞核的mfi显著增高(图1d)。然后我们通过蛋白印迹法确定tfeb在细胞核(nuc.)和细胞质(cyt.)中的分布(图1e),蛋白印迹结果与fcm的发现一致。总之,上述结果显示fcm能够通过细胞核tfeb染色来有效定量tfeb的核转位(图1f)
实验例2
为了比较基于fcm的测定与用于定量分析tfeb活化的传统方法的定量,我们分别通过免疫荧光显微术(图2a,b),蛋白印迹(图2c,d)和fcm方法分析tfeb-hela细胞中的tfeb亚细胞定位(图2e,f)。除torin1外还使用了几种tfeb激活剂。例如,c1和海藻糖也是tfeb核转位的激活剂,并具有不同的机制16,18。通过高通量筛选发现化合物如5g7和6d6可以促进tfeb入核。在对照组,与torin1处理的细胞(超过90%)相比,tfeb入核的细胞占总的细胞百分比很小(约6%)。化合物c1、海藻糖、5g7和6d6则可以不同程度地诱导tfeb核转位(图2a)。随后通过蛋白印迹检测tfeb在细胞质和细胞核中的不同分布图。结果显示胞浆tfeb的减少程度与torin1,c1,海藻糖,5g7和6d6处理组中核tfeb的增加相对应(图2b)。然后我们用指定的处理方法纯化细胞核并进行fcm分析。正如预测的那样,我们通过计算不同组细胞核的mfi得到了与显微结果和蛋白质印迹分析相似的结果(图2c)。以上结果表明,这种新的基于fcm用于测定tfeb核转位的结果可以被常规方法验证。但是值得注意的是,我们的检测方法比免疫荧光显微镜更高效和更客观。此外,与蛋白印迹法相比,新的测定方法耗时更少,工作量更小,定量更加准确。
实验例3
基于fcm的测定法能够定量分析未转染3xflag-tfeb的细胞系中内源性tfeb的核转位。通过使用抗flagm2-fitc抗体,可以用fcm快速分析3xflag-tfebhela细胞中的tfeb亚细胞定位。然而,必须使用3xflag-tfeb质粒转染细胞的要求可能会限制本方法的应用范围。因此,我们在未转染的细胞系如野生型hela细胞(图3a)和neuro-2a(n2a)细胞(图3b)中测试了内源性tfeb抗体。torin1处理后,tfeb信号主要存在于核内。图中结果表明tfeb抗体适用于未转染细胞系中的免疫荧光染色。然后我们用hela细胞(图3c)和neuro-2a(n2a)细胞(图3d)进行基于fcm的分析。在用0.1%dmso或1μmtorin1培养4小时后,纯化hela和n2a细胞的细胞核,随后用tfeb抗体染色,随后进行荧光标记的二抗染色。与对照细胞相比,torin1处理的细胞中hela和n2a细胞的核mfi均显著增加。然后我们通过western印迹检测tfeb的亚细胞定位。结果显示torin1显著诱导hela细胞和n2a细胞中的tfeb核分布(图3e,f)。以上结果表明基于fcm量化tfeb易位的方法在非转染3xflag-tfeb的细胞系中效果良好。
实验例4
基于fcm的测定方法能够定量分析小鼠肝细胞中内源性tfeb的核转位。据报道,用torin1(20mg/kg)处理的c57bl/6小鼠显示mtorc1被显著抑制21,而mtorc1是tfeb入核的负性调节因素,因此我们推测在此情况下,小鼠组织细胞(比如肝脏组织)的tfeb存在明显的核转位。为了确定torin1诱导的肝组织中的tfeb核转位,我们首先通过蛋白印迹法检测溶媒或torin1(20mg/kg,6h)处理c57bl/6小鼠分离的肝组织细胞中tfeb亚细胞定位。正如预测的那样,与对照组相比,tfeb在torin1组中显著易位至核内(图4a)。接下来,我们分离提取了肝组织细胞的细胞核,用内源性tfeb抗体染色,随后用fcm进行分析。torin1组的mfi明显高于对照组(图4b)。总之,以上结果揭示了这种基于fcm的测定法能够检测在体组织细胞内tfeb核转位。
目前使用的定量tfeb核转位的方法包括通过显微拍照然后手动计数tfeb在细胞核中的细胞百分比15或通过蛋白印迹分析16tfeb在细胞核和细胞质部分中的表达,这些方法是相当主观的或具有较大的工作量。
流式细胞术(flowcytometry,fcm)已被应用于通过膜附着的lc3成像来测量自噬活性19-20。本研究中,我们建立了一个更客观,更高通量的检测tfeb核转位的方法。在溶媒或torin1(已知的tfeb活化剂15)处理后,分离3xflag-tfebhela细胞的细胞核并用抗flagm2-fitc抗体染色。核的平均荧光强度用于分析tfeb易位的效率。此外,通过使用内源性tfeb抗体染色,该测定方法可以扩展应用到在体外小鼠模型和离体的未转染3xflag-tfeb的细胞系中评估tfeb核转位。
在这项研究中,我们建立了一种新型的以流式细胞术(fcm)为基础来量化转录因子eb(tfeb)核转位的检测方法,而tfeb核转位是触发自噬-溶酶体途径生物发生的重要事件,对维持细胞内稳态和能量平衡至关重要。该测定法快速且客观,可以分析细胞内和动物组织细胞中的tfeb核转位。
在哺乳动物细胞中,自噬是降解细胞溶质成分和受损细胞器的主要途径,其功能障碍与多种病理生理过程有关2。
tfeb可能通过调节自噬和溶酶体生物合成而成为这些疾病的潜在治疗靶点,因为越来越多的证据强调了tfeb在多种神经退行性疾病4-6和溶酶体贮积病10-11等的细胞和动物模型中的治疗潜力。失活的tfeb(磷酸化形式)分布在细胞质中,而活化的tfeb(去磷酸化形式)转位进入细胞核以启动与自噬和溶酶体生物合成有关的基因的转录12-14。因此,定量tfeb的核转位是评估初始自噬体和溶酶体生物合成中tfeb活性的良好标记物。
监测体内自噬通量一直是一项非常具有挑战性的任务。自噬体标记物lc3-ii印迹或点的增加并不一定意味着自噬通量的增加,因为当自噬体成熟受损时它们增加得更加明显。尽管自噬通量探针如gfp-rfp-lc322或gfp-lc3-rfp-lc3△g23已用于监测体内自噬成熟,但横切过程和引入外源基因可能干扰观察。tfeb是控制自噬-溶酶体途径生物发生的主要转录因子,其核转位可做为自噬-溶酶体途径诱导发生的指示。我们的新测定方法可能提供一种简单且高效定量的方法来分析在体和离体细胞内自噬-溶酶体途径的活化。
fcm独特而强大的功能包括多种荧光标记和随后的事件分类。量化动物组织中特定类型细胞中的tfeb核转位是具有挑战性的,并且只能通过免疫组织荧光实现,这是一种不太客观和耗时的方法。通过选择在核中表达的细胞标记,通过fcm定量所需细胞类型中的tfeb核转位是非常方便和准确的。此外,在我们开发的测定方法中,细胞核完整,因此可以很容易地收集用于进一步分析。例如,可以将感兴趣的特定细胞核群挑选出来用于表观遗传学分析或核转录组分析。此外,当与强大的crispr/cas9介导的全基因组敲除方法结合时,该测定将非常适合于鉴定tfeb核转位的调节基因。
本发明提供的测定tfeb核转位的方法的定量能力已通过常规方法例如蛋白质印迹和显微镜学证实(图2)。基于检测原理,我们认为这种检测方法可以扩展到确定其他核转录因子或在不同情况下穿梭于细胞核和细胞质之间的蛋白质的核定位。
实验例1-4中所涉及的材料和方法参考如下:
材料和方法
细胞培养
过表达3xflag-tfeb(tfeb-hela)12-13的hela细胞来自telethon遗传与医学研究所的andreaballabio博士的慷慨赠与。细胞在含10%胎牛血清(fbs),100u/ml青霉素,100μg/ml链霉素和200μg/mlg418的dmem中培养,并在给药之前撤去g418。将hela和n2a细胞在含有有10%fbs,100u/ml青霉素和100μg/ml链霉素的dmem中培养。
动物饲养和给药
所有动物实验程序均符合澳门大学中华医药研究院伦理准则。
3个月大的c57bl/6雄性小鼠在12小时光照/黑暗循环的房间内饲养。将8只小鼠随机分为两组:对照组和torin1(每组4只小鼠)。torin1组小鼠禁食18h,然后腹腔注射torin1(20mg/kg,6h,sigma-aldrich,475991),6小时后,处死小鼠(co2窒息),收集肝组织,将组织粉碎并均化成单细胞悬液。将细胞悬浮液通过70μm过滤器过滤并用含有0.5%bsa和2mmedta的pbs冲洗。将细胞在450g下离心7分钟并重悬于裂解缓冲液(bdbiosciences,555899)中以裂解红细胞。在450g离心7分钟后,将肝细胞重悬于含有0.5%bsa和2mmedta的pbs中,并分成两份,一份用于蛋白质印迹,另一份用于流式细胞术。
显微镜成像
对于激光共聚焦显微镜成像,先采用本发明提供的基于流式细胞术定量分析细胞内tfeb核转位的方法进行细胞核处理,操作如下:
将tfeb-hela细胞分别接种在6孔板和底部上具有盖玻片的24孔板中,培养12h,并用0.1%dmso和500nmtorin1处理。在37℃12h后,去除6孔板中的培养基,pbs洗三次后,加入500μl核质分离试剂(sigma-aldrich,nuc101),将细胞刮下收集于2mlep管,冰上放置5min后,4℃、500g离心5min,弃去上清胞质,再次往沉淀中加入300μl核质分离试剂(sigma-aldrich,nuc101),冰上放置5min后,4℃、500g离心5min,弃去上清,沉淀即为分离得到的完整细胞核(以上步骤均在冰上或4℃条件下)。与细胞核一起,切片上的细胞用4%多聚甲醛固定,用0.2%tritonx-100透化并用5%正常山羊血清封闭。封闭后,完整细胞和细胞核与抗flagm2-fitc抗体(稀释比例(体积比)1:100,sigma-aldrich,f4049)在室温下孵育1小时。用dapi染色后,细胞核散布在微型载玻片上。将切片上的细胞和微型载玻片上的细胞核用抗褪色封片剂包埋封片,并使用共聚焦显微镜leicatcssp8smd(莱卡,德国)进行成像。
对于高通量显微镜成像,将tfeb-hela细胞,hela细胞和n2a细胞接种于96孔板并培养12小时。用0.1%dmso,500nmtorin1,1μmc1,100mm海藻糖(sigma-aldrich,1673715),10μm5g7和5μm6d6(prestwickchemical,greenpharm天然化合物库)处理tfeb-hela细胞,在补充有1%fbs的dmem中培养12小时。用0.1%dmso,1μmtorin1处理hela和n2a细胞4h。用多聚甲醛固定后,用tritonx-100透化并用正常山羊血清封闭,用抗flag抗体(1:500,sigma-aldrich,f7425)对tfeb-hela细胞进行染色,同时将hela和n2a细胞染色用抗tfeb抗体在4℃过夜。在dapi染色之前,在室温条件下将二抗alexa
蛋白质印迹
为了通过westernblot鉴定tfeb亚细胞定位,根据先前描述的方案分离胞核和胞质部分12。在通过10%sds-page凝胶分离后,将蛋白质转移到pvdf膜(bio-rad,1620177)和随后用5%bsa封闭。用兔抗tfeb(1:2000,bethyllaboratories,inc.,a303-673a),兔抗laminb1(cellsignalingtechnology,13435s),兔抗gapdh(1:1000,cellsignalingtechnology,5174s)和hrp标记的山羊抗兔二抗(1:2000,cellsignalingtechnology,7074s)。使用ecl试剂(bio-rad,1705061)检测蛋白质信号并用imagej软件分析。
流式细胞术
对于流式细胞术测定,将tfeb-hela细胞,野生型hela和n2a细胞接种到6孔板上。培养12小时后,细胞用0.1%dmso和1μmtorin1处理4小时,使用nucleiezprep试剂盒,将wthela细胞,n2a细胞,一半的tfeb-hela细胞和来自小鼠的一半肝细胞应用于分离细胞核。与tfeb-hela的细胞核一起,在用多聚甲醛固定后,用tritonx-100透化并用正常山羊血清封闭,将另一半的tfeb-hela细胞与抗flagm2-fitc抗体孵育。同时,将hela细胞,n2a细胞和肝细胞的细胞核与内源性抗tfeb抗体(bethyllaboratories,inc.,a303-673a)一起在4℃温育过夜,然后加入二抗alexa
统计分析
数值表示为平均值±sd。数据使用单因素方差分析(anova)和dunnett's检验进行分析。概率值p小于0.05被认为具有显著性差异。
相关参考文献如下:
1.mizushima,n.;levine,b.;cuervo,a.m.;klionsky,d.j.,autophagyfightsdiseasethroughcellularself-digestion.nature2008,451(7182),1069-1075.
2.rubinsztein,d.c.;codogno,p.;levine,b.,autophagymodulationasapotentialtherapeutictargetfordiversediseases.naturereviewsdrugdiscovery2012,11,709.
3.galluzzi,l.;bravo-sanpedro,j.m.;levine,b.;green,d.r.;kroemer,g.,pharmacologicalmodulationofautophagy:therapeuticpotentialandpersistingobstacles.naturereviews.drugdiscovery2017,16(7),487-511.
4.decressac,m.;mattsson,b.;weikop,p.;lundblad,m.;jakobsson,j.;
5.xiao,q.;yan,p.;ma,x.;liu,h.;perez,r.;zhu,a.;gonzales,e.;burchett,j.m.;schuler,d.r.;cirrito,j.r.;diwan,a.;lee,j.-m.,enhancingastrocyticlysosomebiogenesisfacilitatesaβclearanceandattenuatesamyloidplaquepathogenesis.thejournalofneuroscience2014,34(29),9607-9620.
6.tsunemi,t.;ashe,t.d.;morrison,b.e.;soriano,k.r.;au,j.;vázquezroque,r.a.;lazarowski,e.r.;damian,v.a.;masliah,e.;laspada,a.r.,pgc-1αrescueshuntington’sdiseaseproteotoxicitybypreventingoxidativestressandpromotingtfebfunction.sciencetranslationalmedicine2012,4(142),142ra97-142ra97.
7.sergin,i.;evans,t.d.;zhang,x.;bhattacharya,s.;stokes,c.j.;song,e.;ali,s.;dehestani,b.;holloway,k.b.;micevych,p.s.;javaheri,a.;crowley,j.r.;ballabio,a.;schilling,j.d.;epelman,s.;weihl,c.c.;diwan,a.;fan,d.;zayed,m.a.;razani,b.,exploitingmacrophageautophagy-lysosomalbiogenesisasatherapyforatherosclerosis.naturecommunications2017,8,15750.
8.pan,b.;zhang,h.;cui,t.;wang,x.,tfebactivationprotectsagainstcardiacproteotoxicityviaincreasingautophagicflux.journalofmolecularandcellularcardiology2017,113,51-62.
9.spampanato,c.;feeney,e.;li,l.;cardone,m.;lim,j.-a.;annunziata,f.;zare,h.;polishchuk,r.;puertollano,r.;parenti,g.;ballabio,a.;raben,n.,transcriptionfactoreb(tfeb)isanewtherapeutictargetforpompedisease.embomolecularmedicine2013,5(5),691-706.
10.medina,diegol.;fraldi,a.;bouche,v.;annunziata,f.;mansueto,g.;spampanato,c.;puri,c.;pignata,a.;martina,josea.;sardiello,m.;palmieri,m.;polishchuk,r.;puertollano,r.;ballabio,a.,transcriptionalactivationoflysosomalexocytosispromotescellularclearance.developmentalcell2011,21(3),421-430.
11.moskot,m.;montefusco,s.;jakóbkiewicz-banecka,j.;mozolewski,p.;
12.settembre,c.;dimalta,c.;polito,v.a.;arencibia,m.g.;vetrini,f.;erdin,s.;erdin,s.u.;huynh,t.;medina,d.;colella,p.;sardiello,m.;rubinsztein,d.c.;ballabio,a.,tfeblinksautophagytolysosomalbiogenesis.science(newyork,n.y.)2011,332(6036),1429-1433.
13.sardiello,m.;palmieri,m.;dironza,a.;medina,d.l.;valenza,m.;gennarino,v.a.;dimalta,c.;donaudy,f.;embrione,v.;polishchuk,r.s.;banfi,s.;parenti,g.;cattaneo,e.;ballabio,a.,agenenetworkregulatinglysosomalbiogenesisandfunction.science2009,325(5939),473.
14.napolitano,g.;ballabio,a.,tfebataglance.journalofcellscience2016,129(13),2475-2481.
15.vega-rubin-de-celis,s.;
17.sha,y.;rao,l.;settembre,c.;ballabio,a.;eissa,n.t.,stub1regulatestfeb‐inducedautophagy–lysosomepathway.theembojournal2017,36(17),2544.
18.song,j.-x.;sun,y.-r.;peluso,i.;zeng,y.;yu,x.;lu,j.-h.;xu,z.;wang,m.-z.;liu,l.-f.;huang,y.-y.;chen,l.-l.;durairajan,s.s.k.;zhang,h.-j.;zhou,b.;zhang,h.-q.;lu,a.;ballabio,a.;medina,d.l.;guo,z.;li,m.,anovelcurcuminanalogbindstoandactivatestfebinvitroandinvivoindependentofmtorinhibition.autophagy2016,12(8),1372-1389.
19.shvets,e.;fass,e.;elazar,z.,utlizingflowcytometrytomonitorautophagyinlivingmammaliancells.2008;vol.4,p621-8.
20.eng,k.e.;panas,m.d.;hedestam,g.b.k.;mcinerney,g.m.,anovelquantitativeflowcytometry-basedassayforautophagy.autophagy2010,6(5),634-641.
21.liu,q.;chang,j.w.;wang,j.;kang,s.a.;thoreen,c.c.;markhard,a.;hur,w.;zhang,j.;sim,t.;sabatini,d.m.;gray,n.s.,discoveryof1-(4-(4-propionylpiperazin-1-yl)-3-(trifluoromethyl)phenyl)-9-(quinolin-3-yl)benzo[h][1,6]naphthyridin-2(1h)-oneasahighlypotent,selectivemammaliantargetofrapamycin(mtor)inhibitorforthetreatmentofcancer.journalofmedicinalchemistry2010,53(19),7146-7155.
22.castillo,k.;valenzuela,v.;matus,s.;nassif,m.;
23.kaizuka,t.;morishita,h.;hama,y.;tsukamoto,s.;matsui,t.;toyota,y.;kodama,a.;ishihara,t.;mizushima,t.;mizushima,n.,anautophagicfluxprobethatreleasesaninternalcontrol.molecularcell2016,64(4),835-849.
以上所述仅为本发明的优选实施例而已,并不用于限制本发明,对于本领域的技术人员来说,本发明可以有各种更改和变化。凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
- 还没有人留言评论。精彩留言会获得点赞!
- 一种基于近红外定性分析的种子品种真实性鉴别方法
- 一种基于近红外提高鉴别结果的定性分析方法
- 一种应用于小分子定性定量分析检测和高通量筛选微生物培养液样品试剂盒及其制备方法
- 一种应用于小肽分子定性定量分析检测和高通量筛选增强试剂盒及其制备方法
- 一种应用于污染物小分子定性定量分析检测和高通量筛选增强试剂盒及其制备方法
- 一种应用于小分子定性定量分析检测和高通量筛选细胞培养液样品试剂盒及其制备方法
- 一种应用于小分子定性定量分析检测和高通量筛选血液样品试剂盒及其制备方法
- 一种应用于碱性小分子定性定量分析检测和高通量筛选增强电离缓冲液试剂盒及其制备方法
- 一种应用于胆碱及脂类分子定性定量分析检测和高通量筛选增强试剂盒及其制备方法
- 一种应用于三聚氰胺分子定性定量分析检测和高通量筛选增强试剂盒及其制备方法